FABP2基因在不同比例花椒籽饲喂育肥猪的组织差异表达、克隆及生物信息学分析
2022-02-15黄晓宇尚学峰杨姣姣张娟丽谢开会张亚丽闫尊强王鹏飞高小莉杨巧丽马艳萍滚双宝
李 杰,黄晓宇,尚学峰,杨姣姣,张娟丽,谢开会,张亚丽,闫尊强,王鹏飞,高小莉,杨巧丽,马艳萍,滚双宝,3
(1.甘肃农业大学 动物科学技术学院,甘肃 兰州 730070;2.甘肃农业大学图书馆,甘肃 兰州 730070;3.甘肃省现代养猪技术工程研究中心,甘肃 兰州 730070)
我国作为世界第一大花椒生产国,花椒种植历史悠久,品种齐全,种植面积超过113万hm2,主产区产量超过37万t[1-2]。花椒果皮常用作调味品,花椒籽为花椒的主要副产品,占花椒总质量的60%以上[3],花椒籽油中富含脂质和蛋白质,多不饱和脂肪酸种类多且含量高[4-5]。李路路[6]研究发现,花椒籽油中多不饱和脂肪酸高达85%。随着人们生活水平的不断提高,对猪肉品质有了更高的要求,肌内脂肪、脂肪酸含量是评价肉品质的指标,直接影响着肉的风味和口感。然而,猪肉中多不饱和脂肪酸的合成需要中间产物(亚油酸、α-亚麻酸等),且胴体中的脂肪酸含量与饲料有直接关系[7]。花椒籽中富含油酸、亚油酸、a-亚麻酸、棕榈酸等脂肪酸[8],是猪从饲料中获取脂肪酸较为理想的来源,在饲料工业中具有较好的利用价值。近年来,对花椒籽的开发利用方面研究炙热,已有学者在鸡[9]、鱼[6]、猪[10]上进行试验,研究均表明,在日粮中添加一定比例的花椒籽粉可提高动物生产性能。由此可见,花椒籽可在饲料行业开发利用,它的合理利用有望节约生产成本和提高经济效益,解决花椒生产中副产品浪费的问题。
脂肪酸结合蛋白2基因(Fatty acid-binding protein 2 gene,FABP2),蛋白分子质量较小(14~15 ku),属于脂肪酸结合蛋白家族(FABPs)[11],该家族基因大多含有多态性位点,通过调节脂肪酸代谢途径来提高肉品质,其多态性位点间接影响肌内脂肪含量[12-13],FABPs的这种生物学功能对猪肉肌内脂肪沉积有潜在影响。肌内脂肪是猪肉品质评定的一项重要指标[14],其含量影响肉的风味。现阶段育种对猪肉肌内脂肪含量要求较高,如何增加肌内脂肪含量、提高肉品质,是当今育种工作者的目标和难点。而FABP2与体内脂肪沉积相关联,可作为影响肌内脂肪含量的候选基因,主要参与哺乳动物长链脂肪酸的运输和吸收,影响脂类代谢[15-16]。其次可以通过调节参与脂质代谢的基因和提高脂肪酸水溶性来促进肠道对多不饱和脂肪酸的吸收[17-18]。除此之外,FABP2基因还与胆汁酸和维生素具有较高的亲和力[19],这表明FABP2具有极其重要的生理作用。
为了寻找与猪脂肪沉积有关的基因,本试验通过研究FABP2在不同比例花椒籽饲喂育肥猪中的组织差异表达,以及对其编码区进行克隆及生物信息学分析,以期为花椒籽的开发利用和FABP2基因作为肌内脂肪沉积候选基因的相关研究提供参考依据。
1 材料和方法
1.1 样品采集
本试验所用组织样为畜牧学养猪产业技术创新团队前期试验保存[10]。试验对象为288头90日龄杜×长×大三元杂交育肥猪(33 kg),由临夏州众惠猪业科技有限公司提供,将其随机分为4组,每组设4个重复,每个重复18头猪;对照组饲喂基础日粮,各试验组分别用花椒籽代替饲粮中2.5%(试验Ⅰ组)、5.0%(试验Ⅱ组)、7.5%(试验Ⅲ组)的玉米,预试验7 d,正试期100 d。试验结束后,屠宰猪只,采集心脏、肝脏、脾脏、肺脏、肾脏、十二指肠、空肠、回肠、结肠、直肠、盲肠和背肌组织,液氮迅速冷冻,-80 ℃超低温冰箱长期保存。
1.2 试验主要试剂
TRIzol试剂购自北京全式金生物技术有限公司,Evo M-MLV反转录预混型试剂盒购自湖南艾科瑞生物工程有限公司,2×Universal Blue SYBR Green qPCR Master Mix购自武汉赛维尔生物科技有限公司,DNA凝胶回收试剂盒购自擎科生物公司(西安),2×Taq PCR Master Mix和DH5α感受态细胞购自北京天根生化科技有限公司,pMDTM19-T Vector购自宝生物工程(大连)有限公司,其他常用国产分析纯试剂。
