南粳3908生育后期剑叶PSⅡ荧光日变化特性及叶绿体超微结构观察
2022-02-06魏晓东张亚东宋雪梅赵庆勇梁文化黄胜东谢寅峰王才林
魏晓东, 张亚东, 宋雪梅,, 陈 涛, 朱 镇, 赵庆勇, 赵 凌, 路 凯, 梁文化, 赫 磊, 黄胜东, 谢寅峰, 王才林
(1.江苏省农业科学院粮食作物研究所/江苏省优质水稻工程技术研究中心/国家水稻改良中心南京分中心,江苏南京210014;2.南京林业大学/南方现代林业协同创新中心, 江苏南京210037)
水稻是中国重要的粮食作物,江苏省是中国粳稻主产省之一,全省粳稻种植面积达2×106hm2以上,仅次于黑龙江省,占全国粳稻总种植面积的22%左右;粳稻年产量约为1.7×107t,约占全国粳稻总产量的24%,在保障江苏省乃至中国粮食安全中的地位举足轻重[1-2]。进入21世纪以来,江苏省水稻育种目标从高产向优质转变,加上高质高效配套栽培新技术的应用,江苏省水稻品种的产量、抗性和品质都有了较大提升,尤其是粳稻品种的食味品质得到显著改良,育成了一批产量潜力大、食味品质好的粳稻品种[3-7]。南粳3908是江苏省农业科学院粮食作物研究所利用南粳5055的自然变异植株,进行多代农艺性状选择及食味品质、穗颈瘟抗性分子标记辅助筛选技术,于2013年育成的一个综合性状好、产量潜力大、食味品质优的早熟晚粳稻新品种。南粳3908与南粳5055一样,直链淀粉含量低,制成的米饭口感软糯,润滑,富有弹性,冷不回生,食味极佳,该品种大穗大粒,丰产性好,穗颈瘟抗性得到改良。2019、2020年农业农村部组织专家对南粳3908百亩方进行了实产验收,单产都超过11 700 kg/hm2,达到了“超级稻”的产量标准,2021年,南粳3908通过农业农村部的超级稻认定。
水稻产量的形成是水稻叶片通过光合作用固定CO2合成光合产物的结果。水稻产量的60%~80%来自水稻抽穗后叶片光合作用形成的光合作用产物[8]。叶绿素荧光是决定光合效率的主要因素,研究光合效率有多种方法,其中通过无损检测植物光合作用的能量吸收、传递、耗散和分配是目前研究叶绿素荧光动力学过程最先进的方法,通过叶绿素荧光方法可以诊断光合结构的运转状况[9]。作物叶绿素荧光参数在不同品种间的差异归根结底是由基因型不同造成的。因此,通过叶绿素荧光分析,可以测定叶片的光合效率,从而为选育、鉴定高光效优良品种提供依据[10]。水稻在灌浆期经常遭受高温强光天气,而植物在正午强光下会发生光合午休现象,检测水稻剑叶1天中不同时段的叶绿素荧光的变化及光系统Ⅱ(PSⅡ)核心蛋白质表达量的变化,能够准确反映水稻植株光系统的性能特点。而叶绿素荧光动力学特性与叶绿体的超微结构有直接关系。但是,目前有关高产粳稻PSⅡ荧光特性及其与叶绿体超微结构关系的报道不多。因此,本研究以超级稻品种南粳3908及其亲本南粳5055和生产上大面积种植的淮稻5号为材料,研究其孕穗期到花后35 d剑叶光合作用及荧光的日变化特性、光系统Ⅱ反应中心蛋白质CP43、CP47的表达变化及叶绿体结构,解析南粳3908高产的光合生理机制,以期为高产优质水稻品种的选育提供参考依据。
1 材料与方法
1.1 供试品种与试验设计
试验材料为江苏省农业科学院粮食作物研究所选育的超级稻品种南粳3908,以南粳5055及江苏省大面积推广的广适性水稻品种淮稻5号为对照。南粳5055既是南粳3908的亲本,又是江苏省生产上大面积种植的晚粳稻品种,生育期与南粳3908相同,是2014年原农业部确认的超级稻品种[11]。
