光照强度和盐度对浒苔幼苗生长和光合生理的影响*
2022-02-18王津果陈泽宇倪嘉璇何昊林
王津果,陈泽宇,倪嘉璇,何昊林,周 伟
(江苏海洋大学 a.江苏省海洋生物资源与环境重点实验室; b.江苏省海洋生物产业技术协同创新中心; c.江苏省海洋生物技术重点实验室; d.自然资源部滨海盐沼湿地生态与资源重点实验室,江苏 连云港 222005)
0 引言
20世纪70年代,在工业化国家的近岸海域中发现某些大型绿藻开始暴发性增殖或高度聚集,这种现象被称为绿潮[1]。自2008年始,我国黄海大规模绿潮持续发生,至今已有15年之久,成为困扰沿海民生的重大生态灾难,严重威胁沿海旅游业、渔业和养殖业的发展,给沿岸省、市带来严重的经济损失。绿潮漂浮规模大、持续时间长、暴发频率高、危害程度深,已引起世界的广泛关注[2]。
现有研究发现,黄海绿潮的优势种为浒苔(Ulvaprolifera)。作为一种机会主义海藻,其形成是自身生物学特点、海水富营养化和环境条件适宜等综合作用的结果[3]。自身生物学特点如适应能力强、繁殖速度快等是绿潮形成的首要关键因素。浒苔通常具有营养繁殖、无性生殖和有性生殖3种繁殖方式,生活史中的任一形态均可以单独发育为成熟藻体[4-5]。其中只进行营养繁殖的二倍体细胞团、脱落下来的组织块、单细胞等碎片即微观繁殖体,具有抗逆性强、增殖快速等特点,在越冬和度夏等恶劣环境下作为绿潮的“种子库”,待到环境条件适宜时重新生长发育,可见其在绿潮早期暴发过程中发挥着不容小觑的作用[6]。目前关于浒苔微观繁殖体的研究主要集中在其时空分布特征等方面[7-9],而关于非生物因素如光照强度、盐度等对浒苔微观繁殖体影响的研究少有报道。
光照是藻类进行光合作用的主要能量来源,藻类生长的快慢和光合作用的强弱在一定程度上取决于光照强度的高低[10]。光照强度适度增加有利于提高藻体的生长速率、光合速率和量子产量、光化学效率、电子传递速率等叶绿素荧光参数,在浒苔成体[10-11]、孔石莼(U.pertusa)[12]、针叶蕨藻(Caulerpasertularioides)[13]等大型海藻中均有报道。另有研究表明,光照强度的高低会影响藻类光合色素的合成。相较于高光强,低光强处理可促进浒苔成体[14]、细基江蓠(Gracilariatenuistipitata)[15]、龙须菜(Gracilarialemaneiformis)[16]、缘管浒苔(U.linza)[17]的光合色素合成。海水盐度易受到潮汐、蒸发和降雨的影响,近海岸生态系统中的大型海藻会受到盐度胁迫强烈且持续的影响[18]。前人研究表明,盐度的适当增加可促进浒苔成体[19]、孔石莼[20]、裂片石莼(U.fasciata)幼苗[21]的生长速率、光合速率;大型海藻如缘管浒苔[22]、绳江蓠(G.chorda)、真江蓠(G.vermiculophylla)[23]、针叶蕨藻(C.sertularioides)[13]的生长速率、光合速率和量子产量、电子传递速率等叶绿素荧光参数随着盐度的降低而下降。此外,盐度变化会影响藻类光合色素的合成。He等[14]的研究表明高盐(40)处理可促进浒苔成体的叶绿素a、叶绿素b和类胡萝卜素的合成;盐度的适当降低(20)可促进孔石莼的生长速率、氮吸收速率、磷吸收速率和叶绿素a含量的提升,盐度过低或过高均会对其生长及营养盐吸收速率产生一定的抑制作用[23]。
