提高小麦光能利用效率机理的研究进展*
2022-02-10李义博陶福禄
李义博,陶福禄
提高小麦光能利用效率机理的研究进展*
李义博,陶福禄**
(中国科学院地理科学与资源研究所/中国科学院陆地表层格局与模拟重点实验室/中国科学院大学,北京 100049)
农作物产量依赖于生物量和收获指数,过去作物产量的增加主要得益于收获指数的增加而生物量增加很小,进一步提升作物产量在很大程度上依赖于生物量的提高,而光能利用效率是提高作物生物量的瓶颈。小麦(L.)是一种在全世界广泛种植的谷类作物,可为全世界人口提供约20.0%的能量,阐明小麦光能利用效率变化的内在机理与外在因素,对提高作物资源利用效率和生产力有重要意义。本文对光能利用效率的定义、主要过程、小麦的光能利用特性和内外影响因素进行综述,表明提高光能利用效率具有较大的潜力;论述了外部因素,即光、水分、养分和耕作制度等对小麦光能利用效率的影响,主要表现为在单株尺度上主要由光合作用等内部因素控制,而在田间尺度则由温度、降水、耕作栽培方式等非生物因素控制。进一步分析了目前存在的问题以及在气候变化背景下小麦的适应机制,旨在为提升小麦光能利用效率提供理论依据。未来光能利用效率的研究可利用高通量表型观测技术,与分子标记结合,设计在目标环境下的具有高光能利用效率的理想株型,为培育高产高效小麦品种提供科学依据。
小麦;气候变化;光能利用效率;理想株型;产量潜力
在人口和粮食需求增加、作物产量提升缓慢甚至发生停滞的背景下,如何进一步提高资源利用效率,提升粮食产量,实现农业高产高效,既是国家重大需求问题也是科学前沿问题。小麦是世界三大作物(小麦、玉米和水稻)之一,作为重要的粮食作物,在国家食品安全与经济发展中起到十分重要的作用[1]。2050年之前,在全球尺度上小麦的需求量每年增加1.7%左右,而产量每年仅增加1.1%[2],据联合国粮农组织中长期预测[3],到2015、2020和2030年,全球小麦总需求量分别为7.5×108t、7.8×108t和8.6×108t,分别比2001年总产量(5.8×108t)增加了29.0%、34.3%和48.7%,其中增加量的70.0%来自单产水平的提高。目前每公顷的生产率需要提高50.0%才能满足其需求[4]。相较其它作物,小麦分布广泛,全球种植面积约2.2亿hm2,其产量变化将对全球食品安全产生重大的影响[5]。
小麦作为中国第二大粮食作物,栽培历史悠久,在农业生产及国民经济中占有重要地位。近年来,中国小麦的收获面积占全球小麦总收获面积的10.7%,单产是全球平均的1.5倍,约为4500.0kg·hm−2;总产量在1.1亿t左右,约占世界总产量的16.1%。在此过程中,主要依靠单产的增加从而使小麦增产,其中间套复种、高效施肥、节水灌溉、机械化操作等模式化和科学化的高产栽培技术体系发挥着重要作用[6−7]。同时,选育优良的小麦新品种也是获得小麦高产和稳产的重要途径[8]。面对耕地面积减少和人口增加的双层压力,提高小麦单产是增加总产以及保证国家粮食安全的唯一途径[9−10]。然而,全球气候变暖对各地粮食产量造成不同程度的影响,对中国大部分地区的粮食产量造成不利影响。1980年以来,气候变化致使全球小麦产量降低了5.5%[11]。近几十年来中国平均气温增加1.1°C,同时气候变化引起降水的年际波动增加,极端天气事件出现频率也显著上升[12],对粮食安全的稳定性造成巨大影响[13]。如何在有限的自然资源条件下确保小麦的高效生产已成为国内外关注的焦点。
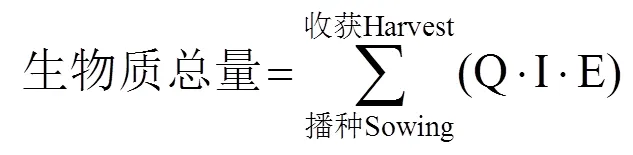
作物产量形成依赖于光合作用。提高作物生物量和收获指数是保证粮食安全和高效生产的重要保障,而提高作物的光能利用效率是增加作物生物产量的重要前提。光能利用效率指在一定时间内,单位土地面积上植物/作物通过光合作用固定的能量与同时间内接受到太阳辐射能的比值。作为最重要的生化反应,植物90.0%~95.0%干物质来源于光合作用的产物。产量的形成是作物将太阳能转化成自身的化学能,因此,提高光能利用效率是打破当前作物产量瓶颈、从而获得高产的理想途径[14]。目前,小麦的收获指数和叶面积指数的增长已经达到瓶颈,为了进一步提高产量,需要提高其生物量,这使得提高作物高光能利用效率成为关键[15−16]。理解光能利用效率的主要损耗过程,分析不同因素对光能利用效率的影响机制对提高作物产量潜力至关重要。为深入研究小麦对光能的高效利用,本文综述了国内外相关文献,从光能的利用与损耗、影响光能利用效率的因素和提高小麦光合作用途径三方面切入,分别阐述了小麦高效光能利用效率的响应机理,旨在为提高小麦光能利用效率和产量潜力的生理基础和高效生产提供理论依据。
1 有关小麦光能利用效率概念内涵的剖析
1.1 光能利用效率的概念
光合作用是决定作物生长和发育的重要因素,其中90.0%~95.0%以上的生物量来自光合作用的积累[17−18]。在其它条件不限制的情况下,光合作用的提高能够增加作物产量,即便是小幅的提升,也能较大地增加生物量,进而提高产量[19]。光能利用效率是指一定时间内单位土地面积上植物通过光合作用固定的能量与同期接受到太阳辐射能的比值[20],不仅表示作物将截获的辐射转化为干物质的能力,也表示作物对太阳总辐射的利用效率[21],其计算公式为[22]
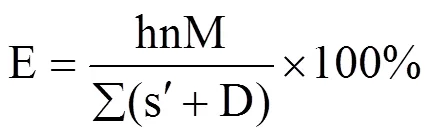
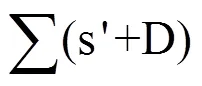
经典作物学认为,作物收获时总干物质量与生长季累积截获太阳辐射量之间呈显著的线性关系,线性方程的斜率即为辐射利用效率,指每单位截获辐射所产生的干物质量(以g·MJ−1计)[23],可利用辐射利用效率获得作物的生物量。
式中,Q是作物生育期内逐日太阳总辐射(MJ);I是作物冠层对光能的截获(%);E指辐射利用效率(g·MJ−1)。根据研究目的的不同,光能利用效率有不同的计算形式,如按时间长短可分为日、月、生长季和年光能利用效率;按不同辐射光光质分为以总辐射、光合有效辐射为基础的光能利用效率;以收获的生物量和以收获的经济产量为基础的光能利用效率[24]。
1.2 光能利用过程
光合作用是小麦光能利用的主要方式。光合作用可大致分为原初反应、同化力形成和碳同化三大阶段。前两个阶段发生在类囊体膜上,最后阶段在类囊体膜外的间质或胞质溶胶中进行[25−26]。原初反应包括光能的吸收、传递以及引起光化学反应,从而产生电荷分离和膜电位[25]。同化力形成是将电荷在膜上一系列定向电子的传递,导致在类囊体内侧H2O被裂解释放出O2,类囊体外侧NADP+等电子受体被还原,在定向电子传递过程中与氢离子相结合并释放,导致膜两侧产生浓度差。氢离子浓度差与原初光反应中产生的膜电位结合,被ATP(腺嘌呤核苷三磷酸)合酶与ADP(腺嘌呤核苷二磷酸)和Pi(磷酸)一同合成ATP。同化力是指形成的NADPH(还原型烟酰胺腺嘌呤二核苷磷酸)和ATP形成的过程,用于碳同化和其它反应。碳同化是由多种酶促反应构成,C3途径和C4途径如图1所示[27]。在C3途径中,CO2和RuBP(1,5-二磷酸核酮糖)在Rubisco(1,5-二磷酸核酮糖羧化/加氧酶)的催化下,产生2分子磷酸甘油酸(PGA),PGA被ATP和NADPH还原成磷酸丙糖,一部分磷酸丙糖用于形成RuBP,以便重新固定CO2;另外,也可在叶绿体内合成淀粉储存能量或进一步合成为蔗糖的形式运输至所需要的部位;在此过程中还会产生光呼吸现象,即RuBP和O2在Rubisco催化下相结合(C2途径)。但在C4途径中,CO2先在叶肉细胞中与PEP(磷酸烯醇式丙酮酸)结合被PEPCase(磷酸烯醇式丙酮酸羧化酶)催化生成OAA(草酰乙酸),再转变成不同植物所需的C4酸,再运输到维管束鞘细胞释放出CO2供C3途径同化。但C4途径运转需要额外消耗ATP,因此只有在强光下方可顺利进行[16]。
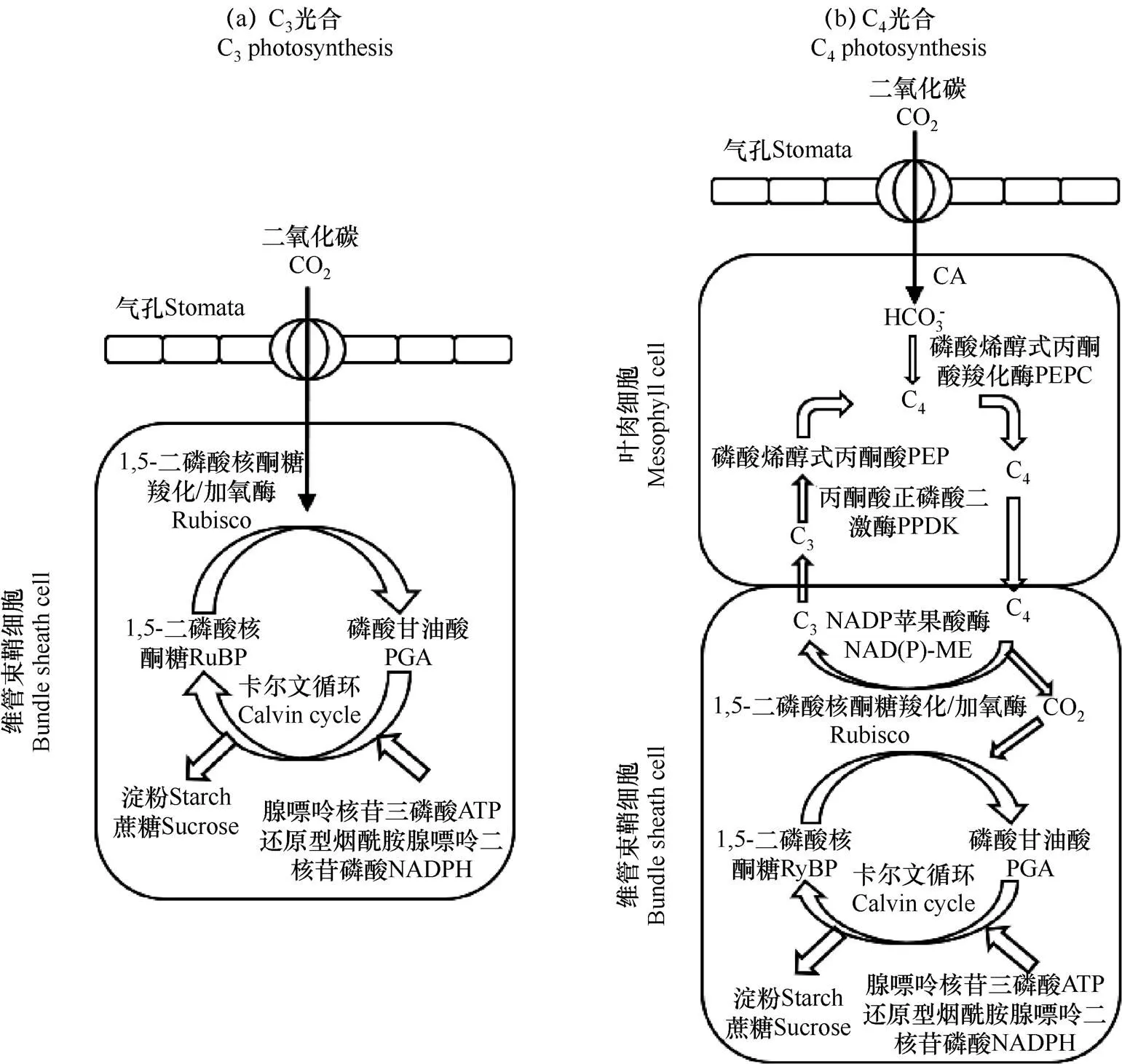
图1 作物C3途径和C4途径光合示意图[27]
1.3 光能损耗过程
对于田间植物,1mol光合有效辐射(PAR)平均含能量约209.0kJ,对于田间植物而言,最大光能利用效率为28.0%[24]。光能的损失发生在植物各个阶段,同化物形成阶段能量的转化效率为32.0%,在碳固定阶段能量转化效率为83.0%,这两个阶段最大光能利用效率为26.6%;另外,田间作物对光合有效辐射的利用率最高为80.0%,因此,光能利用效率最大为9.1%[14]。考虑到呼吸损失,净光能利用效率上限为6.0%。另外,分析照射到叶片表面太阳辐射能在光合过程中的能量损耗可知,通过光合无效辐射、反射和透射、非光合色素吸收、光呼吸和暗呼吸等途径损失,最终光合作用固定的能量约占光能的5.0%(表1)[16,28−29]。C3植物理论的光能转换效率约4.6%,而田间记录的转换效率不足1.6%,表明有很大的空间来提高光合能量转换效率[28,30]。
1.4 小麦利用光能的特性