1.3 总RNA提取及反转录
用TRIzoL提取猪各组织中总RNA,用Evo M-MLV 反转录预混型试剂盒合成cDNA,具体步骤如下:①去除基因组DNA,反应体系10 μL,其中5× gDNA Clean Reaction Mix 2 μL,Total RNA 2 μL,RNase free water 6 μL,反应条件为42 ℃,2 min;4 ℃保存。②反转录反应,步骤①反应液10 μL,5× Evo M-MLV RT Reaction Mix 4 μL,RNase free water 6 μL,总体系20 μL。反应条件:37 ℃,15 min;85 ℃,5 s;4 ℃保存。反应结束后,测定cDNA浓度,-20 ℃保存备用。
1.4 FABP2引物设计与合成
根据NCBI GenBank数据库中猪的FABP2基因参考序列(NM_001031780.1)、内参基因GAPDH参考序列(NM_001206359.1),利用Primer 5.0在线软件分别设计特异性扩增引物,由中科羽瞳生物科技有限公司合成(表1)。

表1 引物信息Tab.1 Primers information
1.5 FABP2的表达检测
以3个处理组中不同组织的cDNA为模板,采用qRT-PCR检测FABP2基因在心脏、肝脏、脾脏、肺脏、肾脏、十二指肠、空肠、回肠、结肠、直肠、盲肠和背肌各组织及各试验组的相对表达量,qRT-PCR反应总体系为20 μL:2×Universal Blue SYBR Green qPCR Master Mix 10 μL,上、下游引物各0.8 μL,cDNA 2 μL,ddH2O 6.4 μL。qRT-PCR反应条件:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火30 s,40个循环。每组设3个重复,以2-ΔΔCt法[20]计算相对表达量。
1.6 三元杂交育肥猪FABP2基因克隆及测序
PCR产物经 1%琼脂糖凝胶电泳检测,按照DNA凝胶回收试剂盒说明书进行回收纯化,将胶回收cDNA与pMDTM19-T Vector载体连接,连接产物转化到DH5α感受态细胞中,加入不含Amp的LB培养基中。摇床振荡培养1 h后,取适量菌液涂到含Amp、IPTG和X-Gal的LB固体培养基上,进行蓝白斑筛选。接着挑取单个白色菌落置于LB(含Amp)液体培养基中,摇床振荡培养4~5 h。经菌液PCR扩增,将阳性克隆菌液送至西安擎科生物公司进行测序。
1.7 生物信息学分析
本试验通过多种生物信息学软件对克隆得到的三元杂交育肥猪FABP2基因的CDS区进行分析。所用在线软件及程序见表2。
1.8 数据处理
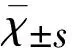
2 结果与分析
2.1 FABP2基因在三元杂交育肥猪各组织差异表达
由图1可知,FABP2在三元杂交育肥猪各组织中均有不同程度表达,在空肠中显著高表达(P<0.05),背肌中表达量最低。但是总体来说在肠道组织的表达量较高。用不同比例花椒籽替代部分玉米饲喂三元杂交育肥猪后,与对照组相比,各试验组在心脏、肺脏、肾脏、十二指肠、空肠和盲肠组织中,FABP2显著下调表达(P<0.05);脾脏和回肠组织中,FABP2在试验Ⅱ组和试验Ⅲ组中显著上调表达(P<0.05);肝脏、结肠、直肠组织中,FABP2在试验Ⅰ组显著上调表达(P<0.05);FABP2在各试验组不同组织中的表达无明显规律(图2)。

表2 生物信息学分析软件Tab.2 Bioinformatics analysis software

不同小写字母表示差异显著(P<0.05)。图2同。Different lowercase letters indicate significant difference(P<0.05).The same as Fig.2.