本试验于2020-2021年在江苏省农业科学院粮食作物研究所南京试验基地进行。每年5月18日播种,浸种催芽后用塑料盘育秧,6月18日移栽,按随机区组设计,共设3次重复,每个小区共9行,每行20穴,每穴栽2苗,行株距30 cm×13 cm。施纯氮270.0 kg/hm2,基肥∶分蘖肥∶穗肥按照4∶3∶3质量比施用。基肥用525.0 kg/hm2N-P2O5-K2O(质量比20∶12∶16)复合肥,分蘖肥分2次施用,移栽后7 d施112.5 kg/hm2尿素,再过7 d追施75.0 kg/hm2尿素。穗肥分2次施用,在倒4叶刚长出时施225.0 kg/hm2N-P2O5-K2O(质量比20∶12∶16)复合肥,10 d后追施75.0 kg/hm2尿素。在水分管理方面,除了移栽后7 d、孕穗至扬花阶段浅水灌溉外,其余时间均采用干湿交替的湿润灌溉方式。病虫草害防治方法与生产大田相同。
1.2 测定项目
1.2.1 产量性状的测定 水稻成熟后,在每个小区第3行、第7行两头第3穴开始各取5穴植株样品,带回室内自然风干后考种。测定项目包括每穴穗数、每穗实粒数、每穗空瘪粒数、结实率、千粒质量和每穴产量。将各小区稻谷全部收获后晒干,称质量并测定小区产量,折算成单位面积产量。
1.2.2 叶片净光合速率的测定 每个品种的植株在剑叶抽出时挂牌标记,选择同一天抽出的主茎剑叶,分别在每个品种的孕穗期(开花前7 d)、开花期与花后7 d、14 d、21 d、28 d、35 d晴天的09:00-10:00采用美国Li-6400便携式光合测定仪测定各品种剑叶上部1/3处的叶片净光合速率,测定时使用红蓝光源,光量子通量密度为1 000 μmol/(m2·s),每次随机测定10张剑叶,每张叶片重复测定3次。
1.2.3 叶片荧光参数的测定 光合色素的测定用SPAD仪,每个小区测定10张剑叶,每张剑叶测定5个不同部位。用Handy-PEA连续激发式荧光仪(Hansatech,UK)活体测量并快速得出叶绿素荧光诱导动力学曲线(O-J-I-P荧光诱导曲线)。每个品种分别在孕穗期、开花期与花后7 d、14 d、21 d、28 d、35 d的08:00、12:00、18:00进行叶绿素荧光测定。测定前将叶片暗适应20 min,然后暴露在饱和脉冲光下[光照度为3 000 μmol/(m2·s)],测得荧光曲线。每个重复测定10次。得到的荧光曲线利用JIP-test进行分析,荧光参数参照Strasser等[12-13]的方法计算。
1.2.4 蛋白质表达的测定 分别于花后21 d和花后35 d 08:00、12:00、18:00取各品种剑叶迅速冻于液氮中,保存于-80 ℃冰箱中用于后续膜蛋白提取和蛋白质免疫印迹试验。蛋白质提取和免疫印迹试验方法参照文献[14]。用化学发光成像仪(Tannon)检测,用Image J软件采集信号,重复测定3次。
1.2.5 叶绿体超微结构的观察 分别于花后21 d、35 d 09:00取各品种剑叶上端1/3处叶片,然后用刀片切成0.1 cm大小的组织,再用4 ℃ 4%戊二醛进行固定抽气,于4 ℃保存备用。观察前用0.1 mol/L磷酸缓冲液浸洗3次,每次15 min。样品经丙酮脱水、树脂包埋、切片、染色后用透射电子显微镜(Hitachi TEM 7650,日本日立高新技术公司产品)观察。
1.3 数据分析
用Excel 2016处理数据,用PEA Plus V1.10对荧光参数进行处理和计算,用SPSS 25.0进行方差分析,用最小显著性差异法(LSD)进行多重比较,用Excel作图。
2 结果与分析
方差分析结果表明,2020、2021年供试品种的产量和产量性状、干物质质量及光合性状在品种间都有显著差异,且品种间的变化趋势一致,年份间和年份与品种间的互作效应都不显著。因此取2年数据的平均值进行多重比较分析。
2.1 南粳3908及对照间产量性状的差异
由表1可知,在穗数方面,淮稻5号显著多于南粳3908、南粳5055;在每穗粒数方面,南粳3908、南粳5055均显著多于淮稻5号,南粳3908和南粳5055间无显著差异;结实率表现出与每穗粒数相似的趋势;在千粒质量方面,南粳3908显著高于南粳5055、淮稻5号;在每穴粒质量、产量方面,排序均为南粳3908>南粳5055>淮稻5号。