如何充分将照射到地球表面的太阳辐射用于光合作用,是农业生产中的一个根本性问题[16]。为了提高小麦光能利用效率,主要从3个方面进行研究:(1)提高冠层截获和捕获光的能力,(2)延长光截获利用的时长,(3)提高冠层的光合作用能力和光能利用效率[15]。小麦在传统高投入系统下,针对光的捕获能力冠层结构已被有效地优化[31];就延长光截获利用的时长而言,仍有很多措施能够实现,如通过提高早期叶面积生长速率使得覆盖地表面积增加或引入“持绿”基因从而提高生育期内总光合作用[30−31]。研究表明,与光能捕获、立木建立和延缓叶片衰老等有关的性状,在常规基因库中表现出显著的遗传变异性,且十分适合视觉选择,表明这些因素并不是提高产量潜力的主要瓶颈。一旦冠层结构、光能截获和光合作用时长实现最优化,总光合作用只能通过提升单位叶面积的光合速率来提高[32−33]。
在不同的生育期内小麦的光能利用效率有很大差别,与不同阶段的发育速度和光合产物的积累量有关。在苗期因植株矮小、叶面积指数小、田间低漏光大且温度低,小麦生长缓慢,致使光能利用效率低;返青期−孕穗期,随着温度逐渐升高,群体生长发育加快,光能利用效率随着叶面积增加而增加,干物质积累量加大;但孕穗期后,随着叶面积指数的下降,光能利用效率也呈下降趋势,而在灌浆期会达到第二个高峰(图2a)[34−35]。光合生产率(R,g·m−2·d)是指单位面积内叶面积每日能生产多少干物质量,表示单位叶面积的光合能力,与群体叶面积指数具有显著的相关关系(图2b),随着叶面积指数的增加,单位叶片生产能力不断下降,受到了田间光照强度(I)在冠层内分布(Beer-Lambert定律[36−37])的影响。灌浆期是小麦产量形成的关键时期[38],维持此期间旗叶的光合作用是获得高产的重要基础,但此时叶片的功能下降,不利于籽粒的灌浆[39]。

表1 截获的太阳辐射通过作物冠层光合作用转化为植物碳水化合物过程中的能量损耗[16]
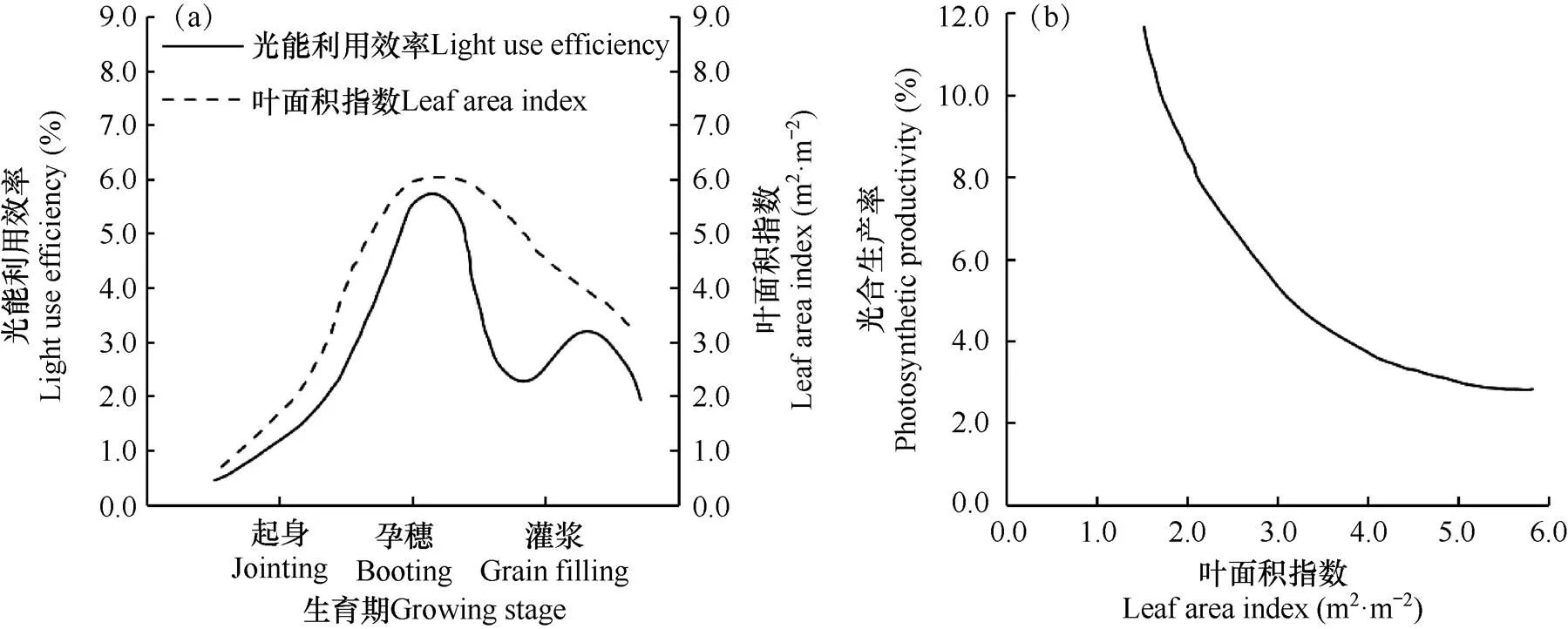
图2 小麦光能利用效率(a)和光合生产率(b)与叶面积指数的关系曲线[34]
1.5 光能利用效率的测定
生物量调查法可直接获得一定时期内植物的光能利用效率。对于草本和经济作物而言,可通过直接收获一定时期内的生物量(干重),再与同时期内所接受到的太阳总辐射相比,即为其光能利用效率[23];木本植物光能利用效率的获得,需要测定对标准地内每株树木的胸高直径,再计算标准木的树干解析,建立生物量或材积与树高、胸径的数学方程,从而计算其生物量,再与同时期内所接受到的太阳总辐射相比,从而计算光能利用效率[40]。生物量调查法最为传统,因所需设备简单,应用最为广泛,但该方法工作量较大,取样具有一定的破坏性[41−42]。
利用光量子效率可获得光能利用效率的理论最大值。光合作用过程分成3个阶段:(1)光能与原料的输送;(2)能量转化:无机物转化为有机物,光能转化为化学能;(3)能量的运输与贮藏,合成的碳水化合物转运或贮藏到其它器官用于生长发育[43];根据理论值计算,每还原1molCO2所生成的自由能约为478.8kJ,需要提供8mol光量子的能量1823.8kJ,光合作用的最大效率为26.3%[44]。中国自然植被与主要作物的光能利用效率分别为0.2%和0.3%,在生长旺盛阶段一些植物的光能利用效率可达7.0%。光量子效率法能够避免对植被的破坏且精度大,但易受光质和光强的影响,同时也会受到光合碳代谢途径的影响,如C3植物量子效率会比C4高30.0%左右[45]。
涡度相关技术作为目前唯一直接观测植被冠层CO2、水热交换的方法,为研究生态系统尺度内光合作用相关参数提供了依据,已被广泛用于陆地生态系统碳通量的研究中。Wang等[46]利用CO2通量观测数据与光合有效辐射计算的半干旱地区玉米的εmax为1.2gC·MJ−1,退化草原的εmax为0.8gC·MJ−1。李正泉[47]利用CO2通量数据估算高寒草甸的εmax为0.9gC·MJ−1。利用涡度相关技术估算光能利用效率,在时间尺度上具有灵敏度高的特点,能够反映日尺度上的变化;在空间尺度上,可从冠层水平延伸到景观水平,甚至能够与卫星遥感的尺度转换有关联[48];但因其设备昂贵,此外对下垫面性质有较高要求,因而限制了其被广泛应用。
利用生产力模型来计算光能利用效率也是一种方法。刘真真等[45]利用CASA模型分析不同区域冬小麦的光能利用效率发现,不同季节内光能利用效率有所差异,3、4和5月的光能利用效率先升高后降低,最高约为1.8g·MJ−1。付刚等[49]利用LUE模型对不同海拔的高寒草甸光能利用效率的研究发现,随着海拔的增加,光能利用效率先增加后降低,最高维持在0.6gC·MJ−1以下。
2 有关内外因素影响小麦光能利用效率机理的分析
2.1 作物自身因素
2.1.1 植株冠层截光
株型是指作物的外在形态、分布形式以及与光能利用效率有关的生理、生态机能的一系列性状[50]。冠层结构的研究一直是作物生理、育种和栽培等领域的研究热点,不仅直接关系到太阳光能的截获量,而且影响冠层水、热、气等分布,从而影响作物群体的光能利用效率,进而影响作物产量[51−52]。矮化基因(Rht)的引入对谷物产量有较大影响,大大提高了收获指数[53]。胡延吉等[54]分析了山东省40a间品种更替的变化,发现植株的矮化以及茎秆向粗壮型变化,株型结构合理,改善了群体内的透光性,使得品种抗倒伏能力增强,从而提高收获指数。在小麦发育后期,旗叶的功能性对籽粒产量影响较大;姚维传等[55]发现,旗叶的叶长、叶宽和叶面积与穗粒重呈现正相关关系,并发现具有较小的开张角和披垂度的宽叶与产量呈正相关关系。张娟等[56]认为,株高应适中,稳定在80.0~85.0cm为好;Berry等[57]发现,与高产相适应的最小株高为70.0cm,既能保证较高的收获指数,又具有较强的抗倒伏能力;叶片应斜立,自上而下叶基角逐渐增大,茎叶夹角小,有助于增加对光能的截获,提高群体光合速率[50]。
叶片的朝向决定着作物对光能的截获,进一步影响光合特性。一天内光合有效辐射(PAR)因太阳辐射角度的变化而变化,植物通过改变叶片取向和叶绿素排列等方式来躲避强光,进而能减少光抑制现象,维持光合特性[15]。陈悦等[58]研究表明,叶角对光合作用和Rubisco羧化效率具有显著影响,旗叶水平时光合速率和羧化效率显著高于直立叶片,可能由于直立叶片受光叶面积小,光合效率低。曾浙荣等[59]设置不同行距与播种密度发现,中间型的籽粒产量大于披叶,最低为直立型叶(P<0.01),表明叶角分布越合理,有利于光能的捕获从而增加产量。姚碧卿[60]指出,只有在叶面积指数大于5.0时,直立叶才有利于光能利用效率的提升。康祥波等[61]提出了“动态株型”的概念,指植株的株型会随生长变化而变化。魏燮中等[62]认为为满足籽粒对光能的需求,灌浆初期旗叶的叶角要小,随着持续灌浆,叶角要逐渐增大,产生由直立到水平、再到倒垂的动态变化,这种特性还有利于调节灌浆高峰期能量的供需关系。良好的株型不仅能有效截获光能,同时能够在生育期内保证光能的有效转化。Long 等[50]比较两种株型(株型X:叶片平展,株型Y:叶片角度自上而下逐渐增大)发现,株型Y的冠层内光量子通量密度分布合理,净光合速率大,使得光能利用效率较株型X提高39.4%。光能利用效率随着冠层截获量的增加而增加,通过改善冠层内部透光结构以增加对光能的截获,可有效提高小麦的光能利用效率,在小麦株型的透光性方面,要求株型紧凑、叶片直立以提高光能利用效率。因此,选育株型抽穗前紧凑,灌浆后期松散,叶水平角随灌浆进程逐渐加大的品种,以及小麦功能叶在整个生育时期均能充分截取和利用光照,以上冠层结构有助于提高光能利用效率,从而增加小麦产量[63]。
2.1.2 根系吸收
良好的根系形态是作物高效生产的基本保证。根系是小麦吸收水分与养分的器官,其发达程度决定了作物吸水能力。倒伏是小麦生育后期遭遇的一种灾害,严重倒伏会使产量降低80%以上,同时还会降低籽粒的品质[64]。研究表明,根部抗倒伏能力与根扩展的幅度和深度有关,同时也与根系周围土质有关[65]。凌启鸿等[66]研究表明,根系的分布决定着作物叶片角度的大小,垂直分布的根系越多,群体叶面积指数越大,群体内的透光性越好,有利于光合作用的进行。因此,可通过改善根部的形态与分布,既能有效减少倒伏,又能提高作物的光能利用效率[51]。
2.1.3 非叶器官光合
小麦籽粒灌浆的碳同化来源主要有三个途径,即旗叶光合、非叶器官光合以及茎秆的转运[67]。Araus等[68]利用同位素与遮光处理发现,穗部光合同化约59.0%干物质形成小麦籽粒。在旗叶节以上的光合器官对籽粒重的贡献率中,非叶器官(穗部、旗叶鞘和穗下节间等)对产量的贡献高达70.0%以上[69]。其中,去掉颖护颖、外稃和内稃、颖外稃和内稃分别使籽粒重减少了6.7%、52.5%和88.2%[70],去掉芒后降低了小麦穗部光合速率,从而影响籽粒产量[71]。尽管碳同化的比例会随着品种、环境因素、测量方法的不同而改变,但穗部光合对产量的贡献率大于旗叶,是小麦产量构成的重要光合器官[72]。由于麦类作物非叶器官光合面积占据大量面积,不仅在时空结构方面利用光能更加合理,同时在生理机能方面占据优势:(1)穗部光合具有较高的光补偿点,无明显的光合午休现象,在经历水分胁迫与高温时,穗部能够维持较高的耐受性和水分利用效率;(2)籽粒生长旺盛时,呼吸所排放的CO2能够被穗部器官重新固定,能够维持较高的光合速率;(3)由于非叶器官面积较大,且衰老程度比旗叶慢,光合作用优势明显[67,69]。因此,要充分发挥非叶器官的光合潜力,对于提高小麦群体光合能力和光能利用效率具有重要意义。
