白细胞介素33在肝细胞癌患者中的表达及其对CD8+T淋巴细胞功能的调节作用
2022-02-11王海鹏李东辉沈光辉
王海鹏, 刘 屹, 李东辉, 沈光辉
1 陕西省人民医院 肿瘤内科, 西安 710068;2 陕西中医药大学附属医院肿瘤医院 三病区, 陕西 咸阳 712000
肝细胞癌(HCC)可诱导机体CD8+T淋巴细胞功能麻痹甚至功能衰竭,不能有效发挥抗肿瘤活性,造成患者疾病进展[1-2],但HCC患者CD8+T淋巴细胞功能衰竭的机制尚未完全阐明。白细胞介素-33(IL-33)是IL-1细胞因子家族成员,主要通过其受体肿瘤抑制素2(suppression of tumorigenicity 2, ST2)发挥生物学功能[3]。IL-33可通过增强效应性CD4+和CD8+T淋巴细胞功能、抑制调节性T淋巴细胞活性在抗肿瘤免疫应答中发挥作用[4-5],但有关IL-33在HCC患者中的表达变化及其对CD8+T淋巴细胞功能调控作用的研究报道较少。因此,本研究检测HCC患者外周血IL-33水平,利用体外细胞培养系统观察IL-33对HCC患者CD8+T淋巴细胞活性的影响和可能的机制,初步探讨IL-33在HCC发病中的作用。
1 资料与方法
1.1 研究对象 选择2019年4月—2020年1 月陕西省人民医院肿瘤内科收住的HCC患者44例作为研究对象。纳入标准:(1)年龄≥18岁且<70岁;(2)诊断符合《原发性肝癌诊疗规范(2019年版)》[6]的标准;(3)未接受手术、介入、免疫、靶向等治疗。排除标准:(1)合并其他恶性肿瘤;(2)合并人获得性免疫缺陷病毒感染;(3)合并自身免疫性疾病或长期使用免疫抑制剂;(4)妊娠期或哺乳期妇女。选择同时期在本院进行查体的健康者20例作为对照组。
1.2 仪器与试剂 美国Sigma 3-16K离心机;德国美天旎公司MACS LS分离柱和MACS磁力分离架;美国伯乐iMark微孔读板仪;美国Applied Biosystem公司ABI7500实时定量PCR仪;德国AID公司酶联斑点吸附试验(enzyme linked immunospot assay, ELISPOT)读板仪;美国BD公司FACS Aria II流式细胞仪。人外周血淋巴细胞分离液(密度:1.077 g/ml)(货号:P8610)购自北京索莱宝科技有限公司;人CD8+细胞分选试剂盒(货号:130-096-495)购自德国美天旎公司;重组人IL-33(货号:200-33)购自美国Peprotech公司;人IL-33(货号:CSB-E13000h)、人ST2(CSB-E13789h)、人IFNγ、人TNFα酶联免疫吸附试验(ELISA)试剂盒购自武汉华美生物公司;Trizol试剂(货号:15596018)购自美国Invitrogen公司;PrimeScript预混反转录试剂盒(货号:RR036A)和TB Green预混Ex Taq Ⅱ (Tli RNaseH Plus)实时定量PCR试剂盒(货号:RR82LR)购自宝日医生物技术(北京)有限公司;细胞计数试剂盒8(cell