南沙珊瑚礁区波纹钩鳞鲀(Balistapus undulatus)食性和营养级分析
2022-02-17张琛胡思敏林先智周天成
张琛, 胡思敏, 林先智, 周天成, 黄 晖,4, 刘 胜
1. 中国科学院南海海洋研究所, 中国科学院热带海洋生物资源与生态重点实验室, 广东省应用海洋生物学重点实验室, 广东广州 510301;
2. 中国科学院大学, 北京 100049;
3. 三亚海洋科学综合(联合)实验室, 海南省热带海洋生物技术重点实验室, 三亚中科海洋研究院, 海南 三亚 572000;
4. 海南三亚海洋生态系统国家野外科学观测研究站, 中国科学院海南热带海洋生物实验站, 海南 三亚 572000
珊瑚礁鱼类作为珊瑚礁生态系统的重要组成部分, 为珊瑚礁生态系统的能量流动和物质循环做出了重要贡献。不同食性鱼类在物质流通中的作用不同, 故可以按照食性将珊瑚礁鱼类分为不同功能群体, 如肉食性、植食性、杂食性和碎屑食性等。鳞鲀科(Balistidae)鱼类是杂食性鱼类的代表之一, 兼具植食性和肉食性鱼类的食谱, 甚至以碎屑为食,其食性也会随着栖息地或发育阶段的改变而发生转变(Von Schiller et al, 2000)。其中, 波纹钩鳞鲀(Balistapus undulatus)是广泛分布于印度-太平洋海域的鳞鲀科鱼类, 常出现于珊瑚丰富的潟湖和向海礁区, 栖息水深范围为2~50m。Bean(2002)使用潜水调查法对5种鳞鲀科鱼类( 黑边角鳞鲀、波纹钩鳞鲀、毒吻棘鲀、颈带多棘鳞鲀、黄鳍多棘鳞鲀)的分布、丰度和微生境利用进行了量化, 发现波纹钩鳞鲀在所有调查生境中出现频率和丰度最高, 能够利用的微生境类型(如岩礁、珊瑚、藻坪、沙地等)也最多, 因此认为波纹钩鳞鲀是大尺度生境上的广布种。摄食行为是鱼类从外界获取能量的唯一途径, 并且其能够通过捕食作用影响被捕食生物的种群数量(洪巧巧 等, 2012)。Randall 在1967 年通过胃容物鉴定发现鳞鲀 科妪鳞鲀(Balistes vetula)的主要猎物是石灰海胆(Diadema antillarum), 但是当生境中没有石灰海胆时, 妪鳞鲀的摄食对象会发生改变, 从主要摄食棘皮动物变为甲壳动物(短尾类)、软体动物和棘皮动物(Menezes, 1979; Reinthal et al, 1984; Von Schiller et al, 2000)。波纹钩鳞鲀与妪鳞鲀同属鳞鲀科, McClanahan(1995) 认为波纹钩鳞鲀具有一定的领域性, 是珊瑚礁上海胆的主要捕食者之一。根据鱼类对梅氏长海胆(Echinometra mathaei)捕食行为的观察和记录发现, 波纹钩鳞鲀 占65%(McClanahan,1995, 2000; Young et al, 2012)。但到目前为止, 对波纹钩鳞鲀食物组成情况的研究还比较缺乏, 故无法准确描述其在珊瑚礁食物网中的生态地位和功能。