2.2 气象等因素
2.2.1 光强影响
小麦作为喜光喜温的作物,其生长、发育和繁殖受气候条件的影响巨大。当外界环境发生变化时,光合作用速率也会随之改变,进而影响产量,尤其是光照条件,充足的光照是小麦正常抽穗、开花和灌浆的必要前提[73]。光照强度的变化对植物的光合传递与光能的分配具有显著的影响[74]。张永强等[75]研究表明,弱光条件能够有效降低灌浆期内冠层温度,但光合产物合成受到抑制,从而降低小麦的灌浆速率。叶片光合作用对光响应曲线及其模拟是研究植物光合作用与环境变化相互作用的重要手段[76]。林植芳等[77]的研究指出,光照强度的提升能够显著提高光反应中心II的总电子流速度,降低了光能向碳同化的分配比率。孙旭生等[78]的研究表明,随着光照强度的增加,小麦旗叶的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)明显增加,胞间CO2浓度(Ci)降低,净光合速率的增加有利于产生较多的干物质量,能够增加小麦产量。李华伟等[79]研究指出,适度的遮光有助于提高产量,但重度遮光则显著降低产量,随着遮光程度的增加,小麦叶片变大、变薄、叶面积指数(LAI)增加,从而提高小麦冠层对光能的截获,一定程度上弥补了光照强度降低的影响。光补偿点(LCP)表征植物在弱光下利用光能的大小,而光饱和点(LSP)是指利用强光的能力[80],小麦冠层上部叶片由于受到光照的直射,光照强度往往超过其光饱和点,而下部叶接收的光照不足,光合较低;通过提高上部叶片光饱和点、降低下部叶片的光补偿点,能够有效地提高小麦群体的光合势。
2.2.2 光质影响
光质即太阳辐射中不同波长的光谱成分,植物不能利用所有波段的太阳辐射,对光合作用有益的辐射称为光合有效辐射(PAR),其波长在380.0~700.0mm。波长较长的波段光能利用效率低,反之光能利用效率高。不同波长所含的能量不同,能够显著影响作物的形态建成与代谢,进而影响营养生长与品质[81]。叶绿素吸收最多的是红橙光,其次为蓝紫光,而绿光吸收量最少。李慧敏等[82]研究表明,在蓝红复合光(BR)较单色光能够增加幼苗的鲜重、干重和光合色素含量。张立伟等[83]研究指出,红光能使植物的株高、鲜重和干重的增加,蓝光能够促进叶绿素含量和可溶性蛋白的增加,红光条件下植物的碳水化合物含量和光合速率较高。利用不同光质组合,能够培养出健壮且籽粒品质高的小麦,是一种简便易行的培育方法。
2.2.3 CO2浓度影响
CO2作为光合作用的原料,大气中CO2浓度升高,有利于作物的光合作用,同时降低气孔导度以改善水分关系,从而提高水分利用效率[84]。在干旱胁迫发生时,大气中CO2浓度升高有助于缓解干旱的负效应,提高作物的抗旱性,保证产量的稳定[85]。大气中CO2浓度的升高不仅是构成气候变化的主要原因,而且作为作物光合作用的必要条件也影响着作物的生长发育[86]。在田间试验中发现[47],CO2浓度升高能够提高作物的生物量和产量;叶片光合作用速率得以提高从而使产量增加,可能是由于CO2浓度升高,降低了气孔导度(Gs)和蒸腾速率(Tr),使得胞间CO2浓度(Ci)与水分利用效率(WUE)分别提高了43.3%和37.3%[87]。韩雪等[88]研究指出,CO2浓度的升高,具有“气肥效应”,能够促进小麦拔节期的株高、单位面积内穗数和穗粒数的增加,从而提高产量。汪东炎等[89]利用开放式CO2浓度升高系统(Free Air CO2Enrichment, FACE)模拟CO2浓度升高对籽粒重的影响,发现CO2浓度升高显著增加籽粒重。CO2饱和点高,施肥效应明显,而C4作物具有CO2浓缩机制,在气孔关闭时也能利用较低的CO2,从而能够维持较高的光合速率,如何将C4途径引入小麦中,是目前研究的重点[90]。
2.2.4 温度影响
近百年来,平均地表温度上升了0.7℃,且近50a来温度以0.1℃·10a−1持续上升[91]。温度过高或过低会影响酶的活性,从而影响光合作用。增温能够使小麦开花前的生育期缩短,延长开花后的生育时长,致使产量增加16.3%[92]。气候变化对自然环境以及人类的生产活动都产生了重要影响,其中对生物物候的影响尤为明显。气候变暖通常会导致作物生长速度加快,生育期缩短,但生物量和产量影响不一,既有增加生物量,又有减少生物量,还有生物量保持不变;吴冰洁等[93]研究表明,在影响华北平原冬小麦产量变异的因素中,温度起到了更为显著的作用,但也受到了品种和管理措施的影响。相关研究指出气温升高尤其是春季平均气温的升高对于冬小麦抽穗期与开花期影响极为显著[94]。以往研究表明,气候变暖可能有利于提高华北平原的小麦产量;增温能够减少霜冻灾害、增加叶面积指数、以及开花期提前降低灌浆期温度,减少高温热害的程度[95−96]。在热带地区,每年可额外获得一次以上的收成,其中一个主要方面是增温缩短生育期以实现快速收成[97]。整体上,小麦物候变化特征为,抽穗开花期和成熟期以提前为主,营养生长期和全生育期缩短,生殖生长期延长[98−99],及时调整农业管理措施,例如种植长生育期品种,提前或推迟播种期等,提高管理水平,延长作物生育期从而提高光能利用效率,可以一定程度上抵消气候变化对作物生育期的不利影响,保证农业生产的安全[100]。
2.3 人为因素
2.3.1 水分影响
作物产量形成中对水分胁迫的响应最为敏感[101]。冬小麦籽粒形成阶段缺水会显著降低群体光合能力,植株冠层衰老速率加快,严重影响小麦产量[102]。过量灌水也会降低光合速率[103],影响籽粒的积累[104]。杨晓亚等[105−106]研究表明,籽粒产量随着灌溉量的增加先升高后降低,适度的水分亏缺有益于作物产量的增加。净光合速率随着干旱的持续不断减小;干旱持续时间越长,影响光合的非气孔限制因素越明显[107]。不同时间灌水对于小麦生长有不同的影响,起身时灌水有利于增加穗数,拔节期灌水可增加穗粒数,挑旗−开花期灌水能够增加粒重,而在灌浆期灌水减少粒重[108]。房全孝等[109]研究指出,通过改善冬小麦生育后期土壤水分条件,可显著提高群体冠层上部的光能截获率与截获量及光能转化效率。另一方面,灌溉通过改变作物的形态从而影响群体内光的分布。Philip等[110]研究发现,灌溉通过改变冠层叶面积垂直分布,从而影响冠层内光的垂直分布,进而造成小麦光能利用效率的差异以及产量的差异。因此,在小麦生长过程中,为保证其稳定高产需进一步优化灌溉方案。
2.3.2 养分影响
氮素作为光合作用和生长发育过程中重要的元素,在叶片中,70.0%~80.0%的氮素集中在叶绿体中用于光合作用[111],因此,养分投入管理能够提高光能的转化效率。施氮能够显著提高小麦叶片的Fv/Fo、最大光化学效率(Fv/Fm)以及实际光化学效率;但在高氮的情况下,实际光化学效率会有所降低[112]。当缺少氮素时,小麦生长发育迟缓,植株矮小,叶片发黄,致使小麦的产量和品质下降。施肥不仅能够提高叶面积指数,而且也能提高单位叶面积的光合速率。开花−成熟期是小麦籽粒产量和品质形成的关键阶段,其中,开花后营养器官中氮素的转运和吸收是蛋白质含量形成的关键[113]。另一方面,施氮能够增强植株对病虫害的抗性,使小麦适应当地的需求(光周期、生长季持续时间和温度),提高广泛农业区的产量[114]。叶片中氮含量随冠层深度而减小,呈现垂直分布特征,与辐射在冠层中垂直变化相匹配,是一种适应光能的方式[115−116]。在施氮的情况下,由于氮素在植物体内运转方便,作物能够在生育期的后期将下部氮素转运至中部叶片和穗部,使下部叶片氮素含量明显下降[117]。小麦灌浆期叶片的光合能力下降,此时追加氮肥能够提高小麦叶片叶绿素的含量,改善光合能力,维持绿叶的功能性,增加光合产物的积累,从而提高小麦产量[118]。
2.3.3 耕作栽培影响
作物冠层光能截获率大小主要取决于叶面积指数[119],适当增加种植密度不仅能够有效增加叶面积指数,提高冠层光能截获率[120],而且能够增加冬小麦地上氮素积累量,提高籽粒的含氮量[121]。另一方面,冠层内充足的光照有利于提高单位叶面积的光合能力,叶面积指数过大会导致群体光合能力下降,冠层底部光合有效辐射与叶面积呈负相关,因此,要处理好密植与光能利用的关系[22]。灌溉与秸秆还田措施能够显著降低冠层反射率和透光率,提高冠层对光能的截获量[122]。陈素英等[123]研究发现,随着冬小麦播期的推迟,叶面积指数和光能截获量均显著降低,但加速了小麦幼苗分化时间,可通过合理安排播种时间,提高复种指数,增加单位土地内叶面积和光合势,从而提高群体的光能利用效率,推迟夏玉米的收获,达到麦玉轮作中两作物均增产的目的;王东兴[124]利用小麦与油葵复种模式,利用当地“两季不足,一季有余”的气候特点,提高复种指数,充分利用土地和光能资源。另外,间作套种的栽培模式能充分发挥作物空间结构特点,利用光能和热量资源,提高农作物产量。有研究指出,间作能够缓解小麦光合作用的“午休”现象,增加光合产物的有效积累[125]。因此,可以通过有效的耕作方式构建合理的小麦群体,从而提高对光能的截获与转化,实现光能利用效率,最终提高冬小麦群体生物量及籽粒产量。