counting kit-8, CCK-8)(货号:C0037)和乳酸脱氢酶(lactate dehydrogenase, LDH)细胞毒性检测试剂盒(货号:C0016)购自武汉碧云天生物技术公司;人穿孔素ELISPOT试剂盒(货号:ab62943)和人颗粒酶B ELISPOT试剂盒(货号:ab62922)购自美国Abcam公司;小鼠抗人CD3-PE CF594(货号:562280)、小鼠抗人CD8-APC Cy7(货号:348793)、小鼠抗人程序性死亡受体-1(programmed death-1, PD-1)-FITC(货号:557860)、小鼠抗人细胞毒性T淋巴细胞相关蛋白4(cytotoxic T-lymphocyte-associated protein 4, CTLA-4)-PE(货号:555853)购自美国BD 公司;小鼠抗人淋巴细胞活化基因-3(lymphocyte activation gene-3, LAG-3)-PerCP-eFluor710(货号:46-2231-82)购自美国ThermoFisher公司。
1.3 方法
1.3.1 血浆和外周血单个核细胞(peripheral blood mononuclear cells, PBMC)的分离 HCC患者和对照者于清晨空腹采集EDTA抗凝外周血20 ml。1000 r/min离心10 min,取上层血浆冻存于-80 ℃备用。使用人外周血淋巴细胞分离液,采用密度梯度离心法分离PBMC,冻存于液氮中备用。
1.3.2 CD8+T淋巴细胞分选 使用德国美天旎公司人CD8+细胞分选试剂盒对28例HCC患者和11例健康对照者PBMC中的CD8+T淋巴细胞进行纯化。取107个PBMC,4 ℃、300 r/min离心10 min,弃上清后保留细胞沉淀,加入40 μL缓冲液重悬细胞,加入10 μL生物素标记的人CD8+细胞抗体鸡尾酒(其中包含生物素标记的抗CD4、抗CD15、抗CD16、抗CD19、抗CD34、抗CD36、抗CD56、抗CD123、抗TCRγ/δ、抗CD235a),4 ℃孵育5 min,加入30 μL缓冲液,并加入20 μL人CD8+细胞MicroBead鸡尾酒(其中包含MicroBead标记的抗CD14、抗CD61、抗生物素),4 ℃孵育10 min,加入缓冲液调整总体积至500 μL。将MACS LS分离柱置于MACS磁力分离架的分离区,加入3 mL缓冲液充分浸润,然后将上述细胞悬液加入LS分离柱中,在重力作用下使细胞悬液穿过分离柱,收集穿过分离柱的未标记细胞即为纯化的CD8+T淋巴细胞。
1.3.3 细胞培养 纯化的CD8+T淋巴细胞使用重组人IL-33(5 ng/mL)刺激培养,同时加入抗CD3/抗CD28维持T淋巴细胞活性,培养24 h后收集细胞。选择HLA-A02限制性HCC患者(11例)和对照者(6例),取104个IL-33刺激后的CD8+T淋巴细胞与105个HLA-A02限制性的HCC细胞系HepG2细胞[7]共培养,向培养液中加入抗CD3/抗CD28维持T淋巴细胞活性,培养48 h后收集上清。