目前已有大量关于浒苔成体响应光照强度和盐度等非生物因素的研究[10-11,14,19,24],但关于二者对绿潮早期暴发种子库的浒苔微观繁殖体幼苗耦合影响的研究少见报道。另外,调查发现黄海沿岸海水养殖池塘和育苗厂进/排水渠中常有浒苔漂浮其中,进/排水渠易受到生活污物排放和降雨的影响,导致光照强度、盐度等环境因子发生较大变化。鉴于此,本研究以浒苔微观繁殖体幼苗为材料,在室外自然光照条件下,探究光照强度、盐度变化对浒苔幼苗生长和光合生理的影响,以期进一步揭示浒苔绿潮早期暴发的原因,为其预警防控提供理论依据。
1 材料与方法
1.1 材料及培养条件
浒苔水样于2022年4月2日采自江苏省盐城市射阳县兴皇长江蟹苗场进/排水渠(33°86′N,120°47′E)。利用孔径为200 μm的筛绢过滤掉其中的大型浮游植物和藻丝体,低温避光保存带回实验室。向过滤后的水样中加入Provasoli培养液[25]和GeO2以抑制硅藻生长,置于温度20 ℃、光照强度100 μmol/(m2·s)、光周期12 L:12 D的智能光照培养箱(GXZ-500C,中国)中通气培养,14 d后微观繁殖体萌发生长为0.5~1 cm的幼苗,用于后续实验。
1.2 实验设计
挑选0.8 cm左右的幼苗(0.20±0.01)g置于含过滤灭菌海水的500 mL圆底通气瓶中,在室外500 L不锈钢水槽中进行藻体培养。设置2个光照强度水平(低光强:LL;高光强:HL)和3个盐度梯度(LS:12;MS:22;HS:32),即共6个光照强度和盐度组合处理组,每个组合设置3个平行样。通过在水槽上方不加遮阳网、加2层遮阳网分别获得高光强、低光强培养条件。实验用培养基均为过滤灭菌后的水样采集地海水,测定其盐度为32,利用超纯水稀释分别获得盐度为22和12的培养基。水槽温度通过恒温循环器(DHX-2005,中国)控制在20 ℃,每2 d更换一次培养基。
1.3 室外太阳辐射测定
实验期间的太阳辐射通过太阳辐射接收器(Logger Net CR3000,美国)监测和记录,单位μmol/(m2·s),日剂量单位MJ/m2。
1.4 相对生长速率测定
每2 d称量一次藻体质量。使用镊子取出藻体,吸水纸轻轻吸干其表面水分后,称量。为减少操作误差,由同一个人进行称量,并保持吸水时间和吸水纸层数一致。另外,应尽量减少在空气中的干露时间,以防损伤藻体生理活性[26]。相对生长速率(RGR,%/d)计算公式如下:
RGR=100×ln(Wt/W0)/t。
(1)
式中:Wt表示第t天藻体的质量,g;W0表示藻体初始质量,g;t表示培养天数,d。
1.5 净光合速率测定