3 有关提高小麦光能利用效率途径的研究
3.1 增加Rubisco含量
提高作物叶片CO2同化率的途径主要集中在核酮糖−1,5−二磷酸羧化酶(Rubisco)上,Rubisco催化反应是将CO2固定成三碳化合物(C3光合作用)。然而,Rubisco与O2的亲和力较CO2大,形成磷酸乙醇酸和CO2会降低光合效率。在当前大气CO2、O2浓度、饱和光的条件下,Rubisco酶量与活性是限制碳固定速率的决定性因素[126]。为了克服这种限制,增加叶片中Rubisco含量是一种有效的措施。原则上,单位叶面积上的光饱和速率可通过提高单位叶面积上光合机制的总量进一步提高,实际上,它也会受叶绿体中可积累的蛋白质量与叶肉细胞中叶绿体的数量限制[127]。植物为了弥补Rubisco酶的低效性,在叶片中积累了大量的Rubisco,已占到C3植物叶片总氮量的15.0%~30.0%[128]。为了提高叶片中的氮素含量,一方面可增加施肥量,但会造成污染和浪费,这使得提高作物的氮素利用效率(NUE)成为关键。如果没有额外的氮素输入,增加Rubisco含量可能会造成植物体内氮素的重新分配,减少其它酶的含量从而产生新的瓶颈,这可能是增加Rubisco含量不能提高光合作用的原因之一[129]。为改善Rubisco的动力学特性,另一种方法是使用转基因手段利用正向遗传筛选来挖掘小麦和相关物种的Rubisco动力学特性的遗传变异[5],平衡好作为N素储量的Rubisco含量与光合功能的关系。
3.2 减少光呼吸
虽然光呼吸具有代谢和保护作用,但是由于CO2浓度加倍会显著提高作物的光合作用速率,致使光呼吸对作物产量形成产生负面影响[130]。另一方面,Rubisco加氧反应主要取决于叶肉细胞的CO2分压,CO2分压增加则加氧反应被抑制[131]。理论模型表明,在有利条件下,消除光呼吸能够显著提高产量。然而,完全阻断Rubisco的光呼吸代谢是无效的,简单地阻断光呼吸而不改变加氧反应会导致中间产物的不利积累[132]。最近,已经采取了一种替代方法,即将部分叶绿体乙醇酸直接转移到甘油酸中,结果导致光呼吸代谢物和生物量增加,在逆境下也具有良好的特性[133]。
3.3 引入C4机制
一些高产的C4作物如玉米()和甘蔗(),具有浓缩CO2的机制,转运到叶肉细胞中的CO2浓度大约是空气中的10倍,从而限制光呼吸进而提高了光合效率。CO2浓缩是通过C4途径实现的,在磷酸烯醇丙酮酸羧化酶(PEPC)的催化下,将CO2连接到磷酸烯醇丙酮酸上,形成草酰乙酸,这种固定CO2的方式称为C4途径,在下一阶段,草酰乙酸被转变成其它的四碳酸(苹果酸和天冬氨酸)后运输到维管束鞘细胞,在维管束鞘细胞中被降解成CO2和丙酮酸,供Rubisco固定[134]。Mitchell等[135]认为这是提高生物产量的唯一途径,能够满足所需提高的产量。这方面的研究最初只是利用编码C4酶的基因,如PEPC、丙酮酸正磷酸二激酶(PPDK)和NADP苹果酸酶(NADP-ME)来增强作物的光合作用[136−137]。尽管某些物种的C4和C3羧化可以在单个细胞中进行,但在大多数植物中,C4路径依赖于kranz解剖结构,这种结构有利于将叶肉细胞中的大气CO2与维束鞘细胞中释放的CO2产生空间分离,避免了叶肉向大气泄漏CO2,并在维束鞘细胞中的Rubisco部位保持适宜的CO2:O2的比值[138]。因此,目前的重点主要是获取kranz解剖结构,而不是开发单细胞系统[139]。
4 提高小麦光能利用效率的建议
4.1 品种培育:高产高效的关键
气温升高、CO2浓度增加,干旱和降水异常等事件,均会对不同主产区的小麦产生较大的影响,因此,未来农业需在气候变化背景下满足不同需求[140]。气候变化对小麦的影响取决于不同的种植区域,总体而言,高纬度地区最初表现有利于小麦生产,而低纬度地区会因气温升高而减产[141],培育抗旱、耐热的小麦品种是不发达国家亟需解决的问题。而针对特定目标区域内如温度变化、水分状况、纬度、种植制度等因素的育种改良是一种高效的育种方法[142]。国际玉米小麦改良中心(CIMMYT)的一项重要内容是在大范围、多环境下,培育出高产和稳产的种质,这些种质能够应对一些地区的极端气候变化及田间动态[143]。向广适、高产的品种中引入抗逆性基因来提高作物遗传改良潜力是应对气候变化的重要手段。Blum[144]研究指出,在一定生物学限制的因素中,将高产的基因型和与产量无关的抗旱基因型相结合,有可能培育出既抗旱又高产的小麦品种。综上,培育适应性广、抗逆性强的品种与可持续的作物和自然资源的有机结合,将为应对气候变化、提高作物生产的稳定性提供保障。
4.2 表型观测:育种和精准农业的加速器
为了提升小麦光能利用效率和产量潜力,不同学科学者在各自领域努力攻关,遗传学家通过分子标记将产量性状分解成单个遗传因子,作物生理生态模型可模拟品种×管理×环境间相互作用,若把作物模型中品种遗传参数与分子标记结合起来,就使基因与环境相互作用预测成为可能[145]。表型作为基因型与环境互作的结果,对其监测是研究作物遗传育种中数量性状与实施精准农业的重要措施。通过搭建多水平尺度的高通量表型监测平台和分析标记辅助育种,包括全基因组选择(WGS),建立与光能利用效率有关的“基因型−表型−环境”有关的数据库,不仅能够深入挖掘种质资源,也有利于实现高效优质生产[146]。
4.3 理想株型:优良性状的有效结合
从重视产量结构到以高产为主的形态、生理、产量构成因素和与环境互作为基础的株型概念,是近代小麦高产育种的一个重要发展,即构建理想株型群体,其外在形态结构合理,既能够更有效利用光能和水分,又能将光合产物最大限度地转化到籽粒中,协调“源”与“库”间关系[147]。另一方面,为寻求产量的突破,不少研究集中在高产与抗逆性相结合的途径来挖掘和利用作物整株的光合潜力与资源,研究品种与环境间相互作用,针对特定环境设计理想株型。在空间形态上最大限度地截获光能,同时具有良好的光合生理特征,协调合理的产量构成,是小麦高光效研究的目的,同时也是小麦高产的重要基础。
5 结论和建议
在有限的自然资源条件下,如何实现农业高产高效和粮食安全,取决于作物生物产量以及收获指数,而提高作物的光能利用效率是增加作物生物量和产量的重要前提[148]。照射到叶片表面的太阳辐射能进入到光合过程中,通过光合无效辐射、反射和透射、非光合色素吸收、光呼吸和暗呼吸等途径损失,最终光合作用固定的能量约占光能的5.0%,而在野外观测中,光能利用效率更低[16,28−29],表明提高光能利用效率具有较大的潜力。植物90.0%以上的干物质积累都来自光合作用[17−18,149]。
本文综述了小麦光能利用效率变化的内在和外在机理。温度、降水和CO2等因素能够直接或间接影响光能利用效率,从而影响小麦产量。良好的株型不仅能够影响对光能截获,而且也是小麦高效生产的重要保障。叶片的朝向决定着作物对光能的截获,进一步影响光合特性。株型也会随着植物生长而发生动态变化[61]。为满足籽粒对光能的需求,灌浆初期旗叶的叶角要小,随着持续灌浆,叶角要逐渐增大,发生由直立到水平、再到倒垂的动态变化,这种特性还有利于调节灌浆高峰期能量的供需关系[62]。培育具有高产潜力的新品种不仅需要了解气候变化中产量潜力的生理基础,同时要准确选择理想的性状及性状组合。在提高小麦光能利用效率的3个方面研究中,有关冠层拦截光能和营养物质分配到种子的研究已取得较大进展,但通过光合作用将吸收的光能转换成糖的效率不高[16,28]。旗叶的光合作用是谷物糖类的主要来源,增加单叶特别是旗叶的光合能力,是实现产量潜力突破的关键。理论上,可通过扩大和优化由叶冠层捕获的光、在光系统II上更快速地释放非光化学猝灭、增加Rubisco酶的羧化能力以及最小化光呼吸、增强光合酶的再生能力、减少碳循环、优化电子传输链、将C3代谢转变为C4代谢,并添加蓝细菌或藻类系统的成分来泵送CO2或分隔Rubisco等途径优化光合作用以提高产量[9]。
灌浆期是小麦产量形成的关键时期[38],维持此期间旗叶的光合作用是获得高产的重要基础,旗叶在灌浆期仍能够维持较高的叶片功能,但此时叶面积指数不断下降,可进一步研究叶片的“持绿”基因,从而提高生育期内尤其是灌浆期内总光合作用[33]。在光能利用效率研究中,不仅要注重株型和旗叶光合的研究,还要充分发挥非叶器官(穗部、旗叶鞘和穗下节间等)和整株的光合能力,对于提高小麦群体光合能力和光能利用效率具有重要意义[67]。农田生态系统作为陆地生态系统的重要组成部分,其光能利用效率直接决定着农田生态系统碳源−碳汇变化。未来光能利用效率的研究可利用高通量表型观测技术,与分子标记结合,设计在目标环境下的具有高光能利用效率的理想株型。
[1] Slafer G A.Genetic basis of yield as viewed from a crop physiologist's perspective[J].Annals of Applied Biology, 2003,142(2):117-128.
