西双版纳不同海拔4种优势树种的叶特性研究
2022-02-16栗忠飞赵晓鹏李雪楠
孙 靖,栗忠飞,屠 晶,赵晓鹏,李雪楠
西南林业大学 生态与环境学院,云南昆明 650224
植物的功能性状是其对生存环境适应策略的集中体现。功能性状的环境适应最大限度地反映了植物的最优环境适应原则[1]。在植物各个器官中,叶片是植物对环境变化感知最为敏锐的部分,叶的形态、结构等随环境变化具有较高的可塑性[2]。植物叶特性随环境变化而发生的改变会进一步体现到植物物种乃至群落尺度上生物量、生产力的变化[3],并在一定程度上反映植物所处环境的历史气候特征。因此,植物叶片功能性状及其与群落生物量、生产力,以及气候环境之间的耦合关联机制等受到了广泛的关注[4]。
植物叶特性中,比叶面积(Specific Leaf Area,SLA)是叶片结构型形状的重要指标[5],是植物资源利用效率、环境适应策略的重要体现,对植物生长速率具有显著影响[6]。植物在特定的环境条件下通过最优化叶干物质量、叶面积大小、薄厚等性状特征,调节其光合、蒸腾等生理生态过程,提高对环境的适应性[7]。群落中,物种相对高的生长速率及其环境适应能力会直接影响群落的物种更新及其结构组成,这些环境适应策略最终也会体现到植物群落的生物量、生产力、碳汇潜力等生态功能上[8~9]。生物量反映植物生长与生产力的重要特征,植物可通过改变生物量来适应环境的变化[10]。叶生物量作为衡量与评价植物叶特性的重要指标,具有很好的研究价值[11]。
海拔是一个重要的环境因素,海拔的变化会引起各种环境因子的变化(温度、降水、光照等),为研究局地尺度上植物对环境及气候变化的响应机制提供了理想的自然条件[12~13]。研究表明,叶特性对海拔梯度变化有一定的响应,许多植物的叶面积(LA)随着海拔升高而减小,但是受其他环境因子的影响,也会出现增大或先增大后减小的情况[14]。海拔高度的增加会使降雨量增加,有研究表明,植物叶特性受降水量的影响在区域与种间上表现一定的差异性,表现出植物叶特性和区域降水量呈现正相关、负相关以及不相关的关系[15]。温度会随着海拔的升高而降低,进一步影响植物的生理生态特性。温度增高会导致一些植物叶面积减小,高温会缩短植物的叶寿命;但也有研究指出,一些植物叶特性与温度并无显著相关[16]。
幼苗是森林生态系统的重要组成,对森林群落结构的完整性与物种更新有着重要作用[17]。目前,针对植物移植后对环境适应性的研究有很多[18],但对植物幼苗阶段对不同环境的适应性研究较少。绒毛番龙眼、云南玉蕊、云南肉豆蔻以及小叶红光树是热带季节雨林中的优势树种,在热带季节雨林的结构与功能上占据重要位置。
本研究选取上述4种优势种树种,在3个海拔梯度上开展移植实验,观测4种幼树叶特性随海拔梯度的变化特征,分析海拔对植物叶特性的影响,为气候环境变化下热带季节雨林树种特性的响应机制提供科学依据。
1 材料与方法
1.1 研究区概况
试验样地位于西双版纳勐仑(600 m)、菜阳河(1 100 m)、勐宋(1 600 m)3个不同海拔高度上。
勐仑试验地位于西双版热带植物园的葫芦岛(21°09′N~22°36′N,99°58′E~101°50′E),该地区气候类型为印度洋季风气候,主要的植被类型为热带季节雨林。