表1 供试品种的产量性状
从图1a可以看出,各水稻品种剑叶绿素含量(SPAD值)均在花后7~14 d达到高峰,之后缓慢下降。其中南粳5055的剑叶叶绿素含量在花后均明显高于南粳3908、淮稻5号,淮稻5号、南粳3908的剑叶叶绿素含量在花后无明显差异,但在花后35 d,淮稻5号的剑叶叶绿素含量下降得最多,相对最高值下降了18.42%,此时南粳5055的剑叶叶绿素含量相对最高值下降了17.91%,南粳3908的剑叶叶绿素含量下降得最缓慢,在花后35 d剑叶的叶绿素含量仅比最高值下降了15.67%。
由图1b可以看出,各个水稻品种的净光合速率大致呈现先升高后下降的趋势,南粳3908、淮稻5号的剑叶净光合速率均在花后14 d达到峰值,南粳5055的剑叶净光合速率在花后7 d达到峰值,3个水稻品种间的净光合速率峰值无明显差异。南粳3908在花后14~28 d的剑叶净光合速率明显高于南粳5055;除花后14 d外,南粳3908在花后7~28 d的剑叶净光合速率明显高于淮稻5号,且南粳3908的剑叶净光合速率在花后7~28 d一直维持在14 μmol/(m2·s)以上,南粳5055的剑叶净光合速率在花后7~14 d维持在14 μmol/(m2·s)左右,淮稻5号的剑叶净光合速率只在花后14 d达到14 μmol/(m2·s)。
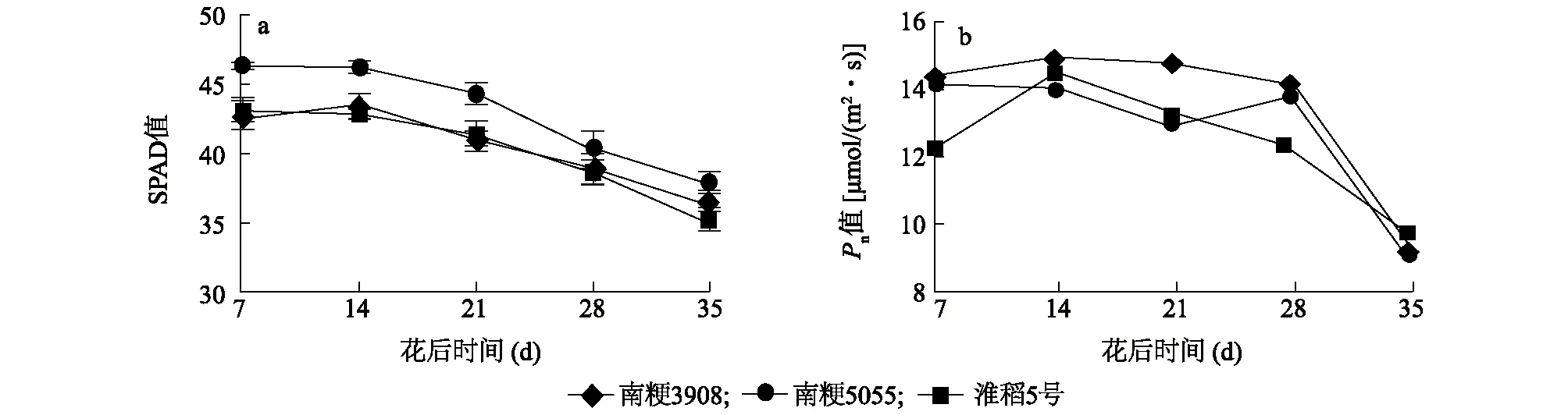
a:不同品种剑叶SPAD值;b:不同品种剑叶净光合速率。图1 南粳3908及对照品种开花后的SPAD值和剑叶净光合速率Fig.1 SPAD value and net photosynthetic rate in flag leaves of Nanjing 3908 and the controls after flowering
2.2 南粳3908及对照剑叶叶绿素荧光参数的日变化特性
植物叶片生理环境的改变会直接影响其光合作用,PSⅡ荧光参数的变化能够准确反映叶片的光合生理状态。图2~图15分别是各水稻品种在孕穗期、花后21 d、花后35 d的剑叶荧光参数日变化。由图2可以看出,各水稻品种的暗适应最小荧光值(F0)在不同时期的日变化均呈现先升高后降低的趋势,在孕穗期,淮稻5号的F0在1 d中的变化最剧烈,12:00测得的剑叶F0较08:00测得的剑叶F0增加了32.5%,其次为南粳5055,12:00测得的剑叶F0较08:00测得的剑叶F0增加了11.3%。测得的南粳3908剑叶F0变化幅度最小,12:00测得的剑叶F0较08:00测得的剑叶F0仅增加了8.3%。在花后21 d、35 d,各品种的剑叶F0间无明显差异。
Fm值为暗适应最大荧光值,各品种的Fm在不同时期的日变化均呈现先降低后升高的趋势,并且随着生育进程的推进,各品种水稻的剑叶暗适应最大荧光值逐渐降低,淮稻5号在孕穗期08:00的剑叶暗适应最大荧光值明显低于其他品种,在花后21 d、35 d,淮稻5号在12:00的剑叶暗适应最大荧光值较08:00下降得最多,且明显低于其他品种。南粳3908、南粳5055的剑叶Fm在各时期间的变化均无明显差异(图3)。