[2] Dixon J,Braun H J,Kosina P,et al(Eds.).Wheat facts and futures 2009[M].CIMMYT,2009.
[3] FAO.FAO statistical yearbook[M].Agriculture Organization of the United Nations,2012.
[4] Murchie E H,Pinto M,Horton P.Agriculture and the new challenges for photosynthesis research[J].New Phytologist, 2009,181(3): 532-552.
[5] Reynolds M,Foulkes J,Furbank R,et al.Achieving yield gains in wheat[J].Plant Cell & Environment,2012,35(10): 1799-1823.
[6] Xiao D,Tao F.Contributions of cultivars,management and climate change to winter wheat yield in the North China Plain in the past three decades[J].European Journal of Agronomy,2014,52:112-122.
[7] 胡立勇,丁艳锋.作物栽培学[M].北京:高等教育出版社, 2008:35-36.
Hu L Y,Ding Y F.Crop cultivation[M].Beijing:High Education Press.2008:35-36.(in Chinese)
[8] 杨靖.小麦产量相关性状的遗传与稳定性分析及QTL定位研究[D].雅安:四川农业大学,2017.
Yang J.Analysis of the genetic and yield stability traits in Wheat (L) and QTL mapping[D]. Ya,an:Sichuan Agricultural University,2017.(in Chinese)
[9] Bailey S J,Parker J E,Ainsworth E A,et al.Genetic strategies for improving crop yields[J].Nature,2019,575 (7781):109-118.
[10] 李克南,杨晓光,刘园,等.华北地区冬小麦产量潜力分布特征及其影响因素[J].作物学报,2012,38:1483-1493.
Li K N,Yang X G,Liu Y,et al.Distribution characteristics of winter wheat yield and its influenced factors in North China[J].Acta Agronomica Sinica,2012,38:1483-1493.(in Chinese)
[11] Lobell D B,Schlenker W,Costa-Roberts J.Climate trends and global crop production since 1980[J].Science,2011, 333(6042):616-620.
[12] 郭建平.气候变化对中国农业生产的影响研究进展[J].应用气象学报,2015(1):1-11.
Guo J P.Advances in impacts of climate change on agricultural production in China[J].Journal of Applied Meteorological Science,2015(1):1-11.(in Chinese)
[13] Ali S,Liu Y,Ishaq M,et al.Climate change and its impact on the yield of major food crops: evidence from pakistan[J]. Foods,2018,6(6):39-58.
[14] Wu A,Hammer G L,Doherty A,et al.Quantifying impacts of enhancing photosynthesis on crop yield[J].Nature Plants, 2019,5(4):380-388.
[15] Parry M A J,Reynolds M,Salvucci M E,et al.Raising yield potential of wheat(II):increasing photosynthetic capacity and efficiency[J].Journal of Experimental Botany,2011, 62(2):453-467.
[16] 程建峰,沈允钢.作物高光效之管见[J].作物学报,2010, 36(8):1235-1247.
Cheng J F,Shen Y G.My humble opinions on high photosynthetic efficiency of crop[J].Acta Agronomica Sinica,2010,36(8):1235-1247.(in Chinese)
[17] Yoshida S.Physiological aspects of grain yield[J].Annual Review of Plant Physiology,1972,23(1):437-464.
[18] Beer C,Reichstein M,Tomelleri E,et al.Terrestrial gross carbon dioxide uptake:global distribution and covariation with climate[J].Science,2010,329(5993):834-838.
[19] Ainsworth E A,Long S P.What have we learned from 15 years of free-air CO2enrichment(face)? a meta-analytic review of the responses of photosynthesis,canopy properties and plant production to rising CO2[J].New Phytologist, 2005,165(2):351-372.
[20] Monteith J L.Solar radiation and productivity in tropical ecosystems[J].Journal of Applied Ecology,1972,9(3):747- 766.
[21] 常清,王靖,余卫东,等.河南省小麦-玉米轮作系统光能利用效率时空分布及其变化原因分析[J].中国农业气象, 2016,37(3):316-325.
Chang Q,Wang J,Yu W D,et al.Tempo-spatial characteristics and impact factors of radiation use efficiency of wheat-maize rotation system in Henan province[J]. Chinese Journal of Agrometeorology,2016,37(3):316-325.(in Chinese)
[22] 龚绍先.粮食作物与气象[M].北京:北京农业大学出版社, 1988.
Gong S X.Food crops and meteorology[M].Beijing:Beijing Agricultural University Press,1988.(in Chinese)
[23] Monteith J L.Climate and the efficiency of crop production in Britain[J].Philosophical Transactions of the Royal Society of London Series(B)Biological Sciences,1977,281 (980):277-294.
[24] 赵育民,牛树奎,王军邦,等.植被光能利用效率研究进展[J].生态学杂志,2007,26(9):1471-1477.
Zhao Y M,Niu S K,Wang J B,et al.Light use efficiency of vegetation:a review[J].Chinese Journal of Ecology,2007, 26(9):1471-1477.(in Chinese)
[25] Leegood R C,Sharkey T D,von Caemmerer S.Photosynthesis: physiology and metabolism[J].Springer Science & Business Media,2006.
[26] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002.
Xu D Q.Photosynthetic efficiency[M].Shanghai:Shanghai Scientific & Technical Publishers,2002.(in Chinese)
[27] Dehigaspitiya P,Milham P,Ash G J,et al.Exploring natural variation of photosynthesis in a site-specific manner: evolution,progress and prospects[J].Planta,2019,250(10): 1033-1050.
[28] Zhu X G,Long S P,Ort D R.What is the maximum efficiency with which photosynthesis can convert solar energy into biomass[J].Current Opinion in Biotechnology, 2008,19(2):153-159.
[29] Zhu X G,Long S P,Ort D R.Improving photosynthetic efficiency for greater yield[J].Annual Review of Plant Biology,2010,61(1):235-261.
[30] Reynolds M P.Avenues for genetic modification of radiation use efficiency in wheat[J].Journal of Experimental Botany,2000,51(90001):459-473.
[31] Horton P.Prospects for crop improvement through the genetic manipulation of photosynthesis:morphological and biochemical aspects of light capture[J].Journal of Experimental Botany,2000,51(suppl-1):475-485.
[32] Dohleman F G,Heaton E A,Leakey A D B,et al.Does greater leaf-level photosynthesis explain the larger solar energy conversion efficiency of Miscanthus relative to switchgrass[J].Plant Cell & Environment,2009,32(11): 1525-1537.
[33] Dohleman F G,Long S P.More productive than maize in the Midwest:how does Miscanthus do it[J].Plant Physiology, 2009,150(4):2104-2115.
[34] 徐恒永,赵君实,徐茂臻.高产冬小麦光能利用效率的研究[J].莱阳农学院学报,1994,11(1):6-11.
Xu H Y,Zhao J S,Xu M Z.Studies on utilization efficiency of solar energy of high-yielding winter wheat[J].Journal of Laiyang Agricultural College,1994,11(1):6-11.(in Chinese)
[35] 龚绍先.对小麦光能利用效率变化的初步探讨[J].气象, 1979,5(8):37-38.
Gong S X.A preliminary study on the change of light energy utilization rate of wheat[J].Meteorological Monthly, 1979,5(8):37-38.(in Chinese)
[36] Monsi M,Saeki T.Uber den lichtfaktor in den pflanzengesellschften und seine bedeutung fur die stoffproduktion[J].Japanese Journal of Botany,1953,14: 22-52.
[37] Guerin W,Rouabah M T,Kaiser R.Light interacting with atomic ensembles:collective,cooperative and mesoscopic effects[J].Journal of Modern Optics,2017,64(9):895-907.
[38] 吴诗光,陈龙,殷贵鸿,等.灌浆期干旱对高产小麦某些生理生化特性的影响[J].河南农业科学,2001,9(2):6-7.