1.3.4 ELISA法检测血浆和培养上清中细胞因子 使用商品化的ELISA试剂盒对血浆中IL-33、ST2和培养上清中IFNγ、TNFα水平进行检测。向抗体包被的ELISA平板中加入100 μL标准品或待测样本,37 ℃孵育2 h,弃去液体,不洗涤,加入100 μL生物素标记抗体工作液,37 ℃孵育1 h,弃去液体,洗涤5次,加入100 μL辣根过氧化物酶标记的酶结合工作液,37 ℃孵育30 min,弃去液体,洗涤5次,加入90 μL TMB溶液,37 ℃避光孵育15 min,加入50 μL终止液,在450 nm波长处测量吸光度,绘制标准曲线,计算待测样本中相关因子的水平。
1.3.5 实时定量PCR法检测PBMC中IL-33和ST2 mRNA相对表达量 使用Trizol试剂对PBMC中的总RNA进行提取。取1 μg总RNA,使用PrimeScript预混反转录试剂盒进行反转录反应,反转录体系:4 μL 5×PrimeScript反转录缓冲液、1 μL PrimeScript反转录酶混合物Ⅰ、1 μL 寡聚核苷酸引物(50 μmol/L)、1 μL 随机六引物(100 μmol/L)、1 μg总RNA,加入DEPC 处理H2O调整总体积至20 μL,反转录反应条件:37 ℃ 15 min、85 ℃ 5 s。使用TB Green预混Ex Taq Ⅱ(Tli RNaseH Plus)实时定量PCR试剂盒进行实时定量PCR反应,实时定量PCR体系:10 μL TB Green预混Ex Taq Ⅱ(Tli RNaseH Plus) (2×)、0.8 μL PCR上游引物(10 μmol/L)、0.8 μl PCR下游引物(10 μmol/L)、0.4 μL ROX参考染料Ⅱ(50×)、2 μL cDNA模板、6 μL灭菌水,总体积为20 μL,实时定量PCR反应条件:95 ℃ 30 s 1个循环,95 ℃ 5 s、60 ℃ 30 s 40个循环。使用ABI7500实时定量PCR仪进行扩增,采用2-ΔΔCT法以内参照基因为对照进行半定量分析。PCR引物序列参照既往发表文献[8]合成。
1.3.6 CCK-8法检测细胞增殖 使用CCK-8试剂盒对细胞增殖进行分析。在培养的最后4 h,向培养液中加入10% CCK-8工作液,培养结束后立即在450 nm波长处测定吸光度,根据已知细胞数量的OD450nm值绘制标准曲线,计算待测样本细胞数量。
1.3.7 ELISPOT法检测CD8+T淋巴细胞分泌穿孔素和颗粒酶B水平 将无IL-33刺激和经IL-33刺激的CD8+T淋巴细胞加入预包被的ELISPOT平板中,加入植物血凝素(100 ng/mL),37 ℃、5% CO2条件下培养12 h,弃去液体,洗涤3次,加入100 μL链霉亲和素碱性磷酸酶耦合物,室温孵育60 min,弃去液体,洗涤3次,加入100 μL BCIP/NBT碱性磷酸酶显色试剂,室温孵育15 min,终止反应,洗涤后使用ELISPOT读板仪进行细胞计数,使用斑点形成细胞数量表示阳性细胞数量。