采用Clark液相氧电极(YSI 5300A,美国)测定藻体的净光合速率。将待测藻体剪成1 cm左右的小段,置于培养条件下适应1 h以上,以减少机械损伤。称取约0.04 g藻段置于含8 mL培养基的反应室中,由恒温循环器(DHX-2005,中国)控制温度在20 ℃。采用卤素灯提供外源光照强度,通过调整卤素灯和反应室间的距离获得8个光化光强(0,10,20,50,100,200,500,1 000 μmol/(m2·s))梯度下的净光合速率(单位μmol/(g·h))。光响应曲线(P—I曲线)根据Henley[27]方法进行拟合并计算光合作用的饱和光强(Ik)和光补偿点(Ic),计算公式如下:
P=Pm×tanh(α×PAR/Pm)+Rd,
(2)
Ik=Pm/α,
(3)
Ic=-Rd/α。
(4)
式中:PAR表示光化光强,μmol/(m2·s);P表示光化光强为PAR时相应的净光合速率,μmol/(g·h);Pm是最大净光合速率,μmol/(g·h);α表示光能利用效率;Rd表示暗呼吸速率,μmol/(g·h)。
1.6 叶绿素荧光参数测定
采用手持式PAM叶绿素荧光仪(AquaPen AP-P 100 Chech,捷克)测定藻体的叶绿素荧光参数。分别在8:30,10:00,11:30,13:00,14:30,16:00,17:30,即每1.5 h测定一次有效光合量子产量(Y(II))。在8个光化光强(0,10,20,50,100,200,500,1 000 μmol/(m2·s))梯度下测定相对电子传递速率(rETR),计算公式如下[28]:
rETR=Y(II)×0.5×PAR。
(5)
式中:Y(II)为光系统II的有效光合量子产量;0.5为光系统II吸收的光量子占总量的比例;PAR为光化光强,μmol/(m2·s)。
快速光响应曲线(RLC)根据Eilers等[29]方法进行拟合,计算公式如下:
rETR=PAR/(a×PAR2+b×PAR+c)。
(6)
式中:rETR为相对电子传递速率;PAR为光化光强;a,b和c为拟合参数。
根据拟合参数计算最大相对电子传递速率(rETRmax)、光能利用效率(α)及饱和光强(Ek),计算公式如下:
rETRmax=1/[b+2(ac)1/2],
(7)
α=1/c,
(8)
Ek=rETRmax/α。
(9)
1.7 光合色素质量分数测定
称取约0.05 g藻体置于离心管中,加入5 mL无水甲醇,于4 ℃冰箱中避光放置24 h,离心,取上清液。分光光度计分别测定提取液在666,653,470 nm波长处的吸光度值。根据以下公式计算叶绿素a(Chl a)、叶绿素b(Chl b)、类胡萝卜素(Cartenoids,Car)的质量分数[30]:
w(Chl a)=15.65A666-7.53A653,
(10)
w(Chl b)=27.05A653-11.21A666,
(11)
w(Car)=(1 000A470+1 403.57A666-
3 473.87A653)/221。
(12)
式中:w(Chl a)为叶绿素a质量分数,mg/g;w(Chl b)为叶绿素b质量分数,mg/g;w(Car)为类胡萝卜素质量分数,mg/g;A666,A653,A470分别为666,653,470 nm波长处的吸光度值。
1.8 数据分析