Wu S G,Chen L,Yin G H,et al.Effects of drought on some physiological and biochemical characteristics of high yield wheat in filling stage[J].Journal of Henan Agricultural Sciences,2001,9(2):6-7.(in Chinese)
[39] 陈国祥,张荣铣.小麦旗叶光合功能衰退过程中PSⅡ特性的研究[J].中国农业科学,2004,37(1):36-42.
Chen G X,Zhang R X.Study on characteristics of PSⅡ of flag leaves during photosynthetic functional decline process of yangmai5(L.)[J].Scientia Agricultura Sinica,2004,37(1):36-42.(in Chinese)
[40] Monteith J L.Climate and the efficiency of crop production in Britain[J].Philosophical Transactions of the Royal Society of London Series B- Biological Sciences,1977, 281(980):277-294.
[41] 许大全,丁焕根,苏丽英,等.红豆草和苜蓿的光合效率比较研究[J].生态学报,1991,11(1):89-91.
Xu D Q,Ding H G,Su L Y,et al.A comparative study of photosynthetic efficiency in Sainfoin and Alfalfa[J].Acta Ecologica Sinica,1991,11(1):89-91.(in Chinese)
[42] 王得祥,刘淑明,雷瑞德,等.秦岭华山松群落能量环境及光能利用效率研究[J].西北林学院学报,2003,18(4):5-8.
Wang D X,Liu S M,Lei R D,et al.Energy environment and utilization of solar energy by Pinus armandi community in the Qingling Mountains[J].Journal of Northwest Forestry University,2003,18(4):5-8.(in Chinese)
[43] 于沪宁,赵丰收.光热资源和农作物的光热生产潜力:以河北省栾城县为例[J].气象学报,1982,4(3):327-334.
Yu H N,Zhao F S.On the light and thermal resources and the crop potential productivity-taking Luancheng county of Hebei province as an example[J].Acta meteorological Sinica,1982,4(3):327-334.(in Chinese)
[44] 朱志辉,张福春.中国陆地生态系统的植物太阳能利用率[J].生态学报,1985,5(4):343-356.
Zhu Z H,Zhang F C.Solar energy utilization efficiency of the land plants in China[J].Acta Ecologica Sinica.1985, 5(4):343-356.(in Chinese)
[45] 刘真真,张喜旺,陈云生,等.基于CASA模型的区域冬小麦生物量遥感估算[J].农业工程学报,2017,33(4):225-233.
Liu Z Z,Zhang X W,Chen Y S,et al.Remote sensing estimation of biomass in winter wheat based on CASA model at region scale[J].Transactions of the Chinese Society of Agricultural Engineering,2017,33(4):225-233. (in Chinese)
[46] Wang Z,Xiao X,Yan X.Modeling gross primary production of maize cropland and degraded grassland in northeastern China[J].Agricultural and Forest Meteorology,2010,150(9): 0-1167.
[47] 李正泉.陆地生态系统生产力的多尺度分析与尺度转换方法研究[D].北京:中国科学院地理科学与资源研究所,2006.
Li Z Q.Muti-scale analysis and scale changing of productivity of terrestrial ecosystem[D].Beijing:Institute of Geographic Sciences and Natural Resources Research,CAS, 2006.(in Chinese)
[48] Ruimy A,Dedieu G,Saugier B.TURC:a diagnostic model of continental gross primary productivity and net primary productivity[J].Global Biogeochemical Cycles,1996,10(2): 269-285.
[49] 付刚,周宇庭,沈振西,等.不同海拔高度高寒草甸光能利用效率的遥感模拟[J].生态学报,2010,31(23):6989-6998.
Fu G,Zhou Y T,Shen Z X,et al.Satellite-based modelling light use efficiency of alpine meadow along an altitudinal gradient[J].Acta Ecologica Sinica,2010,31(23):6989-6998. (in Chinese)
[50] Long S P,Zhu X G,Naidu S L,et al.Can improvement in photosynthesis increase crop yields[J].Plant,Cell & Environment,2006,29(3):315-330.
[51] 李少昆,王崇桃.作物株型和冠层结构信息获取与表述的方法(综述)[J].石河子大学学报:自然科学版,1997,1(3): 250-256.
Li S K,Wang C T.The methods of obtaining and expressing information of crop plant shape and population structure[J]. Journal of Shihezi University(Natural Science), 1997,1(3): 250-256.(in Chinese)
[52] Stewart D W,Costa C,Dwyer L M,et al.Canopy structure,light interception,and photosynthesis in maize[J]. Agronomy Journal,2003,95(6):1465-1474.
[53] Austin R B J,Bingham J,Blackwell R D,et al.Genetic improvements in winter wheat yields since 1900 and associated physiological changes[J].The Journal of Agricultural Science,1980,94(3):675-689.
[54] 胡延吉,陈化榜,赵檀方,等.山东省小麦品种性状演变的回顾与展望[J].山东农业科学,1991,5(1):1-6.
Hu Y J,Chen H B,Zhao T F,et al.Look back and forward to character evolution of wheat cultivars grown in Shangdong province[J].Shandong Agricultural Sciences,1991,5(1):1-6. (in Chinese)
[55] 姚维传,张从宇.小麦旗叶和芒的遗传及其与产量性状的相关性研究[J].种子,2000,5(43):19-21.
Yao W C,Zhang C Y.Inheritance of flag leaf and awn and their correlation with grain yield of wheat[J].Seed, 2000,5(43):19-21.(in Chinese)
[56] 张娟,崔党群,范平,等.小麦冠层结构与产量及其构成因素的典范相关分析[J].华北农学报,2000,15(3):39-44.
Zhang J,Cui D Q,Fan P,et al.Canonical correlations analysis between the canopy structures and yield with its components in wheat[J].Acta Agriculturae Boreali-Sinica, 2000,15(3):39-44.(in Chinese)
[57] Berry P M,Sylvester-bradley R,Berry S.Ideotype design for lodging-resistant wheat[J]. Euphytica, 2007,154(1-2):165- 179.
[58] 陈悦,王学华,廖轶,等.水稻叶片叶取向对光合能力的影响[J].植物生理与分子生物学学报,2002,28(5):396-398.
Chen Y,Wang X H,Liao Y,et al.Effect of leaf orientation on photosynthetic capacity in rice leaf[J].Journal Plant Physiology Molecular Biology,2002,28(5):396-398.(in Chinese)
[59] 曾浙荣,赵双宁,台建祥,等.北部冬麦区小麦产量育种的生理研究[J].北京农业科学,1998,16(3):1-6.
Zeng Z R,Zhao S N,Tai J X,et al.Physiological study on wheat yield breeding in northern winter wheat region[J]. Beijing Agricultural Sciences,1998,16(3):1-6.(in Chinese)
[60] 姚碧卿.小麦株型育种的研究及其成效[J].福建农业科技,1981(2):63.
Yao B Q.Study on plant type breeding of wheat and its effect[J].Fujian Journal of Agricultural Sciences,1981(2): 63.(in Chinese)
[61] 康祥波,钟阳和,张理,等.冬小麦群体叶层结构的研究[J].河南科技学院学报,1990,18(3):13-25.
Kang X B,Zhong Y H,Zhang L,et al.The study of the canopy architecture of winter wheat[J].Journal of Henan Institute of Science and Technology,1990,18(3):13-25.(in Chinese)
[62] 魏燮中,俞世蓉,吴纪民.小麦灌浆期中光能供求关系模拟与南京地区合理株型的探讨[J].南京农业大学学报,1988, 11(4):1-5.
Wei X Z,Yu S R,Wu J M.Simulation of energy accumulation process in wheat crop stand during grain filling period and the optimum plant type in Nanjing region[J].Journal of Nanjing Agricultural University,1988, 11(4):1-5.(in Chinese)
[63] 刘兆晔,于经川,姜鸿明,等.小麦理想株型的探讨[J].中国农学通报,2010,26(8):137-141.
Liu Z Y,Yu J C,Jiang H M,et al.A discussion on ideal plant type of wheat[J].Chinese Agricultural Science Bulletin, 2010,26(8):137-141.(in Chinese)
[64] Easson D L,White E M,Pickles S J.The effects of weather,seed rate and cultivar on lodging and yield in winter wheat[J].The Journal of Agricultural Science, 1993,121(2):145-156.
[65] Berry P M,Sterling M,Baker C J,et al.A calibrated model of wheat lodging compared with field measurements[J]. Agricultural and Forest Meteorology,2003,119(3):167-180.
[66] 凌启鸿.稻麦研究新进展[M].南京:东南大学出版社,1991.
Ling Q H.New progress in rice and wheat research [M].Nanjing:Southeast University Press,1991.(in Chinese)
[67] Tambussi E A,Bort J,Juan J G,et al.The photosynthetic role of ears in C3 cereals:metabolism,water use efficiency and contribution to grain yield[J].Critical Reviews in Plant Sciences,2007,26(1):1-16.
[68] Araus J L,Brown H R,Febrero A,et al.Ear photosynthesis, carbon isotope discrimination and the contribution of respiratory CO2to differences in grain mass in durum wheat[J].Plant,Cell & Environment,2006,16(4):383-392.
[69] 张永平,王志敏,王璞,等.冬小麦节水高产栽培群体光合特征[J].中国农业科学,2003,36(10):1143-1149.
Zhang Y P,Wang Z M,Wang P,et al.Canopy photosynthetic characteristics of population of winter wheat in water-saving and high-yielding cultivation[J].Scientia Agricultura Sinica,2003,36(10):1143-1149.(in Chinese)
[70] Evans L T,Wardlaw I F,Fischcr R A.In crop physiology: somecase historics(ed LT Evans)[M].Cambridge University Press,1992:101-150.
[71] Khaliq I,Irshad A,Ahsan M.Awns and flag leaf contribution towards grain yield in spring wheat(L.)[J].Cereal Research Communications,2008, 36(1):65-76.
[72] 陈磊,索全义,王金莲,等.春小麦非叶器官光合特性及与籽粒蛋白质关系的氮肥调控效应[J].华北农学报,2010, 25(5):198-201.
Chen L,Suo Q Y,Wang J L,et al.Regulating effect of nitrogen fertilizer on photosynthetic characteristics of non-leaf organs of spring wheat and relationship between grain protein[J].Acta Agriculturae Boreali-Sinica,2010, 25(5):198-201.(in Chinese)
[73] Dong C,Fu Y,Liu G,et al.Low light intensity effects on the growth,photosynthetic characteristics,antioxidant capacity, yield and quality of wheat(L.) at different growth stages in BLSS[J].Advances in Space Research,2014,53(11):1557-1566.
[74] Demmig-adams B,Adams W W.Photoprotection and other responses of plants to high light stress[J].Annual Review Plant Biology,1992,43(1):599-626.
[75] 张永强,陈传信,方辉,等.弱光下种植密度对冬小麦冠层温湿度及籽粒灌浆的影响[J].中国农业气象,2019,40(5): 301-307.