研究鱼类摄食常通过分析其胃/肠内含物来确定其食物组成, 以往经常使用传统的内容物镜检法来观察食物组成(Choat et al, 2002; Kulbicki et al,2005), 这种方法为将物种划分到不同的营养类群提供了一个基准。但此方法也有一定的局限性, 如工作量大, 对研究人员的分类专业知识要求高, 以及对于易消化或特征不明显的食物无法辨别等, 由此可能导致无法准确得到鱼类的完整食物组成信息。碳、氮稳定同位素技术也被广泛应用于食物链营养级(Teixeira et al, 2021), 能够反映生物相对长期的摄食情况, 但通常难以确定消费者摄食的饵料种类。基于特异性DNA 片段扩增的高通量测序技术(high-throughput sequencing technology)目前已被广泛使用于海洋生物的食物组成研究中, 如刺参(Zhang et al, 2016)、金钱鱼 (Lin et al, 2018)、龙虾(O'Rorke et al, 2012)、塔形马蹄螺(周天成 等, 2020)等。相比于传统的形态学鉴定, 该技术对生物食物组成成分的有效鉴定能力显著提升, 可以更全面地分析动物食谱, 能够发现许多容易被忽视的饵料生物。在对鱼类食谱进行分析时, 综合使用两种或几种方法可以更全面且准确地了解研究对象的摄食情况, 获得更为全面、可靠的食谱信息。
南海海域处于印度-西太平洋区域的印度-马来西亚的中心, 生境复杂多样, 是世界上热带生物种类最丰富的区域之一(赵焕庭 等, 2017)。南海诸岛海域记录的鱼类有558 种(国家水产总局南海水产研究所, 1979), 其中珊瑚礁鱼类有200 多种(陈真然,1996)。但以往的研究主要关注珊瑚礁鱼类的多样性和分布, 对它们的生态功能知之甚少。本文通过特异性引物扩增波纹钩鳞鲀消化道样本中的线粒体细胞色素C 氧化酶亚基 Ⅰ( CO Ⅰ)基因, 构建高通量测序文库, 并对COⅠ序列进行分析, 准确鉴定波纹钩鳞鲀的食物组成。此外, 通过稳定同位素技术测定并分析其碳、氮同位素, 计算其营养级, 结合两种方法推测其食物来源、营养位置以及在珊瑚礁生态系统中的生态功能。
1 材料与方法
1.1 样品采集
本研究于2017 年夏季在南沙群岛珊瑚礁海域进行了调查与采样, 研究区域范围(112°50′—114°16′E, 8°54′—9°42′N)如图1 所示。采用潜水和网捕的方式采集鱼类, 捕获的鱼类样品立即带回实验室置于–20℃保存。
图1 研究区域(红色矩形)(a)及 波纹钩鳞鲀自然生境情况(b)图a 基于由自然资源部监制的审图号为GS(2016)1554 号的标准地图制作, 比例尺为1:100000000Fig. 1 Study area (red rectangle; a) and a picture of B.undulatus (b)
1.2 样品处理