实验数据均以“平均值±标准差(X±SD)”表示,利用SPSS 21.0和Origin 9.1软件进行数据处理并进行作图和统计分析。采用K-S和Levene检验来检验数据的正态性和方差齐性。利用单因素方差(One-way ANOVA)分析及Turkey’s多重比较检验处理组间浒苔幼苗的生长和光合生理指标的统计学差异。利用双因素方差(Two-way ANOVA)分析光照强度和盐度对浒苔幼苗生长和光合生理指标的交互作用。以p<0.05作为差异显著性水平。
2 结果与分析
2.1 室外太阳辐射
实验期间室外太阳辐射(实际光强)日平均变化如图1a所示,自7:00开始逐渐升高,于11:00左右达到最高值1 349.56 μmol/(m2·s),随着时间推移而逐渐下降,至19:00左右开始趋于平缓。实验期间太阳辐射日剂量变化如图1b所示,在3.21~9.64 MJ/m2范围内波动,15 d的平均日剂量是7.70 MJ/m2。

a 日平均变化
2.2 光照强度和盐度对浒苔幼苗相对生长速率的影响
双因素方差分析结果显示,光照强度、盐度和二者交互作用对浒苔幼苗的相对生长速率(RGR)均有显著影响(p<0.05)。如图2所示,在LL条件下,RGR随着盐度增加呈现出先升高后降低的趋势,在MS处理组达到最高值(27.39±0.10)%/d,相较LS和HS处理组,增幅分别为68.71%(LLLS)和14.36%(LLHS);在HL条件下,RGR随着盐度增加呈现出显著升高的趋势(p<0.05),在HS处理组达到最高值(25.86±0.06)%/d,比LS和MS处理组分别提高19.13%(HLLS)和14.15%(HLMS)。此外,在LS和HS条件下,HL处理组的RGR显著高于LL处理组(p<0.05),增幅分别为33.62%和7.97%;在MS条件下,HL处理组的RGR显著低于LL处理组(p<0.05),降幅17.31%。

注:不同小写字母表示在LL条件下不同处理间差异显著(p<0.05),不同大写字母表示在HL条件下不同处理间差异显著(p<0.05);“*”表示同一盐度下不同光照强度水平间差异显著(p<0.05)。后图同此。
2.3 光照强度和盐度对浒苔幼苗净光合速率的影响
光响应曲线(P—I曲线)显示,不同处理下浒苔幼苗的净光合速率随着光照强度的增加逐渐增加而后趋于平稳,且在LLMS处理组出现最高值(见图3)。根据P—I曲线计算得出的浒苔幼苗光合作用的最大净光合速率(Pm)、光能利用效率(α)、暗呼吸速率(Rd)、饱和光强(Ik)、光补偿点(Ic)见表1。
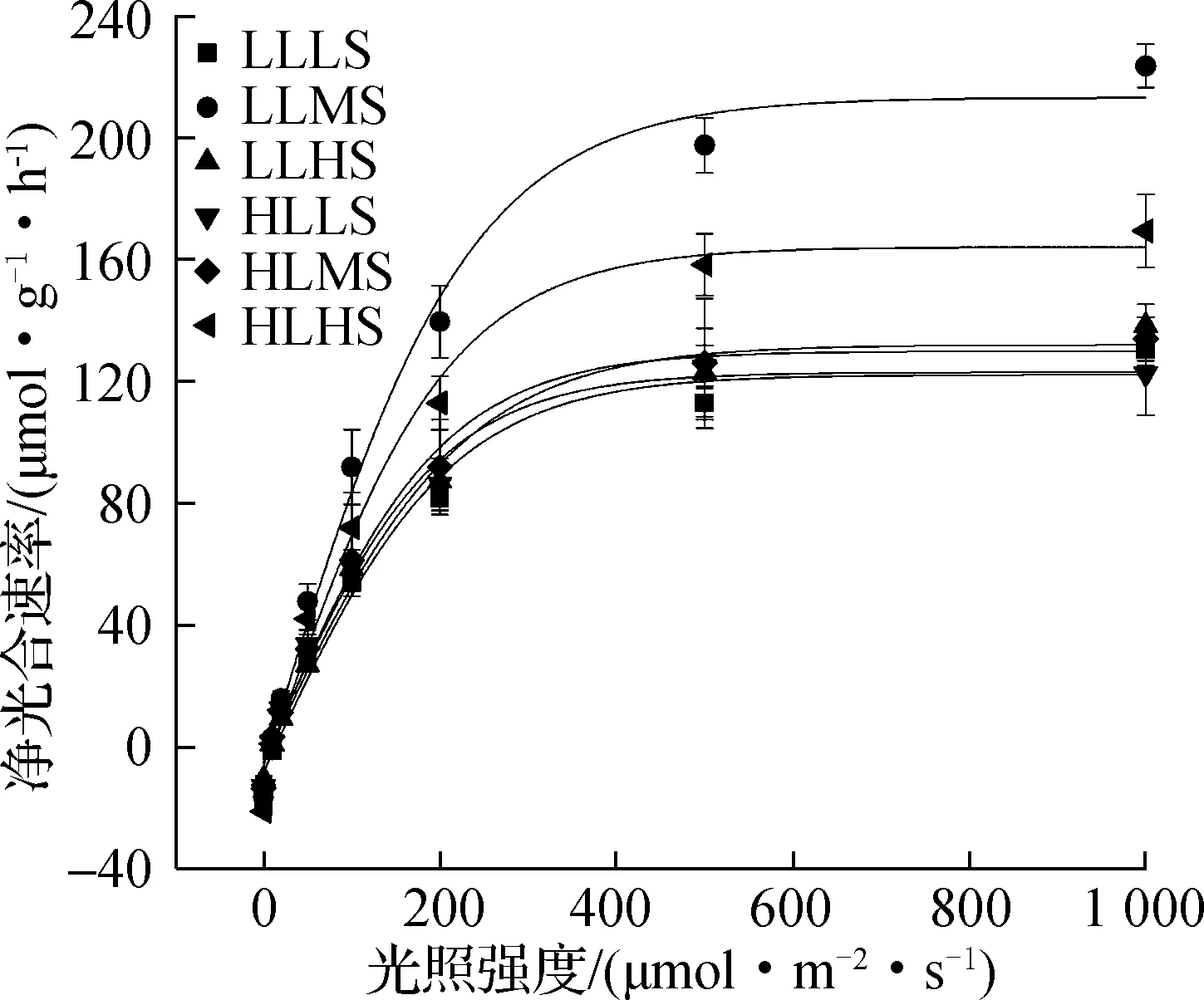
图3 不同处理下浒苔幼苗的光响应曲线(P—I曲线)
双因素方差分析结果显示,光照强度、盐度和二者交互作用对浒苔幼苗的Pm均有显著影响(p<0.05)。从表1中可以看出,在LL条件下,Pm随着盐度增加呈现出先升高后降低趋势,与RGR变化趋势相同,在MS处理组达到最高值,相较LS和HS处理组,增幅分别为68.44%(LLLS)和60.33%(LLHS);在HL条件下,Pm随着盐度增加呈现出逐渐升高趋势,与RGR变化趋势相同,在HS处理组达到最高值,比LS和MS处理组分别提高35.41%(HLLS)和27.97%(HLMS)。此外,在LS条件下,HL处理组的Pm低于LL处理组,但无显著差异(p>0.05);在MS条件下,HL处理组的Pm显著低于LL处理组(p<0.05),降幅38.37%;在HS条件下,HL处理组的Pm显著高于LL处理组(p<0.05),增幅26.44%。