图2 FABP2基因在不同比例花椒籽饲喂三元杂交育肥猪的组织差异表达Fig.2 FABP2 gene was differentially expressed in tissues of DLY fattening pigs fed with different proportions of Zanthoxylum seed
2.2 三元杂交育肥猪FABP2基因CDS区PCR扩增及克隆测序
用TRIzol法提取三元杂交育肥猪回肠组织的总RNA,反转录为cDNA,以cDNA为模板进行PCR扩增,经1%琼脂糖凝胶电泳检测,所得一条427 bp的特异性条带,与预期片段大小一致(图3)。测序结果表明,FABP2基因序列全长427 bp,包括5′UTR区10 bp,3′UTR区18 bp和CDS区399 bp,编码132个氨基酸,其中152位点处的碱基A突变为G(图4),导致第51位点的赖氨酸突变为精氨酸;核苷酸序列308位点的碱基T突变为C,导致第103位点的亮氨酸突变为丝氨酸,均为错义突变(图5)。
M.DNA Marker DL2000;1,2.FABP2基因PCR扩增产物。M.DNA Marker DL2000 ;1,2.PCR amplification product of FABP2 gene.
2.3 同源性分析及系统进化树的构建
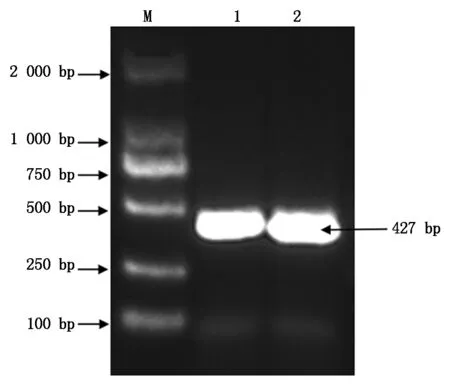
用NCBI中的Blast在线软件,分析克隆所得三元杂交育肥猪FABP2CDS区序列与其他物种CDS区序列的同源性,结果显示,三元杂交育肥猪FABP2CDS区序列与绵羊(XM_004009588.5)、马(NM_001081903.2)、牛(NM_001025332