1.3.8 流式细胞术检测CD3+CD8+T淋巴细胞比例和CD8+T淋巴细胞中PD-1、LAG-3和CTLA-4比例 PBMC转入FACS管中,洗涤后加入小鼠抗人CD3-PE CF594、小鼠抗人CD8-APC Cy7,室温避光孵育30 min,洗涤后加入2%多聚甲醛/PBS溶液固定。将无IL-33刺激和经IL-33刺激的CD8+T淋巴细胞转入FACS管中,洗涤后加入小鼠抗人CD8-APC Cy7,室温避光孵育30 min,洗涤后加入100 μL固定液,室温避光孵育30 min后, 加入2 mL透化液,室温400 r/min离心5 min,弃上清,加入100 μL透化液,然后加入小鼠抗人PD-1-FITC、小鼠抗人LAG-3- PerCP-eFluor710、小鼠抗人CTLA-4-PE,室温避光孵育30 min,加入2%多聚甲醛/PBS溶液固定,使用FACS Aria Ⅱ流式细胞仪获取细胞,使用FlowJo V10软件分析结果。
1.3.9 靶细胞死亡比例检测 使用LDH细胞毒性检测试剂盒对CD8+T淋巴细胞诱导的靶细胞死亡比例进行检测。在培养的最后1 h,将细胞培养板于400 r/min离心5 min,加入150 μL LDH释放试剂,混匀后继续培养1 h,培养结束后吸取120 μL上清液,加入新的96孔板中,各孔中加入60 μL LDH检测工作液,室温避光孵育30 min,然后在490 nm处测定吸光度。以HepG2细胞培养上清中的LDH吸光度为“对照值”,以Triton X-100处理的HepG2细胞培养上清中的LDH吸光度为“最大活性值”,靶细胞死亡比例(%)=(待测样本值-对照值)/(最大活性值-对照值)×100%。
1.4 伦理学审查 本研究方案经陕西省人民医院医学伦理委员会,批号:省医伦2017-082号,所有患者或家属行告知并签署知情同意书。
2 结果
2.1 一般资料 HCC组44例,均有HBV感染史,对照组20例,两组在性别比例、年龄方面的差异均无统计学意义(P值均>0.05);但HCC组AFP显著高于对照组(P<0.001)(表1)。
2.2 HCC患者外周血CD8+T淋巴细胞比例、IL-33和ST2水平变化 HCC组和对照组外周血CD3+CD8+T淋巴细胞典型流式分析图见图1。HCC组血浆IL-33水平显著低于对照组,HCC组PBMC中IL-33 mRNA相对表达量亦显著低于对照组(P值均<0.001)(表2)。外周血CD3+CD8+T淋巴细胞比例与血浆IL-33、血浆ST2水平无显著相关性(P值均>0.05)(图2)。