表1 不同处理下浒苔幼苗的净光合速率(P)与光照强度关系的最佳拟合参数
双因素方差分析结果显示,光照强度对浒苔幼苗α无显著影响(p>0.05),盐度及二者交互作用对其有显著影响(p<0.05)。从表1中可以看出,在LL条件下,α随着盐度增加呈现出先升高后降低趋势,与Pm变化趋势相同,在MS处理组达到最高值,相较LS和HS处理组,增幅均为54.84%(LLLS和LLHS);在HL条件下,α随着盐度增加呈现出逐渐升高趋势,与Pm变化趋势相同,在HS处理组达到最高值,比LS和MS处理组分别提高30.30%(HLLS)和26.47%(HLMS)。此外,在LS和MS条件下,LL和HL处理组α无显著差异(p>0.05);在HS条件下,HL处理组的α显著高于LL处理组(p<0.05),增幅38.71%。
双因素方差分析结果显示,光照强度、盐度对浒苔幼苗的Rd无显著影响(p>0.05),但二者交互作用对其有显著影响(p<0.05)。如表1所示,在LL条件下,Rd随着盐度增加呈现出逐渐下降的趋势,在HS处理组达到最低值,相较LS和MS处理组,降幅分别为37.52%(LLLS)和20.22%(LLMS);在HL条件下,Rd随着盐度增加呈现出逐渐升高的趋势,与Pm变化趋势相同,在HS处理组达到最高值,比LS和MS处理组分别提高43.44%(HLLS)和31.09%(HLMS)。此外,在LS条件下,HL处理组的Rd显著低于LL处理组(p<0.05),降幅40.56%;在MS条件下,LL和HL处理组间的Rd无显著差异(p>0.05);在HS条件下,HL处理组的Rd显著高于LL处理组(p<0.05),增幅36.45%。
双因素方差分析结果显示,光照强度、盐度及二者交互作用对Ik无显著影响(p>0.05)。表1中,在同一光照强度条件下,不同盐度处理组藻体的Ik无显著差异(p>0.05)。此外,在同一盐度条件下,不同光照强度处理组藻体的Ik无显著差异(p>0.05)。
双因素方差分析结果显示,光照强度对浒苔幼苗的Ic无显著影响(p>0.05),盐度及二者交互作用对其有显著影响(p<0.05)。表1中,LL条件下,Ic随着盐度增加呈现出先降低后升高的趋势,在LS处理组达到最高值,相较MS和HS处理组,增幅分别为95.83%(LLMS)和58.55%(LLHS);在HL条件下,Ic随着盐度增加呈现出逐渐升高的趋势,与Pm变化趋势相同,在HS处理组达到最高值,比LS和MS处理组分别提高47.21%(HLLS)和39.25%(HLMS)。此外,在LS条件下,HL处理组的Ic显著低于LL处理组(p<0.05),降幅44.15%;在MS和HS条件下,LL和HL处理组间的Ic无显著差异(p>0.05)。
2.4 光照强度和盐度对浒苔幼苗荧光参数的影响