.1)、犬(XM_038444525.1)、智人(NM_000134.4)、猕猴(XM_015139143.2)、小鼠(NM_007980.3)、大鼠(NM_013068.1)、鸡(NM_001007923.2)的同源性分别为87.72%,83.33%,87.47%,88.97%,85.96%,86.97%,79.39%,81.42%,72.08%。其中,三元杂交育肥猪与绵羊亲缘关系最近,与鸡亲缘关系最远,利用MEGA 7.0软件计算不同物种间的遗传距离(表3)并构建系统进化树(图6),分析结果与同源性比对相吻合。

图4 测序结果分析Fig.4 Analysis of sequencing results

图5 氨基酸序列比对Fig.5 Alignment of amino acid sequences
2.4 三元杂交育肥猪FABP2蛋白理化性质分析
用ProtParam软件预测三元杂交育肥猪FABP2蛋白的理化性质,预测结果显示,分子式为C677H1066N186O205S4,分子质量为15.219 27 ku,原子总数为2 138,理论等电点(pI)为6.61,偏酸性;其中天冬酰胺(Asn)、谷氨酸(Glu)和赖氨酸(Lys)所占比例最高(9.80%),组氨酸(His)最低(0.80%)(图7);哺乳动物网织红细胞体外估计半衰期为30 h,大肠杆菌体内半衰期大于10 h,酵母体内大于20 h;消光系数(γ=280 nm)为16 960;脂肪族氨基酸指数为78.94,不稳定指数(Ⅱ)为25.92,表明该蛋白是稳定蛋白。
2.5 三元杂交育肥猪FABP2蛋白二三级结构预测