菜阳河试验地位于菜阳河自然保护 区(101°7′E~101°15′E、22°30′N~22°38′N),在云南思茅东南部,气候类型为印度洋季风气候,主要植被类型是季风常绿阔叶林。
勐宋试验地位于西双版纳西南端的中缅边境勐宋地区(100°25′E~100°35′E,21°27′N~21°34′N),属典型的南亚热带气候区,气候类型为印度洋季风气候,雨量充沛,全年干湿季明显,冬季有轻霜,主要植被类型是热带山地雨林。具体气象数据见表1。

表1 3个海拔梯度上的气象数据
1.2 研究方法
1.2.1 物种选择本试验选取热带季节雨林优势种绒毛番龙眼(Pometia tomentosa)、云 南 玉 蕊(Barringtonia macrostachya)、云南肉豆蔻(Myristica yunnanensis)和小叶红光树(Knema globularia)幼树作为试验物种。
1.2.2 幼苗移植实验选择上述4个物种的种子,在装有菜园土的育苗袋中培育,当幼苗生长到20 cm高度时,移植到上述3个海拔梯度的实验样地上,即勐仑(600 m)、菜阳河(1 100 m)和勐宋(1 600 m)样地,分别定义为低、中、高海拔实验样地。移植后,抚育管理1年后进入自然生长阶段,3年后采集移植树种的叶片,每个海拔上每种幼树选取3株植株,每株上取形态结构优良的叶片10片,测定其单叶面积、干物质重等,同步测定植株树高、基茎、生物量等参数。
1.2.3 叶特性指标测定用LI-3000C叶面积仪,测定叶片面积,用天平测定每片叶子的湿重,后将叶片装进牛皮纸袋中并编号,置于烘箱中杀青后烘干至恒重,测定其干重(叶生物量)。
比叶面积(SLA)=总叶面积/叶干重(cm2/g)
1.2.4 数据处理用Excel 13.0、SPSS 25.0和Origin 2019对试验所得数据进行统计分析及绘图。
2 结果与分析
2.1 4种优势树种SLA在不同海拔下的差异
绒毛番龙眼的SLA随海拔的升高而增大,尤其是在高海拔上显著大于中、低海拔,达到97.8 cm2/g(P<0.05)。云南玉蕊在中海拔上表现最高,为142.1 cm2/g,在高海拔上显著下降,为116.4 cm2/g(P<0.01)。另外2种幼树SLA均表现出随海拔的升高而显著降低的特征(图1)。

图1 4种优势树种在不同海拔下的比叶面积的差异
2.2 4种优势树种SLA在相同海拔下的差异
600 m海拔SLA以小叶红光树为最高,平均达到133.2 cm2/g,绒毛番龙眼最低,仅为84.4 cm2/g;1 100 m海拔SLA以云南玉蕊为最高,平均达到142.1 cm2/g;云南肉豆蔻最低,仅为84.5 cm2/g;1 600 m海拔SLA以云南玉蕊为最高,平均达到116.4 cm2/g,云南肉豆蔻最低,仅为61.2 cm2/g(图2)。
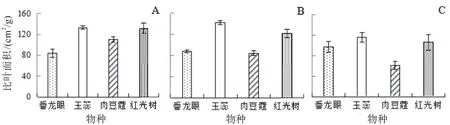
图2 4种优势树种在相同海拔下的比叶面积的差异
2.3 4种优势树种SLA随海拔、温度的变化