不同处理下浒苔幼苗的有效光合量子产量(Y(II))日变化如图4所示,在0.24~0.40范围内波动。在LLMS,LLHS,HLMS和HLHS培养条件下,Y(II)随着时间推移呈现出先升高后降低的趋势,在11:30达到峰值,分别为0.40±0.01,0.37±0.01,0.34±0.01,0.39±0.01,随后急剧下降,于13:00之后趋于稳定;在LLLS和HLLS条件下,Y(II)随着时间推移也呈现出先升高后降低的趋势,在10:00达到峰值,均为0.33±0.01,随后缓慢下降,于13:00之后趋于稳定。

图4 不同处理下浒苔幼苗的有效光合量子产量日变化
不同处理下浒苔幼苗的相对电子传递速率(rETR)随着光照强度的增加呈现出先升高后降低的趋势,在光照强度500 μmol/(m2·s)时达到峰值(见图5)。根据图4计算得出的不同处理下浒苔幼苗的最大相对电子传递速率(rETRmax)、光能利用效率(α)、饱和光强(Ik)如表2所示。

图5 不同处理下浒苔幼苗的相对电子传递速率
双因素方差分析结果显示,光照强度对浒苔幼苗rETRmax无显著影响(p>0.05),盐度及二者交互作用对其存在显著影响(p<0.05)。如表2所示,在LL条件下,rETRmax随着盐度增加呈现出先升高后降低的趋势,且在不同盐度处理组间存在显著差异(p<0.05);在HL条件下,rETRmax随着盐度增加呈现出逐渐升高的趋势,且HS处理组的rETRmax显著高于LS处理组(p<0.05),增幅18.32%。此外,在LS和HS条件下,LL和HL处理组间的rETRmax无显著差异(p>0.05);在MS条件下,HL处理组的rETRmax显著低于LL处理组(p>0.05),降幅12.80%。

表2 不同处理下浒苔幼苗的相对电子传递速率(rETR)与光照强度关系的最佳拟合参数
双因素方差分析结果显示,光照强度、盐度及二者交互作用对浒苔幼苗α和Ik均无显著影响(p>0.05)。
2.5 光照强度和盐度对浒苔幼苗光合色素含量的影响
双因素方差分析结果显示,光照强度、盐度及二者交互作用对浒苔幼苗叶绿素a(Chl a)、叶绿素b(Chl b)、类胡萝卜素(Car)质量分数存在显著影响(p<0.05)。
如图6a所示,在同一光照强度条件下,浒苔幼苗的Chl a质量分数随着盐度增加呈现出先降低后升高的趋势,在LS处理组达到高值,分别为(1.24±0.13)mg/g(LLLS)和(0.96±0.05)mg/g(HLLS),比HS处理组分别增加51.71%和154.76%,MS处理组的Chl a质量分数显著低于HS处理组(p<0.05),降幅分别为46.15%(LL)和20.00%(HL)。此外,在同一盐度条件下,HL处理组的Chl a质量分数显著低于LL处理组(p<0.05),降幅分别为22.58%(LS),32.61%(MS)和53.66%(HS)。

a 叶绿素a