采用在线软件SOMPA预测三元杂交育肥猪FABP2蛋白二级结构,结果显示(图8),FABP2蛋白的二级结构主要由延伸链和无规则卷曲构成,其中延伸链占37.12%,由49个氨基酸残基组成;无规则卷曲占32.58%,由43个氨基酸残基组成;α-螺旋占18.94%,由25个氨基酸残基组成;β-转角占11.36%,由15个氨基酸残基组成。用Phyre2在线软件通过同源建模法构建三元杂交育肥猪FABP2蛋白三级结构模型,预测结果与二级结构预测一致。同时对其亲缘关系最近的绵羊和最远的鸡构建模型,三元杂交育肥猪三级结构与绵羊相比差异较小,鸡延伸链和无规则卷曲所占比例均低于三元杂交育肥猪,表明二者蛋白在三级结构上存在差异,在生物体内发挥的作用也有所不同,但三者的三级结构都主要由延伸链和无规则卷曲构成,这也是氨基酸序列保守性的原因(图9)。
表3 不同物种间遗传距离Tab.3 Genetic distance between different species

图6 不同物种FABP2基因系统进化树分析Fig.6 Phylogenetic tree analysis of FABP2 gene in different species
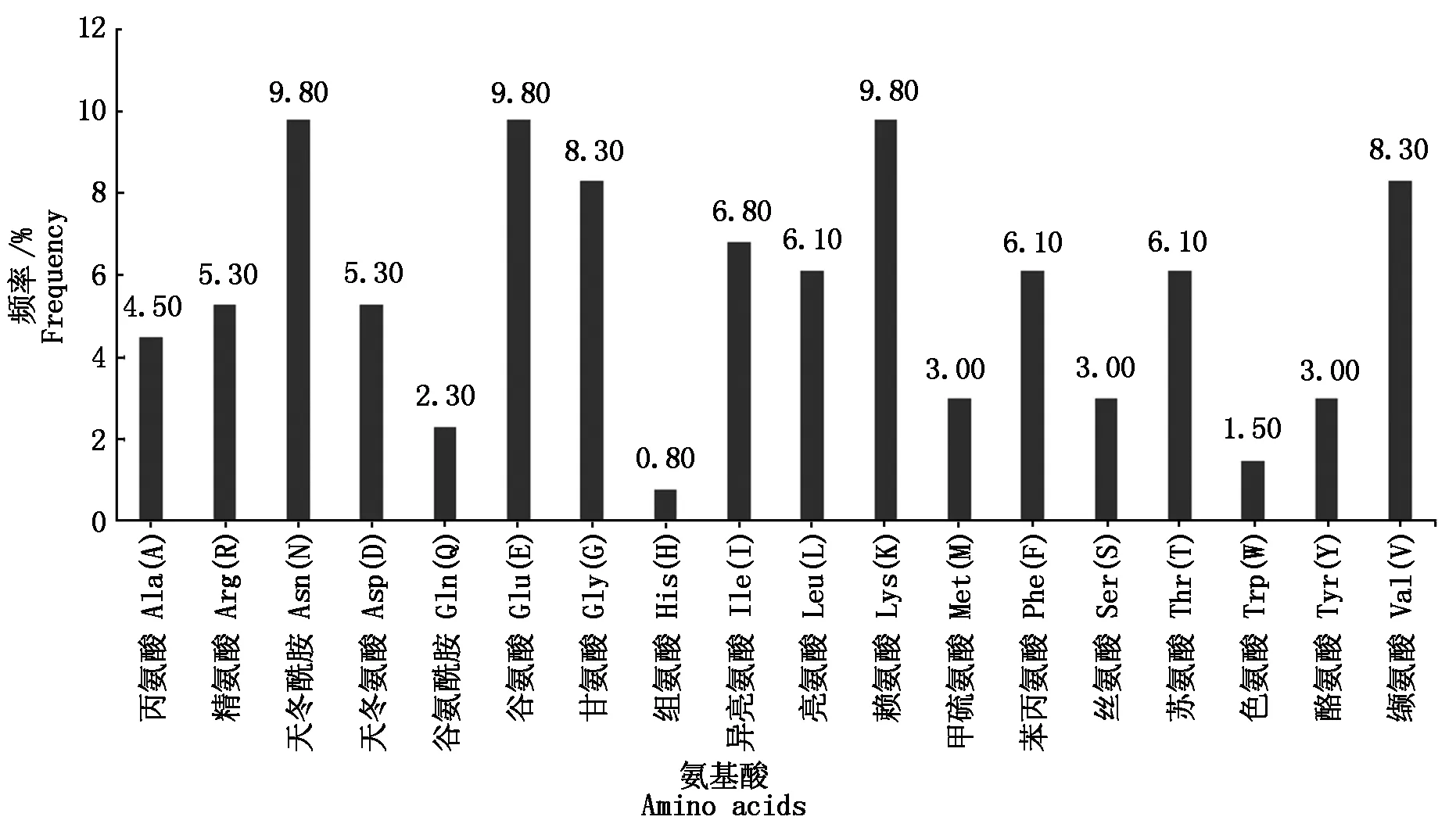
图7 氨基酸组成及频率Fig.7 Amino acid composition and frequency
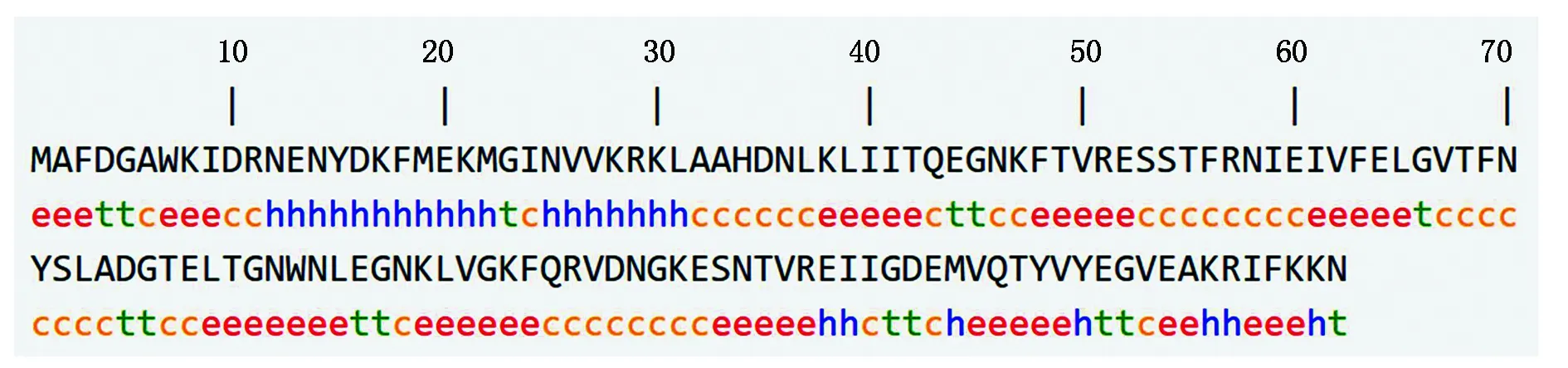
h.α-螺旋;t.β-转角;c.无规则卷曲;e.延伸链。h.α-helix;t.Beta angle;c.Random crimp;e.Extension chain.

A.三元杂交育肥猪;B.绵羊;C.鸡。A.DLY fattening pigs ;B.Sheep ;C.Chicken.