绒毛番龙眼(图3A、图4A)SLA随海拔、温度变化差异显著,随海拔的升高而增大(P<0.05),1 100 m海拔时SLA达到最大,随温度降低而减小(P<0.05),22.4℃时SLA最小。云南玉蕊(图3B、图4B)SLA随海拔、温度先增大后减小(P<0.01),在1 100 m海拔、19.8℃时,SLA达到最大。云南肉豆蔻(图3C、图4C)SLA随海拔的升高而减小(P<0.001),在1 600 m海拔时SLA最小,随温度升高而增大(P<0.001),22.4℃时SLA达到最大。小叶红光树(图3D、图4D)SLA随海拔的升高而减小(P<0.05),在1 600 m海拔时SLA最小,随温度升高而增大(P<0.05),22.4℃时SLA达到最大。
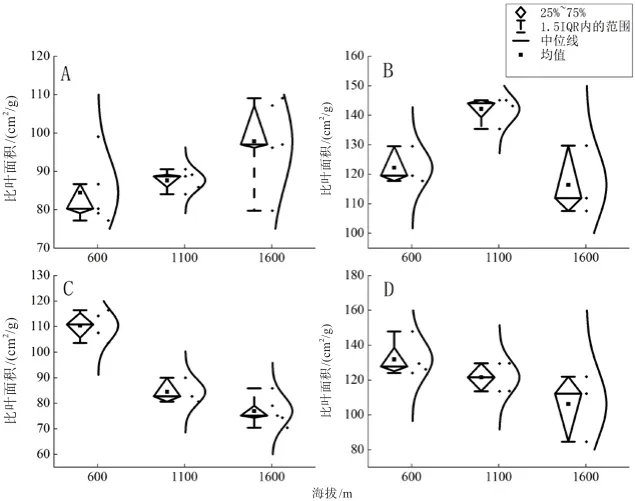
图3 4种优势树种SLA随海拔梯度性变化

图4 4种优势树种SLA随温度的变化
2.4 4种优势树种叶生物量随海拔、温度的变化
绒毛番龙眼(图5A、图6A)叶生物量随海拔的升高而减小(P<0.001),在1 100 m海拔时叶生物量最小,随温度升高而增大(P<0.001),22.4℃时叶生物量达到最大。云南玉蕊(图5B、图6B)叶生物量随海拔的升高而减小(P<0.01),在1 600 m海拔时叶生物量最小,随温度升高而增大(P<0.01),22.4℃时叶生物量达到最大。云南肉豆蔻(图5C、图6C)叶生物量随海拔、温度先增大后减小(P<0.01),在1 100 m海拔、19.8℃时叶生物量达到最大。小叶红光树(图5D、图6D)叶生物量随海拔的升高而减小(P<0.001),在1 600 m海拔时叶生物量最小,随温度的升高而增大,22.4℃时叶生物量达到最大。
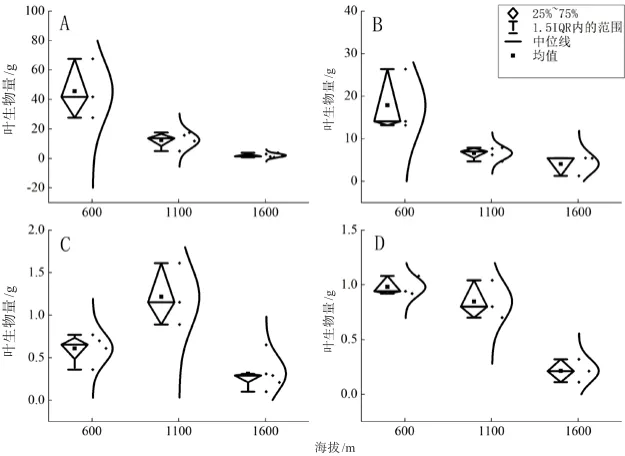
图5 4种优势树种叶生物量随海拔梯度性变化
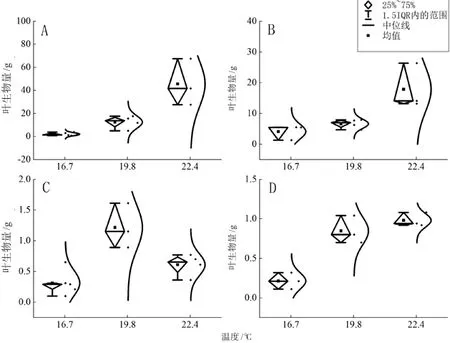
图6 4种优势树种叶生物量随温度变化
3 讨论与结论
3.1 讨论
3.1.1 4种优势树种比叶面积随海拔梯度性变化植物叶面积的变化会影响植物的光合作用、呼吸作用和蒸腾作用,与叶面积相关的植物叶片性状成为衡量群落结构稳定性的指标[19]。比叶面积可以反映植物生长与养分吸收之间的权衡[20]。一般来说,对比叶面积比较低的植物,叶片中大部分的物质都用于构建保护结构或增加叶肉细胞的密度,通常会形成厚度比较大而面积比较小的叶片,延长叶片寿命[22]。
同一物种在低海拔区域的比叶面积较高,随着海拔梯度的升高,比叶面积会逐渐减小[21]。出现这原因可能是高海拔地区土壤温度较低,使植物根系对土壤水分的吸收受到了影响,因此植物受到的水分胁迫逐渐加剧,为了适应这种状况,选择增加自身的叶片厚度、减小比叶面积,从而使水分胁迫减缓并提高抗旱性[22];另一种解释是,高海拔地区植物为了适应寒冷的环境,会减小自身的叶长与叶面积,这仅仅是植物获得更高抗寒性的一种反应[24]。在高海拔地区,植物往往具有较厚且密集的叶片,使植物能够充分吸收且利用光能,提高自身的固碳能力,而且还可以使自身对强光的适应力增强[22]。