2.3 IL-33对HCC患者CD8+T淋巴细胞功能的调节作用和机制 105个纯化的CD8+T淋巴细胞(28例HCC患者和11例对照者)使用重组人IL-33刺激24 h后,无论HCC组还是对照组纯化的CD8+T淋巴细胞的增殖水平在无IL-33刺激和经IL-33刺激之间的差异均无统计学意义(P值均>0.05)。在无IL-33刺激和经IL-33刺激的情况下,HCC组CD8+T淋巴细胞分泌穿孔素和颗粒酶B的水平显著低于对照组;经IL-33刺激后两组CD8+T淋巴细胞分泌孔素和颗粒酶B的水平均显著升高(P值均<0.05)。HCC组和对照组CD8+T淋巴细胞无IL-33刺激和经IL-33刺激后PD-1、LAG-3和CTLA-4表达的典型流式分析图见图3。在无IL-33刺激的情况下,HCC组CD8+T淋巴细胞中免疫检查点分子(PD-1、LAG-3、CTLA-4)阳性细胞比例显著高于对照组(P值均<0.05),但经IL-33刺激后两组CD8+T淋巴细胞中PD-1、LAG-3和CTLA-4阳性细胞的比例与无IL-33刺激比较无统计学差异(P值均>0.05)。经IL-33刺激后HCC组CD8+T淋巴细胞中PD-1、LAG-3、CTLA-4阳性细胞比例亦显著高于对照组(P值均<0.05)(表3)。
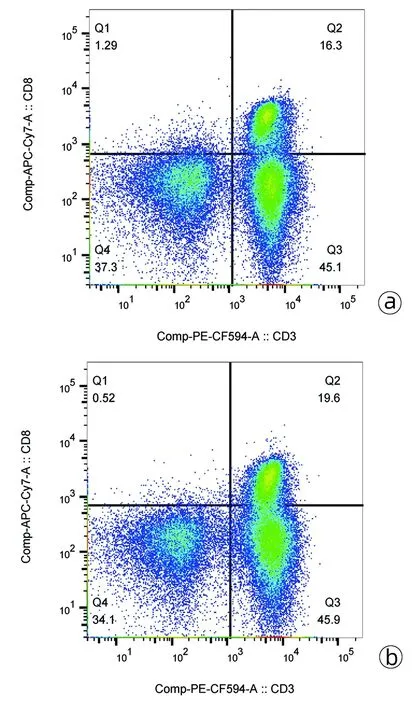
注:a,HCC组; b,对照组。图1 CD3+CD8+T淋巴细胞典型流式分析图

图2 CD3+CD8+T淋巴细胞与IL-33、ST2水平相关性分析

表1 两组一般资料比较
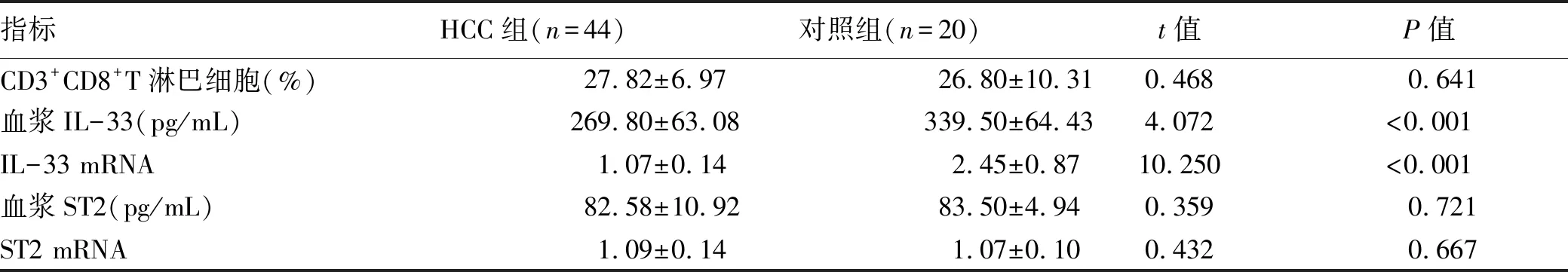
表2 两组IL-33和ST2水平比较
选择11例HLA-A02限制性HCC患者和6例HLA-A02限制性对照者的CD8+T淋巴细胞,IL-33刺激培养后与HepG2细胞共培养。在无IL-33刺激的情况下,HCC组CD8+T淋巴细胞诱导靶细胞死亡比例、IFNγ和TNFα分泌均低于对照组(P值均<0.05)。重组IL-33刺激后两组CD8+T淋巴细胞诱导靶细胞死亡比例、IFNγ和TNFα分泌均显著升高(P值均<0.05)(表4)。
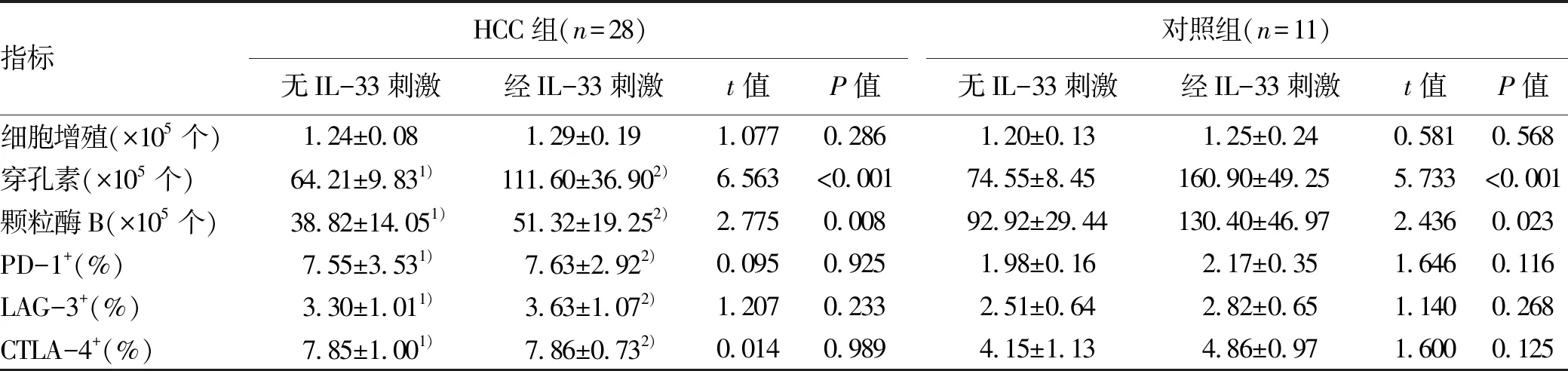
表3 两组无IL-33刺激和经IL-33刺激后CD8+T淋巴细胞增殖、毒性分子分泌和免疫检查点分子表达的比较

图3 两组CD8+T淋巴细胞无IL-33刺激和经IL-33刺激后PD-1、LAG-3和CTLA-4表达的典型流式分析图
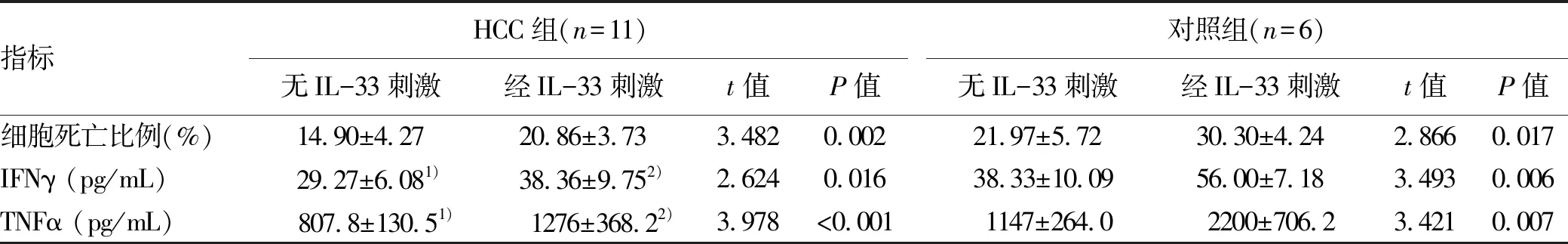
表4 无IL-33刺激和经IL-33刺激后CD8+T淋巴细胞杀伤功能比较
3 讨论
在组织损伤过程中,IL-33可作为一种警戒分子释放至组织和外周血中,通过ST2受体介导的信号通路活化固有免疫和适应性免疫应答中的多种免疫细胞,在多种疾病中发挥多能性效应[9]。但IL-33和ST2在肝脏疾病中的表达水平和功能在不同研究中却存在差异性结果。慢性乙型肝炎患者血清IL-33水平显著升高,与HBV DNA载量呈负相关,与ALT水平呈正相关,可能抑制HBV复制[10]。肝脏浸润免疫细胞中的IL-33/ST2介导的信号通路可增加药物性肝损伤诱导的肝脏炎症[11],通过增强中性粒细胞胞外诱捕网形成促进肝脏炎症损伤[12],促进HBV相关慢加急性肝衰竭患者单核细胞的炎症风暴效应[13]。