本次研究 共采集并解剖波纹钩鳞鲀(Balistapusundulatus)7 尾, 平均体重 119.6±42.6g, 平均体长13.2±1.6cm(表1)。提 取波纹钩鳞鲀的消化道内容物,使用试剂盒(FastDNA® Spin Kit for Feces, MP Biomedicals, Santa Ana, USA)从中提取DNA, 溶于30μL Tris-HCl(10mmol·L–1, pH8.0), DNA 纯度和含量使用分光光度法检测(Thermo NanoDrop ND-2000;Gene Company Limited, Waltham, Massachusetts,USA), DNA 于–20℃冷冻保存。
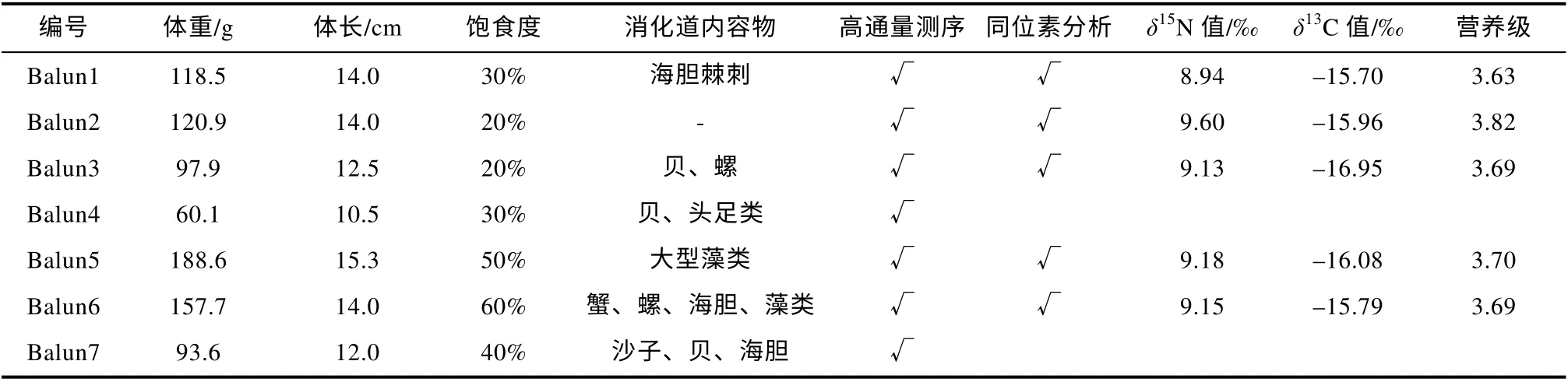
表1 波纹钩鳞鲀样品信息Tab. 1 Information on B.undulatu samples
解剖时取鱼类背部白肌用于同位素分析, 使用真空冷冻干燥机冷冻干燥肌肉48h, 研磨样品并装入样品管中, 干燥保存后送至中国科学院南海海洋研究所仪器设备公共服务中心进行检测, 利用元素分析仪-稳定同位素质谱仪(Delta V)联机测定样品中的TOC、TN、δ13C 和δ15N 值。载气He 流速200mL·min–1, 反应管温度950℃, 还原管温度600℃, 使用实验室碳氮同位素标准物质(Acetanilide#1:δ13C=-29.53‰,δ15N=1.18‰)。
1.3 高通量测序
使用一组通用的 PCR 引物 mlCOIintF/jgHCO2198(Geller et al, 2013, Leray et al, 2013), 从每个肠道内容物样本中扩增出线粒体细胞色素c 氧化酶亚基I(COI)区的313bp 区域。20μL 扩增体系包括0.4μL FastPfu 聚合酶、2μL 2.5mmol·L–1dNTPs、4μL 5×FastPfu 缓冲液、5μmol·L–1的正反引物各0.8μL、10ng DNA 模板。PCR 扩增程序设定为: 94℃预变性5min; 94℃变性30s, 57℃退火参数30s, 72℃延伸30s, 72℃终延伸参数10min, 共进行25 个循环。
PCR 产物经 2%琼脂糖电泳检测后, 使用IlluminaHiSeq 仪器(Illumina, San Diego, CA, USA)进行测序。测序得到的正反向序列首先进行两两拼接, 过滤拼接结果, 保留序列长度大于200bp 的序列。经过质量过滤, 去除嵌合体序列等, 最终得到的序列用于OTU 聚类(operational taxonomic units)。
1.4 数据分析
基于有效序列按97%的相似度进行OTU 聚类划分, 每个OTU 选择一条代表性序列, 使用RDP classifier 对代表序列进行物种分类注释。将OTU 代表序列与GenBank 数据库进行比对, 使用匹配度最高的序列, 与OTU 代表序列排列(CLUSTAL W), 剔除掉波纹钩鳞鲀自身序列后, 使用MEGA X 中的Maximum Likelihood 法并以大鼠(Rat sp.)为外群构建系统进化树, Bootstrap 值设置为1000 次, 使用Kimura 2-parameter Model 进行建树。