在本研究中,云南肉豆蔻与小叶红光树的比叶面积具有明显的海拔梯度变化趋势,即2种树种的比叶面积随着海拔的增高而减小,这可能与2种植物为了应对高海拔的低温而增加叶片厚度有关。植物为了抵抗持续的低温和冬季干旱,会在叶片中合成多种抗冻蛋白,糖和脯氨酸来调节渗透压,减缓细胞的水分散失以及质壁分离,从而间接降低叶片的SLA值。绒毛番龙眼的比叶面积随着海拔的增高而增大,这是由于绒毛番龙眼的生存能力较强,1 600 m海拔处没有达到绒毛番龙眼生存的相对低温。云南玉蕊的比叶面积随海拔的增加先增大后减小,并在1 100 m处达到最大值。这种变化主要受植物的生长环境影响:在低海拔地区,气候通常是气温高、降水少。为了适应这种干燥和炎热的环境,云南玉蕊不得不增加叶肉细胞的密度以抵抗高温,从而增加单位叶面积的干物质。同时,在相对干燥和炎热的环境中,较小的叶片可以减少植物水分流失,这种对环境条件的适应性导致植物拥有较大的叶片厚度和较小的面积,所以云南玉蕊在低海拔地区叶片比叶面积较小;随着海拔的升高,温度、水分条件逐渐转优,叶片面积逐渐增大,使植物能更好地吸收水分和养分;但在海拔1 100 m左右的区域,水热综合条件达到最佳,使得云南玉蕊的比叶面积达到最大,能更好地适应自身的生长和发育;在海拔1 100 m以上的区域,温度因子逐渐成为制约云南玉蕊正常生长的主要环境因子,海拔升高后所带来的低温环境会使云南玉蕊产生抵御寒冷的生理反应,叶片生长速度减缓,比叶面积减小,云南玉蕊获得较高的抗寒性。
低温会降低植物各个器官的吸水能力,植物所受的干旱胁迫逐渐加剧,植物通过增加叶片的厚度和减少叶片的面积,使水分散失减缓,同时增加自身的抗旱性[23]。此外,一些研究认为,随着海拔和光强的增加,植物为了避免灼伤,通常会选择增加叶片厚度、减小比叶面积,避免捕获过多的光能[24]。这与本研究云南玉蕊在海拔1 100 m以上的区域比叶面积随海拔的升高而减小的结果相同。
3.1.2 4种优势树种叶生物量随海拔梯度性变化本研究中,4种优势树种叶生物量随海拔梯度都有明显的变化,绒毛番龙眼、云南玉蕊以及小叶红光树叶生物量随海拔的升高而减少,导致这类现象的原因有:第一,这3种优势树种普遍分布在低海拔地区,降水相对于高海拔较少,在高海拔水分竞争中不占据优势,被排斥所以生物量降低。第二,随着海拔升高,温度显著降低,体现了低温对植物幼苗生长的限制[25]。云南肉豆蔻叶生物量随海拔的升高先增大后减小,这与李武斌等[26]的研究结果一致,可能是因为中低海拔较为适合大部分植物的生长,随海拔的升高,生长环境逐渐变得恶劣,不适合植物的生长。
3.2 结论
(1)绒毛番龙眼的比叶面积随着海拔的升高而增大(由于1 600 m海拔没有达到绒毛番龙眼生长的相对低温)。(2)云南玉蕊的比叶面积随海拔的升高先增大后减小(对于云南玉蕊海拔1 100 m左右水热综合条件达到最佳)。(3)云南肉豆蔻与小叶红光树的比叶面积随着海拔的增高而减小。(4)绒毛番龙眼、云南玉蕊以及小叶红光树叶生物量随海拔的升高而减少(高海拔低温、降水少,抑制生物量积累)。(5)云南肉豆蔻叶生物量随海拔的升高先增大后减小(中低海拔较为适合大部分植物的生长,随海拔的升高,温度以及湿度显著降低,不适合植物的生长)。研究4种优势树种叶特性与环境因子的关系,有利于了解植物对海拔变化所带来的环境因子变化的适应策略,为研究海拔对植物叶特性的调控作用提供一定的理论依据。