2.6 三元杂交育肥猪FABP2蛋白亲疏水性和跨膜区分析
运用ExPASy在线软件Protscale对三元杂交育肥猪FABP2氨基酸序列进行亲疏水性分析(图10-A),氨基酸序列中,第14位出现最小疏水值(-2.711),第65位出现最大疏水值(1.967),亲水性氨基酸占比(65%)高于疏水性氨基酸(35%)。可推测三元杂交育肥猪FABP2蛋白为亲水性蛋白。用在线软件TMHMM预测跨膜螺旋区域(图10-B),预测期望值为0,表明该蛋白不存在跨膜区,为非跨膜蛋白,主要在细胞质中发挥作用。
2.7 三元杂交育肥猪 FABP2 蛋白亚细胞定位和信号肽分析
用PSORT Ⅱ Prediction对蛋白亚细胞定位分析,结果表明(图11-A),该蛋白在细胞质(56.50%)、细胞核(30.40%)、线粒体(8.70%)和过氧化物酶体(4.30%)中发挥作用;SignalP 4.1预测信号肽,结果表明,信号肽S预测均值为0.104,该蛋白不具有信号肽,可推测为非分泌蛋白(图11-B)。

图10 FAPB2亲疏水性分析(A)和跨膜螺旋结构域预测(B)Fig.10 FAPB2 hydrophobicity analysis(A)and transmembrane helical structure domain prediction(B)
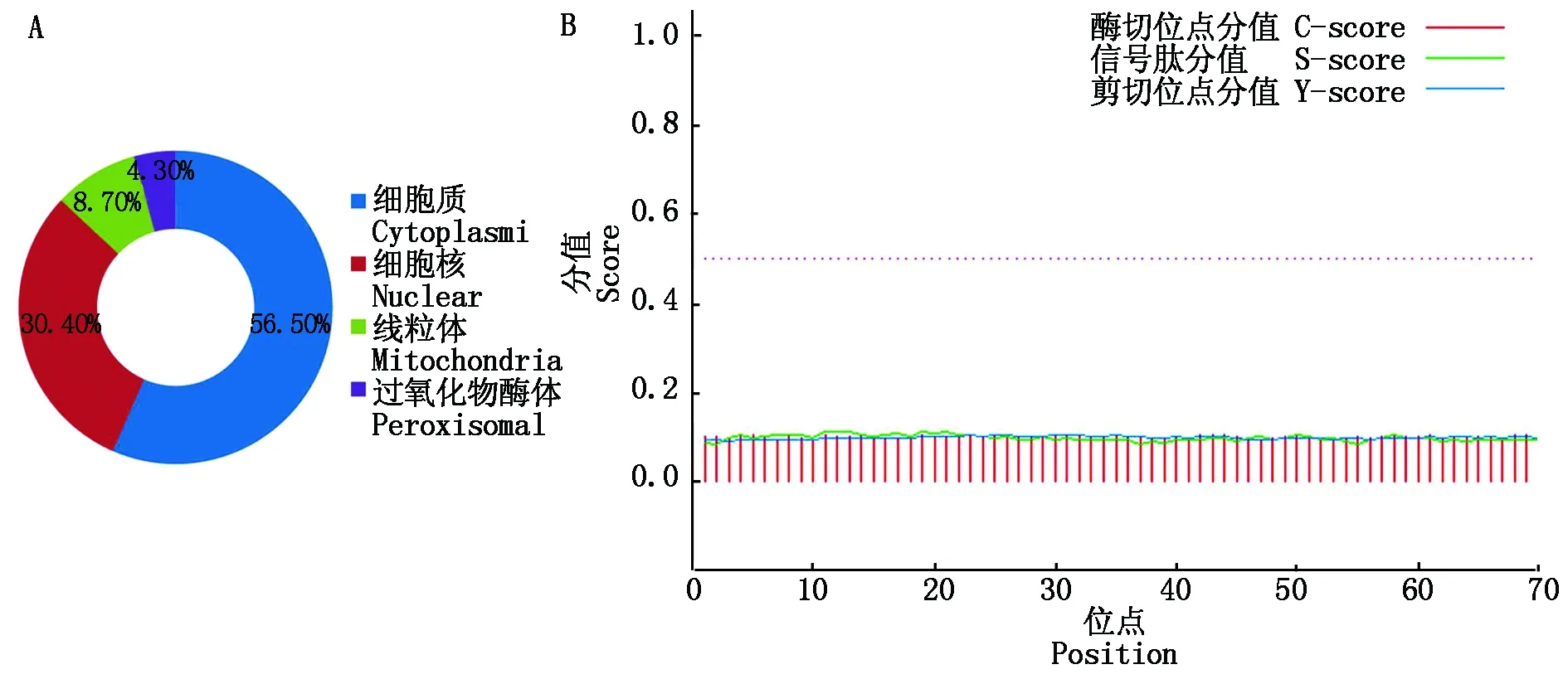
图11 蛋白亚细胞定位(A)和信号肽预测(B)Fig.11 Protein subcellular localization(A)and signal peptide prediction(B)
2.8 三元杂交育肥猪 FABP2 蛋白激酶磷酸化和N端糖基化位点分析
利用NetPhos 3.1 Server 在线软件预测三元杂交育肥猪FABP2蛋白的激酶磷酸化修饰位点(图12-A),预测到丝氨酸在53(2个),54,72,103位点出现磷酸化位点,得分分别为0.692,0.503,0.993,0.502,0.520;苏氨酸在42,49(2个),55,80,105(2个)位点出现磷酸化位点,得分分别为0.573,0.817,0.779,0.662,0.508,0.855,0.507;酪氨酸在15位出现活跃位点,得分为0.788,该蛋白的激酶位点有DNAPK、PKC、PKA、CKI、CKII,共5种(表4)。
NetNGlyc预测三元杂交育肥猪N端糖基化位点(图12-B),预测值(0.554 6)大于0.5,预测结果可靠,氨基酸序列中,在70位点存在1个潜在的N端糖基化位点。