如图6b所示,在同一光照强度条件下,浒苔幼苗的Chl b质量分数随着盐度增加呈现出先降低后升高的趋势,与Chl a质量分数的变化趋势相同,在LS处理组达到高值,分别为(1.40±0.04)mg/g(LLLS)和(0.73±0.09)mg/g(HLLS),比HS处理组分别增加12.90%和135.48%,MS处理组的Chl b质量分数显著低于HS处理组(p<0.05),降幅分别为66.94%(LL)和12.90%(HL)。此外,在同一盐度条件下,HL处理组的Chl b质量分数显著低于LL处理组(p<0.05),降幅分别为47.86%(LS),34.15%(MS)和75.00%(HS)。
如图6c所示,在LL条件下,浒苔幼苗的Car质量分数随着盐度增加呈现出先升高后降低的趋势,在MS处理组达到最高值(0.23±0.01)mg/g,分别是LS和HS处理组的2.30倍和5.75倍;在HL条件下,Car质量分数随着盐度增加呈现出先降低后升高的趋势,在LS处理组达到最高值(0.34±0.03)mg/g,分别是MS和HS处理组的2.43倍和1.70倍。此外,在LS和HS条件下,HL处理组Car质量分数显著高于LL处理组(p<0.05),增幅分别为2.40倍和4.00倍;在MS条件下,HL处理组Car质量分数显著低于LL处理组(p<0.05),降幅39.13%。
3 讨论
本研究结果显示,光照强度、盐度及二者交互作用明显影响浒苔幼苗的生长和光合生理指标,在不同处理下始终保持16.24%/d~27.39%/d的相对生长速率,表明浒苔幼苗可以耐受光照强度和盐度的范围比较广。另外,浒苔幼苗对环境变化较为敏感,低盐(LS)对其相对生长速率、净光合速率、有效光合量子产量、相对电子传递速率存在抑制作用,低光强、盐度适当降低(MS)可促进其相对生长速率、净光合速率、光能利用效率、有效光合量子产量、相对电子传递速率。在大型海藻如绿藻缘管浒苔[31]、红藻伊谷藻(Ahnfeltiaplicata)、褐藻齿缘墨角藻(Fucusserratus)和鞭状索藻(Chordariaflagelliformis)[32]等的研究中,藻体存在光抑制,即光照强度高于饱和光强后净光合速率出现下降的现象。在不同处理下浒苔幼苗的净光合速率均随着光照强度的增加而逐渐增加并趋于平稳,藻体净光合速率光照强度高于饱和光强后没有明显下降,而是出现增加的趋势,表明本研究中不同处理下的浒苔幼苗没有出现明显的光抑制现象,与Xiao等[19]关于不同温度和盐度处理下浒苔成体光响应曲线的研究结果相一致。
光照是藻类光合作用过程中同化力形成所需的能量来源,光照强度过低时,藻类光合碳同化过程因光合作用关键酶未充分活化或者同化力不足而受到限制;光照强度过高时,可能引发藻类光抑制,甚至破坏光合结构,进而影响藻类光合作用[19]。本研究结果显示,光照强度对浒苔幼苗相对生长速率存在显著影响(p<0.05)。在LS和HS条件下,低光强处理组浒苔幼苗相对生长速率显著低于高光强处理组(p<0.05),表明在高光强条件下浒苔幼苗生长更具优势,该现象在绿藻浒苔成体[10-11]、针叶蕨藻[13]、缘管浒苔[17]和红藻绳状龙须菜[33]、真江蓠[34]、褐藻裙带菜(Undariapinnatifida)[35]等大型海藻研究中均有报道。叶绿素一方面可作为天线色素,参与光能吸收和传递,另一方面少数可作为反应中心色素,被激发后发生电荷分离。