1.5 稳定同位素测定及分析
波纹钩鳞鲀肌肉碳、氮稳定同位素比值使用稳定同位素质谱仪测定。使用δ值表示碳、氮稳定同位素比值,δ13C 和δ15N 的计算公式如下:
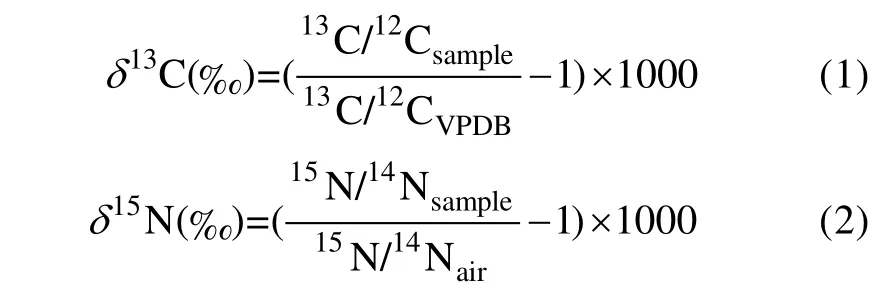
式中:13C/12CVPDB是国际标准物(VPDB)的碳稳定同位素比值;15N/14Nair是大气氮(N2)的氮稳定同位素比值。δ13C 值的精度为±0.15,δ15N 值的精度为±0.18。
营养级使用δ15N 值计算, 公式如下:

式中: TLfish表示波纹钩鳞鲀的营养级;δ15Nfish表示波纹钩鳞鲀的氮稳定同位素比值;δ15Nbaseline表示所选取的基线生物δ15N 值, 本研究选取研究区域中使用浅海Ⅱ型浮游生物网所采集的浮游动物为基线生物, 经测定, 其δ15N 平均值为3.4‰; Δ15N 为一个营养级的氮稳定同位素富集度, 本研究取 3.4‰(Minagawa et al, 1984); “2”为基线生物(浮游动物)的营养级。
2 结果与分析
2.1 高通量测序数据
测序结果显示, 共获得总有效序列53807 条,去除波纹钩鳞鲀自身序列, 剩余有效食物序列共30348 条, 占总有效序列的56.4%。当序列的相似度≥98%时, 将其划分到种水平; 相似度低于98%时,将其归为更高的分类阶元; 序列相似度低于90%的则定为未鉴定真核生物(Unclassified Eukaryota)。根据比对结果, 有41.18%的OTU 鉴定到种, 29.41%鉴定到属, 鉴定为科及科以上的OTU 占29.41%(图2)。
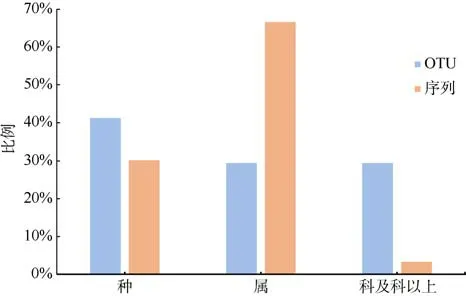
图2 波纹钩鳞鲀食物序列和OTU 在各分类水平的比例Fig. 2 The proportion of food sequence and OTU(operational taxonomic units) in different taxonomic levels of B.undulatus
2.2 波纹钩鳞鲀的食物组成
为了确保结果的准确度, 去除波纹钩鳞鲀 自身序列、数量较少的序列和无法识别的序列后, 剩余17 个OTU, 分属于3 个门类: 节肢动物门(Arthropoda)、脊索动物门(Chordata)、环节动物门(Annelida)。节肢动物门是食物序列中最丰富的类群, 占总食物序列的75.6%, 包含12 个OTU。食物序列中大部分(70.3%)为扇蟹科(Xanthidae)的蟹类, 如滑面蟹(Etisussp.)、Luniella pubescens、皱纹花瓣蟹(Liomera rugata)等, 分别占61.8%、6.7%和1.8%, 此外还发现了少量隐白硬壳寄居蟹(Calcinus latens)的序列(1.1%), 以及虾类(4.2%), 如鼓虾(Alpheidaespp.)、岩虾(Periclimenessp.)、藻虾(Hippolytidaespp.)等(图3)。