图12 激酶磷酸化位点(A)和N-端糖基化位点预测(B)Fig.12 Kinase phosphorylation site(A)and N-terminal glycosylation site prediction(B)

表4 三元杂交育肥猪FABP2蛋白可能的磷酸化位点Tab.4 Possible phosphorylation sites of FABP2 protein in DLY fattening pigs
2.9 三元杂交育肥猪FABP2蛋白的保守结构域和折叠无序化分析
NCBI中预测到2~131位点含有一个Lipocalin-FABP2超家族保守结构域,是脂质运载蛋白(图13)。运用FoldIndex进行固有无序化分析,存在3个无序化区域,分别在1~35,90~102,105~132区段,所含氨基酸数分别为35,13,28个,未折叠氨基酸总数76个(图14-A)。与亲缘关系最远的鸡比较,鸡在1~35区域存在含35个氨基酸的1个无序化区域,存在很大差异(图14-B)。与亲缘关系最近的绵羊比较,绵羊存在1个无序化区域,无序化区域与鸡相同,表明不同物种的蛋白质发挥的功能有所不同(图14-C)。
3 结论与讨论
FABPs家族成员现已发现至少9种,该家族成员的基因结构基本相同[21],均属于低分子量的细胞内脂质伴侣,在各组织中广泛表达,主要参与脂肪酸转运和调控细胞增殖[22]。FABP2蛋白是哺乳动物细胞内的一种可溶性蛋白质[22],可特异性结合长链脂肪酸,吸收膳食脂肪酸[23-24],参与长链脂肪酸的细胞内运输[25]。此生物学特性对改善肉品质具有潜在价值,可作为体内脂肪沉积的候选基因,在分子育种中前景广阔。已有学者克隆了斑马鱼[26]和标枪虾虎鱼[22]的FABP2基因,发现该基因编码132个氨基酸。Larkina等[27]认为,FABP2基因可作为鸡腹部脂肪沉积的候选基因,但是有关FABP2基因在三元杂交育肥猪上的研究甚少。
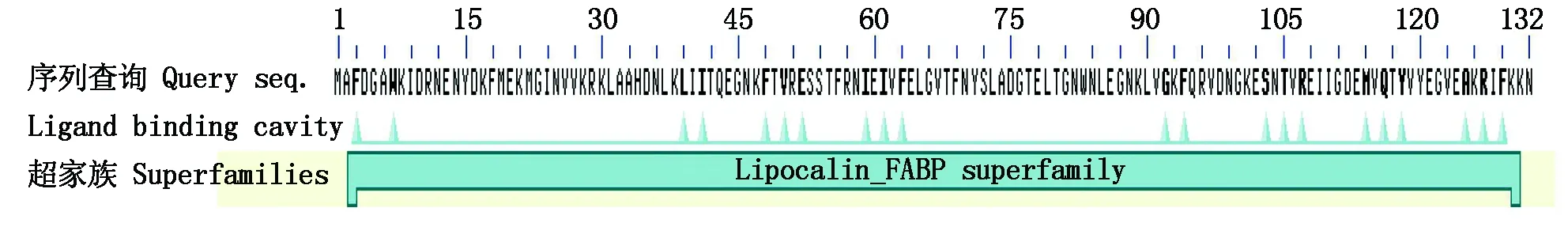
图13 三元杂交育肥猪FABP2蛋白的保守结构域Fig.13 Conserved domain of FABP2 protein in DLY fattening pigs

A.三元杂交育肥猪;B.鸡;C.绵羊。A.DLY fattening pigs;B.Chicken;C.Sheep.