Zhang Y Q,Chen C X,Fang H,et al.Effects of planting population under poor light on the canopy temperature and humidity and grain-filling properties of winter wheat[J].Chinese Journal of Agrometeorology,2019,40(5): 301-307.(in Chinese)
[76] 李义博,宋贺,周莉,等.C4植物玉米的光合-光响应曲线模拟研究[J].植物生态学报,2017,41(12):1289-1300.
Li Y B,Song H,Zhou L,et al.Modeling study on photosynthetic-light response curves of a C4 plant, maize[J].Chinese Journal of Plant Ecology,2017,41(12): 1289-1300.(in Chinese)
[77] 林植芳,彭长连,孙梓健,等.光强对4种亚热带森林植物光合电子传递向光呼吸分配的影响[J].中国科学:生命科学,2000,30(1):72-77.
Lin Z F,Peng C L,Sun Z J,et al.Effects of light intensity on the distribution of photosynthetic electron transport to photorespiration in four subtropical forest plants[J].Scientia Sinica Vitae,2000,30(1):72-77.(in Chinese)
[78] 孙旭生,林琪,赵长星,等.施氮量对超高产冬小麦灌浆期旗叶光响应曲线的影响[J].生态学报,2009,29(3):378-387.
Sun X S,Lin Q,Zhao C X,et al.Effects of nitrogen application rate on light -response curves of flag leaves in super-high yielding winter wheat at grain filling stage[J].Acta Ecologica Sinica,2009,29(3):378-387.(in Chinese)
[79] 李华伟.遮光和渍水对小麦产量和品质的影响及其生理机制[D].南京:南京农业大学,2011.
Li H W.Grain yield and quality in wheat as affected by shading and waterlogging and the unfderlying eco- physiology priciples[D].Nanjing:Nanjing Agricultural University,2011.(in Chinese)
[80] 韩刚,赵忠.不同土壤水分下4种沙生灌木的光合光响应特性[J].生态学报,2010,30(15):4019-4026.
Han G,Zhao Z.Light response characteristics of photosynthesis of four xerophilous shrubs under different soil moistures[J].Acta Ecologica Sinica,2010,30(15): 4019- 4026.(in Chinese)
[81] 许大全,高伟,阮军.光质对植物生长发育的影响[J].植物生理学报,2015,51(8):1217-1234.
Xu D Q,Gao W,Ruan J.Effects of light quality on plant growth and development[J].Plant Physiology Communications, 2015,51(8):1217-1234.(in Chinese)
[82] 李慧敏,陆晓民,高清海,等.不同光质对黄秋葵幼苗生长、光合色素和气孔特征的影响[J].草业学报,2016,25(6): 62-70.
Li H M,Lu X M,Gao Q H,et al.Effects of different light qualities on the growth, photosynthetic pigments and stomatal characteristics of okra(Abelmoschus esculentus) seedlings[J].Acta Prataculturae Sinica,2016,25(6):62-70.(in Chinese)
[83] 张立伟,刘世琦,张自坤,等.不同光质下香椿苗的生长动态[J].西北农业学报,2010,19(6):115-119.
Zhang L W,Liu S Q,Zhang Z K,et al.Dynamic of different qualities on growth ofseedlings[J].Acta Agriculturae Boreali-occidentalis Sinica,2010,19(6):115- 119.(in Chinese)
[84] 马丙菊,常雨晴,景文疆,等.水稻水分高效利用的机理研究进展[J].中国稻米,2019,25(3):15-20.
Ma B J,Chang Y Q,Jing W J,et al.Research progress on the mechanism of high water use efficiency in rice[J].China Rice,2019,25(3):15-20.(in Chinese)
[85] Zhang X X,Yang L L,Hua K.Effects of CO2concentration on tall fescue drought resistance and water use efficiency[J].Acta Agrestia Sinica.2015.
[86] Ainsworth E A,Rogers A,Nelson R,et al.Testing the “source-sink” hypothesis of down-regulation of photosynthesis in elevated CO2in the field with single gene substitutions in glycine max[J].Agricultural and Forest Meteorology,2004,122(1-2):85-94.
[87] 勾俊英.CO2浓度升高与水分胁迫对小麦光合生理特征的影响研究[D].太古:山西农业大学,2018.
Gou J Y.Effects of elevated CO2concentration and water shortage on photosynthetic physiological characteristics of wheat[D].Taigu:Shanxi Agricultural University,2018.(in Chinese)
[88] 韩雪,郝兴宇,王贺然,等.FACE条件下冬小麦生长特征及产量构成的影响[J].中国农学通报,2012,28(36):154-159.
Han X,Hao X Y,Wang H R,et al.Effect of free air CO2enrichment(FACE) on the growth and grain yield of winter wheat[J].Chinese Agricultural Science Bulletin,2012,28 (36):154-159.(in Chinese)
[89] 汪东炎,郭李萍,李豫婷,等.大气CO2浓度升高对不同穗型冬小麦灌浆动态的影响[J].中国农业气象,2019,40(5): 284-292.
Wang D Y,Guo L P,Li Y T,et al.Effects of elevated atmospheric CO2concentration on grain filling dynamics of different spike-type winter wheat[J].Chinese Journal of Agrometeorology,2019,40(5):284-292.(in Chinese)
[90] Furbank R T,von Caemmerer S,Sheehy J,et al.C4 rice:a challenge for plant phenomics[J].Functional Plant Biology,2009,36(11):845-856.(in Chinese)
[91] IPCC.Climate change 2013:the physical science basis[M]. Cambridge:Cambridge University Press,2013.
[92] Tian Y,Chen J,Chen C,et al.Warming impacts on winter wheat phenophase and grain yield under field conditions in Yangtze Delta Plain,China[J].Field Crops Research,2012, 134:193-199.
[93] 吴冰洁,王靖,唐建昭,等.华北平原冬小麦产量变异的气象影响因子分析[J].中国农业气象,2018,39(10):623-635.
Wu B J,Wang J,Tang J Z,et al.Meteorological influencing factors on variation in winter wheat yield in the North China Plain[J].Chinese Journal of Agrometeorology. 2018,39(10):623-635.(in Chinese)
[94] Rezaei E E,Siebert S,Ewert F.Intensity of heat stress in winter wheat-phenology compensates for the adverse effect of global warming[J].Environmental Research Letters,2015, 10(2):024012.
[95] Tao F,Reimund P,Palosuo T,et al.Designing future barley ideotypes using a crop model ensemble[J].European Journal of Agronomy,2017,82:144-162.
[96] Li Y B,Hou R X,Tao F L.Wheat morpho-physiological traits and radiation use efficiency under interactive effects of warming and tillage management[J].Plant Cell & Environment,2020,44:2386-2401.
[97] Peng S,Laza R C,Visperas R M,et al.Grain yield of rice cultivars and lines developed in the philippines since 1966[J].Crop Science,2000,40(2):307-314.
[98] Tao F L,Zhang S,Zhang Z.Spatiotemporal changes of wheat phenology in China under the effects of temperature, day length and cultivar thermal characteristics[J].European Journal of Agronomy,2012,43:201-212.
[99] Tao F L,Zhang S,Zhang Z,et al.Maize growing duration was prolonged across China in the past three decades under the combined effects of temperature, agronomic management, and cultivar shift[J].Global Change Biology,2014,20(12): 3686-3699.
[100] 赵彦茜,肖登攀,柏会子,等.中国作物物候对气候变化的响应与适应研究进展[J].地理科学进展,2019,38 (2): 74-85.
Zhao Y X,Xiao D P,Bai H Z,et al.Research progress on the response and adaptation of crop phenology to climate change in China[J].Progress in Geography,2019,38(2): 74-85.(in Chinese)
[101] Boyer J S.Plant productivity and environment[J]. Science, 1982,218:443-448.
[102] 徐海成.冬小麦高产高效群体构建的栽培模式研究[D].泰安:山东农业大学,2016.
Xu H C.Cultivation strategy for constructing winter wheat population with high yield and high efficiency[D]. Tai’an:Shandong Agricultural University, 2016. (in Chinese)
[103] 王宏杰,李文阳,邵庆勤,等.灌浆期涝害对小麦旗叶光合特性影响及降渍恢复效应[J].中国农业气象,2019, 40(7):460-466.
Wang H J,Li W Y,Shao Q Q,et al.Effect of waterlogging on photosynthetic characteristics of wheat flag leaves during grain filling and recovery effect of water stress relief[J].Chinese Journal of Agrometeorology.2019,40(7): 460-466.(in Chinese)
[104] Xu Z Z,Yu Z W.Nitrogen metabolism in flag leaf and grain of wheat in response to irrigation regimes[J].Journal of Plant Nutrition and Soil Science,2006,169(1),118-126.
[105] 杨晓亚,于振文,许振柱.灌水量和灌水时期对小麦耗水特性和氮素积累分配的影响[J].生态学报,2007,29(2): 846-853.
Yang X Y,Yu Z W,Xu Z Z.Effects of irrigation regimes on water consumption characteristics and nitrogen accumulation and allocation in wheat[J].Acta Ecologica Sinica,2007,29(2):846-853.(in Chinese)
[106] 李运生,王菱,刘士平,等.土壤-根系界面水分调控措施对冬小麦根系和产量的影响[J].生态学报,2002,20(10): 1680-1687.
Li Y S,Wang L,Liu S P,et al.The influence of different amount of water supplied at different depths in soil-root interface on root distribution and yield of winter wheat[J].Acta Ecologica Sinica,2002,20(10):1680-1687. (in Chinese)
[107] 刘荣花,成林,胡程达.冬小麦根冠指标对干旱持续发展的响应[J].中国农业气象,2019,40(11):702-711.
Liu R H,Chen L,Hu C D.Response of root-canopy index of winter wheat on persist drought[J].Chinese Journal of Agrometeorology.2019,40(11):702-711.(in Chinese)
[108] 王俊儒,李生秀.不同生育时期水分有限亏缺对冬小麦产量及其构成因素的影响[J].西北植物学报,2000,20(2): 193-200.
Wang J R,Li S X.Effect of water-limited deficit stress in different growth stages on winter wheat grain yields and their yield constituents[J].Acta Botanica Boreali- Occidentalia Sinica,2000,20(2):193-200.(in Chinese)
[109] 房全孝,陈雨海,李全起,等.土壤水分对冬小麦生长后期光能利用及水分利用效率的影响[J].作物学报,2006,32 (6):861-866.
Fang Q X,Chen Y H,Li Q Q,et al.Effects of soil moisture on radiation utilization during late growth stages and water use efficiency of winter wheat[J].Acta Agronomica Sinica,2006,32(6):861-866.(in Chinese)
[110] Philip E,Mustafa P.Crop row spacing and its influence on the partitioning of evapotranspiration by winter-grown wheat in northern syria[J].Plant and Soil,2005,268(1), 195-208.
[111] Xu Z Z,Zhou G S.Combined effects of water stress and high temperature on photosynthesis,nitrogen metabolism and lipid peroxidation of a perennial grass Leymus chinensis[J].Planta,2006,224(5):1080-1090.
[112] 董博,张绪成,张东伟,等.水氮互作对春小麦叶片叶绿素含量及光合速率的影响[J].干旱地区农业研究,2012, 30(6):88-93.