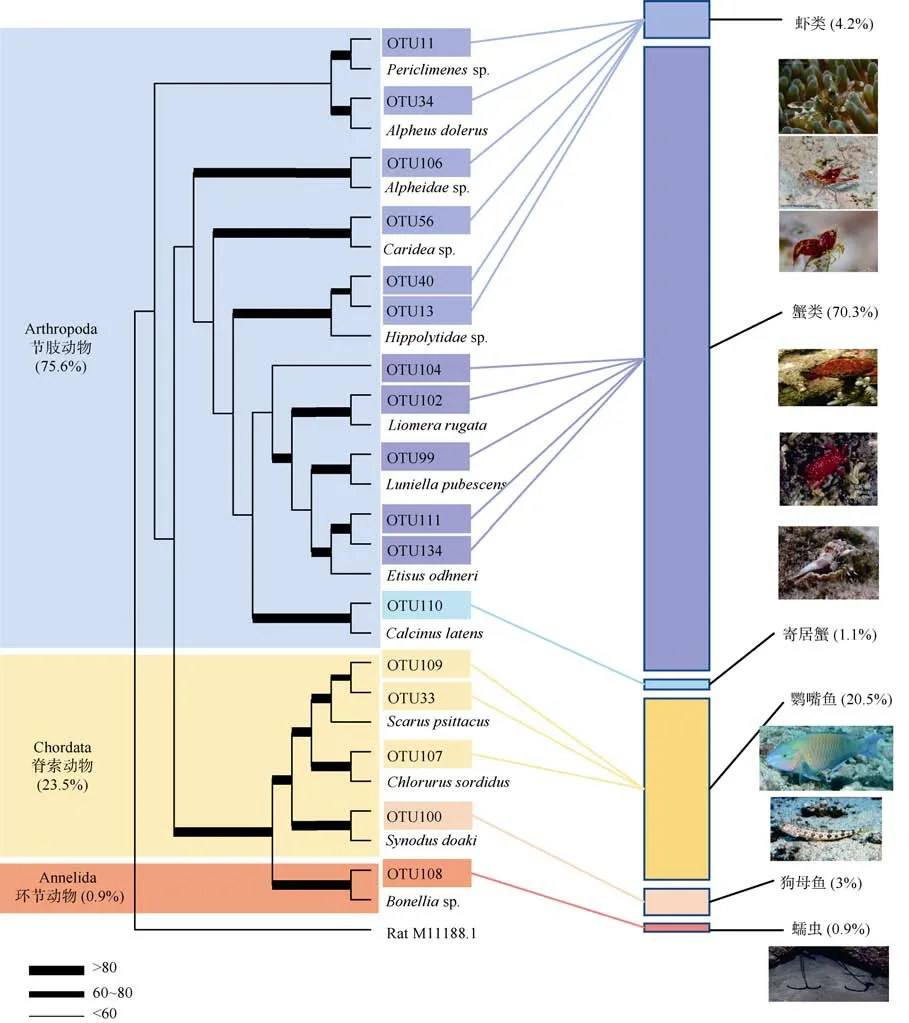
图3 波纹钩鳞鲀的食物分布与组成Fig. 3 Dietary partitioning and composition of B.undulatus
脊索动物占总食物序列的 23.5%, 包含 3 个OTU, 分别为17.5%的隆头鱼科(Labridae)棕吻鹦嘴鱼(Scarus psittacus)、3%的污色绿鹦嘴鱼(Chlorurus sordidus)以及3%的狗母鱼科狗母鱼(Synodussp.)。食物序列中还检测到了海生蠕虫, 占0.9%, 包含1个OTU, 为环节动物门后 螠属(Bonelliasp.)的生物。
综上, 波纹钩鳞鲀的食物序列在门水平(phylum)上, 总食物序列数的大部分(75.6%)被节肢动物门所占据, 此外鱼类占23.5%, 还有少量的蠕虫占0.9%。其食物序列及所占比例显示, 波纹钩鳞鲀的主要饵料生物是小型甲壳类和鱼类等。
2.3 波纹钩鳞鲀的碳、氮同位素比值及营养级
对所采集的部分波纹钩鳞鲀(n=5)进行了稳定同位素分析(表1), 样品平均体重为136.70±36.10g,平均体长为 14.0±1.0cm;δ15N 值范围为 8.94‰~9.60‰, 平 均 值 为(9.20±0.24)‰;δ13C 值 范 围 为–16.95‰~–15.70‰, 平 均 值 为(–16.10±0.50)‰(表1)。根据营养级计算公式, 利用δ15N 值计算得到波纹钩鳞鲀的营养级范围为3.63~3.82, 营养级均值为3.71±0.07。
3 讨论
鱼类是珊瑚礁生态系统中的重要组成部分和顶级捕食者(Arai, 2015), 而摄食分析是一种可以深入了解珊瑚礁鱼类生态作用的有力方法, 但一直以来其低分辨率的食物分类结果往往掩盖了营养联系和摄食策略的复杂性。目前使用分子标记方法的摄食研究日趋增多, 该方法解决了以往的传统胃容物鉴定法分辨率低, 以及对消化程度高的食物无法鉴别的问题, 有效地提高了分类的分辨率, 使得我们能够更加准确地发现复杂的营养联系和摄食策略。本研究使用了一组通用的 PCR 引物——mlCOIintF/jgHCO2198, 其鉴定结果比使用传统的DNA 条形码标记引物LCO1490/HCO2198 要更精确(Geller et al,2013; Leray et al, 2013)。此外, Leray 等(2013)对6 种不同的引物组合的实验分析发现, mlCOIintF/jgHCO 2198 的扩增效果最好, 其扩增成功率达到了91%。本研究共检测到13 种波纹钩鳞鲀的饵料生物, 主要为小型甲壳类和鱼类, 包括以往相关研究中没有被检测到的海洋蠕虫, 扩展了以往对其海胆捕食者的认识。
3.1 波纹钩鳞鲀的食物来源及营养级