但新近的研究[14]发现,IL-33可缓解Poly I∶C和刀豆素A诱导急性肝炎所致的肝脏炎症损伤。本研究结果发现,HCC患者血浆IL-33和PBMC中IL-33 mRNA水平均显著降低,这与在HCC组织中IL-33的表达较癌旁组织中降低的研究结果一致[15]。本研究结果亦发现,无论是可溶型ST2还是PBMC中的ST2 mRNA水平在HCC患者和健康对照者之间的差异均无统计学意义,提示HCC患者IL-33受体的水平和活性可能未受影响,HCC免疫发病过程中可能通过影响IL-33水平诱导机体免疫麻痹或功能衰竭。
在病理学状态下,IL-33可通过抑制Toll样受体介导的固有免疫信号通路[16]、抑制IL-17受体信号通路介导的炎症应答[17]、增加IFNγ分泌[18]、促进中性粒细胞向感染部位的募集浸润[19]等多种机制改善疾病严重程度。但也有研究[20]发现,IL-33可通过促进调节性T淋巴细胞亚群的增殖和活化,促进疾病诱导的长期免疫抑制。但有关IL-33对HCC患者CD8+T淋巴细胞功能的调节作用罕见相关报道。在HCC和结肠癌等肿瘤发生发展过程中,IL-33可在组织局部发挥作用,通过增强效应性CD8+T淋巴细胞向肿瘤组织浸润,增强机体抗肿瘤免疫功能[4,21]。而在营养缺乏和鼠嗜血细胞综合征中,IL-33可通过诱导IFNγ分泌调控CD8+T淋巴细胞的效应功能[22-23]。本研究发现,HCC患者CD8+T淋巴细胞的杀伤功能显著降低,分泌穿孔素、颗粒酶B等毒性分子的水平下降,而多种免疫检查点分子表达水平升高,说明HCC患者中存在CD8+T淋巴细胞功能衰竭。重组IL-33可显著增强HCC患者CD8+T淋巴细胞的毒性作用,但IL-33并不影响CD8+T淋巴细胞的增殖以及免疫检查点分子的表达,但穿孔素、颗粒酶B的分泌显著升高,提示IL-33主要通过促进穿孔素-颗粒酶B途径增强HCC患者CD8+T淋巴细胞活性。但由于新近的研究[24-25]发现IL-33可通过ST2信号通过活化JNK通路诱导肿瘤微环境重塑,促进HCC肿瘤生长。因此,IL-33在HCC发病中的作用仍需要进一步体内实验证实。同时,虽然IL-33可促进HCC组和对照组CD8+T淋巴细胞分泌穿孔素和颗粒酶B。但IL-33是否直接参与穿孔素和颗粒酶B的表达调控仍不清楚,后续研究中将对IL-33对穿孔素、颗粒酶B表达启动子区域的活性影响进行分析,评估IL-33是否直接调控者两种分子的表达。
总之,HCC患者外周血IL-33水平降低。外源性IL-33可通过促进穿孔素-颗粒酶B信号通路增强HCC患者CD8+T淋巴细胞杀伤活性,IL-33可能成为HCC治疗新的靶点。
利益冲突声明:本研究不存在研究者、伦理委员会成员、受试者监护人以及与公开研究成果有关的利益冲突。
作者贡献声明:王海鹏、李东辉参与研究数据的分析解释,并起草文章;刘屹参与实验设计及修改文章关键内容;沈光辉对研究的思路或设计有关键贡献,并参与修改文章。