本试验成功克隆了三元杂交育肥猪FABP2基因编码区,全长399 bp,编码132个氨基酸。通过组织表达谱分析发现,FABP2基因在各组织中广泛表达,在空肠中相对表达量最高,但背肌中相对表达量最低。饲粮中添加不同比例花椒籽饲喂三元杂交育肥猪后,试验Ⅰ组和试验Ⅲ组背肌组织中FABP2显著上调表达。与对照组相比,空肠中FABP2在各试验组均显著下调表达。当添加量为7.5%,三元杂交育肥猪背肌中FABP2表达量显著高于对照组。该研究结果说明,饲粮中添加花椒籽显著影响三元杂交育肥猪不同组织中FABP2基因表达水平。Gajda等[28]研究发现,FABP2在哺乳动物空肠中表达量最高,这与本研究结果相似。通过三元杂交育肥猪FABP2氨基酸序列与猪氨基酸参考序列的比对发现,三元杂交育肥猪FABP2氨基酸序列存在2处错义突变,即152位点处的碱基A突变为G,导致第51位点的赖氨酸突变为精氨酸;核苷酸序列308位点的碱基T突变为C,导致第103位点的亮氨酸突变为丝氨酸,初步推测这可能是影响猪脂肪沉积的潜在原因之一,但FABP2基因参与脂质代谢的机制尚不明确,有待进一步研究。
同源性分析表明,三元杂交育肥猪与绵羊亲缘关系最近,核苷酸序列同源性为87.72%,种间相似性高,说明FABP2蛋白在物种间具有高度保守性。蛋白质磷酸化对蛋白质发挥功能具有极其重要的作用,可通过共价修饰的方式修饰1/3以上的细胞蛋白,蛋白激酶仅对丝氨酸、苏氨酸和酪氨酸3种氨基酸残基的磷酸化表现出严格的特异性[29]。本试验预测到三元杂交育肥猪FABP2蛋白存在5个丝氨酸位点、7个苏氨酸位点和1个酪氨酸位点,预测结果合理。此外,还预测到一个N端糖基化位点,蛋白质糖基化大多在膜结合蛋白中分布,这也是影响该蛋白稳定性和功能发挥的原因[30]。FABP2蛋白二级结构主要由延伸链和无规卷曲构成,不存在跨膜螺旋结构,是一种亲水性非跨膜蛋白,主要在细胞质、细胞核、线粒体和过氧化物酶体中发挥作用。当信号肽位点S均值大于0.5时,才具有潜在的信号肽[31],三元杂交育肥猪FABP2蛋白信号肽S预测均值为0.104,表明该蛋白不具有信号肽,是非分泌蛋白,主要在胞质中。蛋白质固有无序化在生物体中具有极其重要的意义,是评价蛋白质功能的重要指标之一,与蛋白质生物学特性密切相关[32]。本试验预测到三元杂交育肥猪存在3个无序化区域,无序化程度高。陈旖婷[33]研究发现,由脂肪细胞分泌的Lipocalin与脂肪沉积有关,三元杂交育肥猪FABP2蛋白在2~131位点含有一个Lipocalin-FABP2超家族保守结构域(脂质运载蛋白),具有配体结合腔,该研究与本研究结果相似。
本试验结果表明,不同比例花椒籽替代部分玉米能显著影响三元杂交育肥猪各组织中FABP2的表达水平,尤其对背肌和空肠组织中的表达影响最为显著。三元杂交育肥猪FABP2基因CDS序列全长399 bp,共编码132个氨基酸,其CDS序列存在2处错义突变,三元杂交育肥猪FABP2基因与绵羊和牛物种同源性较高。三元杂交育肥猪FABP2蛋白为酸性稳定的非分泌蛋白,该蛋白主要以延伸链和无规则卷曲构成,该蛋白无序化程度高,主要在细胞质中发挥作用。本试验结果可为进一步研究FABP2蛋白的生物学特性提供科学依据。