根据 FishBase (https://www.fishbase.in/search.php)的描述, 波纹钩鳞鲀以各种底栖生物如藻类、棘皮动物、软体动物、海绵以及鱼类等为食, 属于杂食性鱼类。本研究结果与Von Schiller 等(2000)的结果相似, 在检测到的波纹钩鳞鲀食物类群中, 短尾类是主要的食物来源之一, 并且得益于测序技术的高分辨率, 确定了主要种类为扇蟹科滑面蟹(Etisussp.)、Luniella pubescens、皱纹花瓣蟹(Liomera rugata)等。扇蟹是印度-西太平洋区珊瑚礁最常见的蟹类之一, 大都生活在潮间带或沿岸的浅水区, 喜藏匿于岩石缝隙和珊瑚枝杈之间, 同时也是珊瑚礁小型甲壳类的主要物种之一。本文的食物序列中还检测到了隐白硬壳寄居蟹(Calcinus latens), 这是在前人相关研究中并未被发现的物种。
本文 在波纹钩鳞鲀的食物序列中还检测到了较为丰富的虾类, 主要种类为真虾类(Carideaspp.)、鼓虾(Alpheidaespp.)、岩虾(Periclimenessp.)、藻虾(Hippolytidaespp.)等。鼓虾是非常常见的礁栖生物,是重要的珊瑚共栖或共生物种, 能够通过帮助珊瑚抵御敌害、清理沉积物等行为促进珊瑚的健康和生长; 而长臂虾科的岩虾通常为清洁虾, 在热带珊瑚礁中广泛分布, 且常与鱼类、珊瑚、棘皮动物和海绵动物等共栖或共生生存; 藻虾多数种类为浅海生活的小型虾, 喜欢与海藻或海葵等共栖, 隐藏于海滩石块下、珊瑚礁中或海草丛间。海洋蠕虫也在波纹钩鳞鲀的食物序列中出现, 为 螠虫科(Bonelliidae)的后 螠(Bonelliasp.)。
波纹钩鳞鲀另一个主要的食物来源是鱼类, 共检测到 3 个种类: 有植食性棕吻鹦嘴鱼(Scarus psittacus)、污色绿鹦嘴鱼(Chlorurus sordidus)以及肉食性的狗母鱼(Synodussp.)。鹦嘴鱼是健康珊瑚礁区广泛分布的植食性鱼类, 主要以底栖藻类为食, 它们能够通过摄食行为缓和藻类与珊瑚的空间竞争,并且鹦嘴鱼在摄食过程中还会将沉积物从觅食的礁区移至礁外围, 因此是对珊瑚健康有利的植食性鱼类。McClanahan(1995, 2000)认 为波纹钩鳞鲀有一定的领域性, 它们攻击性较强, 会杀死并食用比自身小的生物, 而根据本文波纹钩鳞鲀的样品大小推测,其摄食的鹦嘴鱼可能是仔稚鱼或幼体。以上结果说明, 波纹钩鳞鲀的主要食物是小型甲壳类、植食性鹦嘴鱼等初级消费者, 应该处于较高营养级(3~4)。根据稳定同位素分析结果显示, 波纹钩鳞鲀的营养级均值为3.71±0.07, 表明本文研究方法所反映的食物组成是可信的。尽管本文研究结果与Reinthal 等(1984)和Von Schiller 等(2000)的研究结果相似, 但由于本研究样品数量较少, 故无法推断波纹钩鳞鲀是否与妪鳞鲀有类似的食物组成变化情况。本研究所采集的 波纹钩鳞鲀均为成体, 据Bean(2002)的描述, 波纹钩鳞鲀幼体和成体占据的微生境底质类型有所差异, 成体倾向于生活在岩礁、叶状珊瑚和枝状珊瑚区域, 而幼体对藻坪、海绵、沙地、大型藻类、皮壳状珊瑚和软珊瑚等环境更为偏爱。本研究未检测到大型藻类, 可能与成体波纹钩鳞鲀的栖地选择和食物偏好有关, 其幼体和成体在个体发育过程中是否存在食物选择的变化尚不能确定, 这有待今后进一步研究和探讨。