本研究结果显示,同一盐度条件下,低光强处理组浒苔幼苗的叶绿素a、叶绿素b含量显著高于高光强处理组(p<0.05),这种光合色素含量与光照强度成负相关的现象在其他大型海藻如浒苔成体[14]、龙须菜[16]、缘管浒苔[17]等的研究中均有报道。原因可能是在低光强条件下,藻体通过补偿性地合成光合色素来弥补光照不足引起的相对生长速率和光能利用效率下降的问题,是藻体积极响应外界环境变化的一种机制,具有重要的生理生态作用[35-36]。
盐度变化会改变藻体细胞的渗透压,进而影响其生长和光合性能[37]。本研究结果显示,在LL条件下,盐度的适当降低(MS)促进了浒苔幼苗的相对生长速率、净光合速率、光能利用效率、有效光合量子产量、相对电子传递速率,表明盐度的适当降低(MS)有利于浒苔幼苗的生长和光合作用,与绿藻针叶蕨藻[13]、浒苔成体[19]、裂片石莼幼苗[21]、孔石莼[24]和红藻绳江蓠、真江蓠[23]、菊花心江蓠(G.lichenoides)[38]、江蓠(G.corticata)[39]、褐藻半叶马尾藻(Sargassumhemiphyllum)[40]等其他大型海藻研究中的结果一致。推断原因可能是由于降雨量增加或生产、生活污水排放导致海水中营养盐增加,促使藻体的生长速率和光合效率提高[41-42]。叶绿素参与光能的吸收传递和原初光化学反应,直接影响藻类的光合作用,是最重要的一类光合色素分子。本研究结果显示,叶绿素a和叶绿素b含量随着盐度增加呈现出与相对生长速率相反的变化趋势,先降低后升高,在LS处理组最高,在MS处理组最低。分析原因在于“光合色素经济性”,即光合色素合成减少,节省的能量则用于藻体的其他生物合成和代谢途径,从而提高其生长速率。藻类通过调整自身光合色素的合成来应对环境变化是一种较为普遍的现象[43-44]。
本研究结果表明,光照强度和盐度对浒苔幼苗的相对生长速率、最大光合速率、光合作用的光能利用效率、暗呼吸速率、光补偿点和最大相对电子传递速率及叶绿素a、叶绿素b、类胡萝卜素含量存在显著的交互作用(p<0.05)。在LLMS培养条件下,相对生长速率、最大净光合速率、光合作用的光能利用效率、有效光合量子产量、最大相对电子传递速率达到最高值,表明在低光强条件下,盐度的适当降低明显促进了浒苔幼苗的生长和光合作用。此外,在MS培养条件下,低光强处理组浒苔幼苗的相对生长速率、最大净光合速率、最大相对电子传递速率显著高于高光强处理组(p<0.05),说明盐度适当降低条件下,高光强对浒苔幼苗的生长和光合作用存在一定的抑制作用,这也可能是浒苔幼苗适应低盐环境的一种调节机制。
4 结论
育苗厂进/排水渠中水环境变化较为剧烈,易受到生产、生活污物和污水及降雨的影响,导致盐度和光照强度等非生物因素变化大且快。本文研究光照强度和盐度对浒苔微观繁殖体幼苗的影响,结果显示,光照强度、盐度及二者交互作用对浒苔微观繁殖体幼苗的生长和光合生理指标存在显著影响,低光强条件下盐度适当降低有利于浒苔幼苗的生长。因此,浒苔绿潮早期暴发规模的大小,一定程度上取决于光照强度与海水盐度的变化及耦合作用,可为浒苔绿潮早期暴发的预警防治提供科学依据。