Dong B,Zhang X C,Zhang D W,et al.Effect of water and nitrogen interaction on chlorophyll content and photosynthetic rate of spring wheat leaves[J].Agricultural Research in the Arid Areas,2012,30(6):88-93.(in Chinese)
[113] 江晓东,李旭冉,张涛,等.春小麦冠层氮素垂直分布与转运特征[J].中国农业气象,2021,42(3):190-199.
Jiang X D,Li X R,Zhang T,et al.Vertical distribution and transport characteristics of nitrogen in spring wheat canopy[J].Chinese Journal of Agrometeorology,2021, 42(3):190-199.(in Chinese)
[114] Evans L T.Crop evolution,adaption and yield[M]. Cambridge UK:Cambridge University Press,1993.
[115] Hikosaka K,Anten N P R,Borjigidai A,et al.A meta-analysis of leaf nitrogen distribution within plant canopies[J].Annals of Botany,2016,118:239-247.
[116] Muryono M,Chen C P,Sakai H,et al.Nitrogen distribution in leaf canopies of high-yielding rice cultivar takanari[J]. Crop Science,2017,57:2080-2088.
[117] 党蕊娟,李世清,穆晓慧,等.施氮对半湿润农田冬小麦冠层叶片氮素含量和叶绿素相对值垂直分布的影响[J].西北植物学报,2008,28(5):1036-1042.(in Chinese)
Dang R J,Li S Q,Mu X H,et al.Effect of nitrogen on nitrogen vertical distribution and chlorophyll relative value of winter wheat canopy in sub-humid areas[J].Acta Botanica Boreali-Occidentalia Sinica,2008,28(5):1036- 1042.
[118] 孙曦.植物营养原理[M].北京:中国农业出版社,1997.
Sun X.Principles of plant nutrition[M].Beijing:China Agriculture Press,1977.(in Chinese)
[119] Massignam A M,Chapman S C,Hammer G L,et al.Physiological determinants of maize and sunflower grain yield as affected by nitrogen supply[J].Field Crops Research,2009,113(3):256-267.
[120] 陈雨海,余松烈,于振文.小麦生长后期群体光截获量及其分布与产量的关系[J].作物学报,2003,29(5):91-95.
Chen Y H,Yu S L,Yu Z W.Relationship between amount or distribution of par interception and grain output of wheat communities[J].Acta Agronomica Sinica,2003, 29(5):91-95.(in Chinese)
[121] Dai X,Zhou X,Jia D,et al.Managing the seeding rate to improve nitrogen-use efficiency of winter wheat[J].Field Crops Research,2013,154:100-109.
[122] 李全起,陈雨海,吴巍,等.秸秆覆盖和灌溉对冬小麦农田光能利用效率的影响[J].应用生态学报,2006,17(2): 243-246.
Li Q Q,Chen Y H,Wu W,et al.Effects of straw mulching and irrigation on solar energy utilization efficiency of winter wheat farmland[J].Chinese Journal of Applied Ecology,2006,17(2):243-246.(in Chinese)
[123] 陈素英,张喜英,毛任钊,等.播期和播量对冬小麦冠层光合有效辐射和产量的影响[J].中国生态农业学报,2009,17(4):681-685.
Chen S Y,Zhang X Y,Mao R Z,et al.Effect of sowing date and rate on canopy intercepted photo-synthetically active radiation and yield of winter wheat[J].Chinese Journal of Eco-Agriculture,2009,17(4):681-685.(in Chinese)
[124] 王东兴.春小麦复种油葵高产栽培技术[J].现代农业,2018,(10):45.
Wang D X.High yield cultivation techniques of spring wheat multiple cropping oil sunflower[J].Modern Agriculture,2018,(10):45.(in Chinese)
[125] 刘桐华,余彦波.提高小麦光能利用效率的一个重要途径[J].北京农业科学,1989(4):1-4.
Liu T H,Yu Y B.An important way to improve the utilization of light energy in wheat[J].Beijing Agricultural Science,1989(4):1-4.(in Chinese)
[126] Parry M A,Andralojc P J,Mitchell R A,et al.Manipulation of rubisco:the amount,activity,function and regulation[J]. Journal of Experimental Botany,2003,54(386):1321- 1333.
[127] Pyke K A,Leech R M.The control of chloroplast number in wheat mesophyll cells[J].Planta,1987,170(3):416-420.
[128] Makino A,Harada M,Kaneko K,et al.Whole-plant growth and N allocation in transgenic rice plants with decreased content of ribulose-1,5-bisphosphate carboxylase under different CO2partial pressures[J].Functional Plant Biology, 2000,27(1):1-12.
[129] Suzuki Y,Ohkubo M,Hatakeyama H,et al.Increased Rubisco content in transgenic rice transformed with the ‘sense’rbcS gene[J].Plant and Cell Physiology,2007, 48(4):626-637.
[130] Kimball B A.Carbon dioxide and agricultural yield:an assemblage and analysis of 430 prior observations1[J]. Agronomy Journal,1983,75(5):779-788.
[131] Nakano H,Makino A,Mae T.The effect of elevated partial pressures of CO2on the relationship between photosynthetic capacity and N content in rice leaves[J]. Plant Physiology,1997,115(1):191-198.
[132] Medrano H,Keys A J,Lawlor D W,et al.Improving plant production by selection for survival at low CO2concentrations[J].Journal of Experimental Botany,1995, 46(special):1389-1396.
[133] Kebeish R,Niessen M,Thiruveedhi K,et al.Chloroplastic photorespiratory bypass increases photosynthesis and biomass production in arabidopsis thaliana[J].Nature Biotechnology,2007,25(5):593-599.
[134] Gowik U,Westhoff P.The path from C3 to C4 photosynthesis[J].Plant Physiology,2011,155(1):56-63.
[135] Mitchell P L,Sheehy J E.Supercharging rice photosynthesis to increase yield[J].New Phytologist,2006, 171(4):688-693.
[136] Kaisa K,Sarah C,Shanta K,et al.Strategies for engineering a two-celled C(4) photosynthetic pathway into rice. Journal of Experimental Botany,2011,9:3001-3010.
[137] Mitsue M,Chisato M,Shin-Ichi M,et al.Lessons from engineering a single-cell C(4) photosynthetic pathway into rice. Journal of Experimental Botany,2011,9:3021.
[138] Edwards G E,Voznesenskaya E,Smith M,et al.Breaking the Kranz paradigm in terrestrial C4 plants:does it hold promise for C4 rice? In Charting new pathways to C4 rice[M].Los Banos,Philippines:International Rice Research Institute,2007.
[139] Hibberd J M,Sheehy J E,Langdale J A.Using C4 photosynthesis to increase the yield of rice-rationale and feasibility[J].Current Opinion in Plant Biology,2008, 11(2),228-231.
[140] Ortiz R,Sayre K D,Govaerts B,et al.Climate change: Can wheat beat the heat[J].Agriculture Ecosystems & Environment,2008,126(1-2):46-58.
[141] IPCC.Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change[M].Cambridge,UK:Cambridge University Press,2008.
[142] Reynolds M P,Borlaug N E.Impacts of breeding on international collaborative wheat improvement[J].Journal of Agricultural Science,2006,144(1):3-17.
[143] Hodson D,White J.Climate change:what future wheatIn:Dixon J, Braun H J, Kosina P.(Eds.) Wheat Facts and Futures[M].Mexico,D.F:CIMMYT.
[144] Blum A.Drought resistance,water-use efficiency,and yield potential-are they compatible,dissonant,or mutually exclusive[J].Australian Journal of Agricultural Research, 2005,56(11),1159-1168.
[145] Gu J,Yin X,Zhang C,et al.Linking ecophysiological modelling with quantitative genetics to support marker-assisted crop design for improved yields of rice() under drought stress[J].Annals of Botany,2014,114:499-511.
[146] 郭庆华,杨维才,吴芳芳,等.高通量作物表型监测:育种和精准农业发展的加速器[J].中国科学院院刊,2018,33(9): 56-62.
Guo Q H,Yang W C,Wu F F,et al.High-throughput crop phenotyping:accelerators for development of breeding and precision agriculture[J].Bulletin of Chinese Academy of Sciences,2018,33(9):56-62.(in Chinese)
[147] Reynolds M,Manes Y,Izanloo A,et al.Phenotyping approaches for physiological breeding and gene discovery in wheat[J].Annals of Applied Biology,2009,155(3): 309-320.
[148] Gastón Q,Victoria B,Sebastián F,et al.Light-use efficiency and energy partitioning in rice is cultivar dependent[J].Photosynthesis Research,2018,140:51-63.
[149] Driever S M,Lawson T,Andralojc P J,et al.Natural variation in photosynthetic capacity,growth,and yield in 64 field-grown wheat genotypes[J].Journal of Experimental Botany,2014,65:4959-4973.
Research Progress on the Mechanism of High Light Use Efficiency in Wheat
LI Yi-bo, TAO Fu-lu
(Institute of Geographic Sciences and Natural Resources Research/Key Laboratory of Land Surface Pattern and Simulation/China University of Chinese Academy of Sciences, Beijing 100049, China)
Crop yield is dependent on crop biomass and harvest index. The increases in crop yield in the past benefit mainly from the increase in harvest index, the increase in biomass has been small. Light use efficiency is a bottleneck to further increase crop biomass and yield. Wheat (L.) is a widely cultivated cereal crop globally, providing energy to up to 20% of the world's population. It is necessary to reveal the intrinsic mechanism and extrinsic factors affecting the light use efficiency of wheat, which is important for improving the utilization efficiency of crop resources and productivity. Achieving high light efficient production of wheat under limited natural resources has become the focus of study at home and abroad. Here, the definition of light use efficiency, the main processes, the characteristics of light use efficiency in wheat were summarized from relevant references, showing a more significant potential to improve light use efficiency. The effects of external factors such as light, water, nutrient and tillage system on wheat light use efficiency were summarized. The results indicated that light use efficiency was regulated by internal factors such as photosynthesis at the individual plant scale, and by abiotic factors such as temperature, precipitation and tillage practices at the field scale. The current problems and the adaptation mechanism of wheat in the context of climate change were also analyzed, aiming to provide a theoretical reference for the practical research of the high light use efficiency of wheat. Future research on light use efficiency can use high-throughput phenotypic observation techniques combined with molecular markers to design ideotypes in the target environment, providing scientific evidence for crop breeding.
Wheat; Climate change; Light use efficiency; Ideotypes; Yield potential
10.3969/j.issn.1000-6362.2022.02.002
李义博,陶福禄.提高小麦光能利用效率机理的研究进展[J].中国农业气象,2022,43(2):93-111
收稿日期:2021−06−09
国家自然科学基金项目“提升小麦光利用效率和产量潜力的生理遗传基础与理想株型设计研究”(3176113006)
通讯作者:陶福禄,博士,研究员,主要从事生态系统与全球变化生态学研究,E-mail: taofl@igsnrr.ac.cn
李义博,E-mail: lybmails@163.com