3.2 波纹钩鳞鲀的生态功能探讨
研究结果显示, 波纹钩鳞鲀主要摄食底栖甲壳类动物, 这些甲壳类动物大多与珊瑚共生或共栖,可能会对石珊瑚的健康产生直接的正面或负面作用(Stella et al, 2011)。例如, 有些无脊椎动物如扇蟹科黑指波纹蟹(Cymo melanodactylus)可以帮助珊瑚抵御疾病的发展(Pollock et al, 2013), 鼓虾(Alpheus lottini)和梯形蟹(Trapeziaspp.)等可以直接保护珊瑚免遭以珊瑚为食的动物的侵害, 并且可以将沉积物从珊瑚宿主中移除, 以及减轻珊瑚竞争对手带来的不利影响(Stier et al, 2012; McKeon et al, 2014;Rouzé et al, 2014)。本次研究区域的珊瑚礁健康程度较低, 石珊瑚覆盖率仅为14%~16%, 主要优势种为滨珊瑚(团块滨珊瑚Porites lobata和澄黄滨珊瑚Porites lutea)和疣状杯形珊瑚Pocillopora verrucosa,由于过度捕捞和珊瑚礁退化等原因, 鹦嘴鱼数量比较稀少。本研究检测到的扇蟹和鼓虾可能具有帮助珊瑚抵御疾病、驱赶敌害以及移除沉积物的作用,寄居蟹也能够通过摄食大型藻类来为珊瑚减轻竞争压力(Altman-Kurosaki et al, 2018)。研究区域鹦嘴鱼种群数量较少, 波纹钩鳞鲀对鹦嘴鱼的摄食可能会对其种群造成影响, 从而影响到珊瑚, 但由于对小型甲壳类在研究区域的分布和丰度还不清楚, 所以无法推断波纹钩鳞鲀的摄食活动是否会因影响到小型甲壳类动物群落而对珊瑚造成间接影响。
以往研究中发现波纹钩鳞鲀主要捕食海胆(McClanahan, 1995, 2000; Young, 2012)。海胆是珊瑚礁生态系统中的植食性重要功能生物(Hughes, 1994;McClanahan, 1994)。McClanahan(1995, 2000)的研究表明波纹钩鳞鲀是珊瑚礁海域中能够控制海胆种群数量的主要捕食者之一, 其捕食行为有利于珊瑚的健康和生长。本研究在样品解剖过程中, 于少数(n=3) 波纹钩鳞鲀的肠容物中发现了残留的海胆棘刺, 因棘刺无法作为有效测序成分, 故测序结果中并未发现海胆序列。但同时分子方法带来的高分辨率鉴定结果显示, 一些可能对珊瑚健康有益的小型甲壳动物也是波纹钩鳞鲀的主要食物来源, 因此本文研究结果丰富了波纹钩鳞鲀的食谱, 提高了人们对珊瑚礁生态系统营养联系的认识。
4 结论
南沙海域波纹钩鳞鲀的食物来源主要为底栖小型甲壳动物、鱼类以及海洋蠕虫, 其平均营养级为3.71±0.07。波纹钩鳞鲀摄食的扇蟹、鼓虾、寄居蟹等小型甲壳动物以及鹦嘴鱼, 大多是对珊瑚健康有利的珊瑚共栖或共生动物。研究结果拓展了以往对波纹钩鳞鲀所扮演生态角色的认识, 展现了礁栖生物之间营养联系的复杂性。