Fm:暗适应最大荧光值。图3 南粳3908及对照品种从孕穗期到开花后35 d剑叶的暗适应最大荧光值(Fm)的日变化Fig.3 Diurnal variation of dark adaptation maximum fluorescence (Fm) of flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering
比活性参数是评判作物光合器官对光能吸收、转化和耗散等状况的重要指标。由图4可以看出,不同水稻品种单位反应中心吸收的光能(ABS/RC值)在生育后期有升高的趋势,且在不同时期的日变化值均呈现先升高后降低的趋势,淮稻5号在各个时期12:00的ABS/RC值明显高于其他品种,淮稻5号的ABS/RC值在孕穗期变化最剧烈,该时期南粳5055ABS/RC值的变化次之,南粳3908ABS/RC值的变化幅度最小。

ABS/RC:单位反应中心吸收的光能。图4 南粳3908及对照品种从孕穗期到开花后35 d剑叶单位反应中心吸收的光能(ABS/RC)的日变化Fig.4 Diurnal variation of light energy absorbed by unit reaction center (ABS/RC) of flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering
由图5可知,PSⅡ系统中单位反应中心捕获的光能(TR0/RC值)与ABS/RC值的变化趋势相同,在生育后期升高,淮稻5号TR0/RC值与ABS/RC值在各时期的日变化最剧烈,尤其在孕穗期12:00的升高幅度最大,南粳5055次之,南粳3908的变化幅度最小。在花后21 d、35 d,淮稻5号在08:00、12:00的TR0/RC值明显高于其他品种,南粳5055和南粳3908间无明显差异。
由图6可知,单位反应中心用于电子传递的能量(ET0/RC值)在水稻生育后期有降低趋势,孕穗期淮稻5号的ET0/RC值在1 d中呈现先升高后下降的趋势,其他品种无明显变化。在花后21 d、35 d,各水稻品种ET0/RC值的日变化均呈现先下降后升高的趋势,其中淮稻5号在这2个时期的12:00下降得最多,在花后21 d、35 d的12:00,ET0/RC值分别较08:00下降了32.3%、32.5%。
由图7可知,各水稻品种单位反应中心热耗散的能量(DI0/RC值)在生育后期有增加趋势,8:00-18:00的DI0/RC值呈现先升高后下降的趋势,其中淮稻5号在各时期8:00-18:00的DI0/RC值变化程度最剧烈。在孕穗期12:00,淮稻5号的DI0/RC值较08:00升高了1.21倍,南粳5055的DI0/RC值较08:00升高了58.4%,南粳3908的DI0/RC值较08:00升高了26.9%,3个水稻品种12:00的DI0/RC值差异明显。在花后21 d、35 d的12:00,淮稻5号的DI0/RC值明显高于其他品种,南粳5055、南粳3908的DI0/RC值之间无明显差异。

TR0/RC:单位反应中心捕获的光能。图5 南粳3908及对照品种从孕穗期到开花后35 d剑叶PSⅡ系统中单位反应中心捕获的光能(TR0/RC)的日变化Fig.5 Diurnal variation of light energy captured by unit reaction center (TR0/RC) of flag leaves of Nanjing 3908 and the controls in PSⅡ system from booting stage to 35 days after flowering

ET0/RC:单位反应中心用于电子传递的能量。图6 南粳3908及对照品种从孕穗期到开花后35 d剑叶单位反应中心用于电子传递的能量(ET0/RC)的日变化Fig.6 Changes of energy used for electron transport captured by unit reaction center (ET0/RC) of flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering

DI0/RC:单位反应中心热耗散的能量。图7 南粳3908及对照品种从孕穗期到开花后35 d剑叶单位反应中心热耗散的能量(DI0/RC)的日变化Fig.7 Diurnal variation of energy dissipated per reaction center (DI0/RC) of flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering
由图8可知,各水稻品种单位截面吸收的光能(ABS/CSm值)随着生育进程的推进有降低的趋势,各时期、各品种的日变化大部分呈现先降低后升高的趋势。除孕穗期外,淮稻5号在各时期12:00的ABS/CSm值较08:00下降得最多,南粳5055和南粳3908各时期12:00的ABS/CSm值无明显差异。

ABS/CSm:单位截面吸收的光能。图8 南粳3908及对照品种从孕穗期到开花后35 d剑叶单位截面吸收的光能(ABS/CSm)的日变化Fig.8 Diurnal variation of light energy absorbed by unit cross section (ABS/CSm) of flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering
由图8、图9可知,单位截面捕获的光能(TR0/CSm值)与单位截面吸收的光能变化趋势一致,各水稻品种的TR0/CSm值在不同时期的日变化呈现先降低后升高的趋势,淮稻5号在各个时期12:00的TR0/CSm值较08:00降低得最多,各时期12:00南粳5055和南粳3908间的TR0/CSm无明显差异。
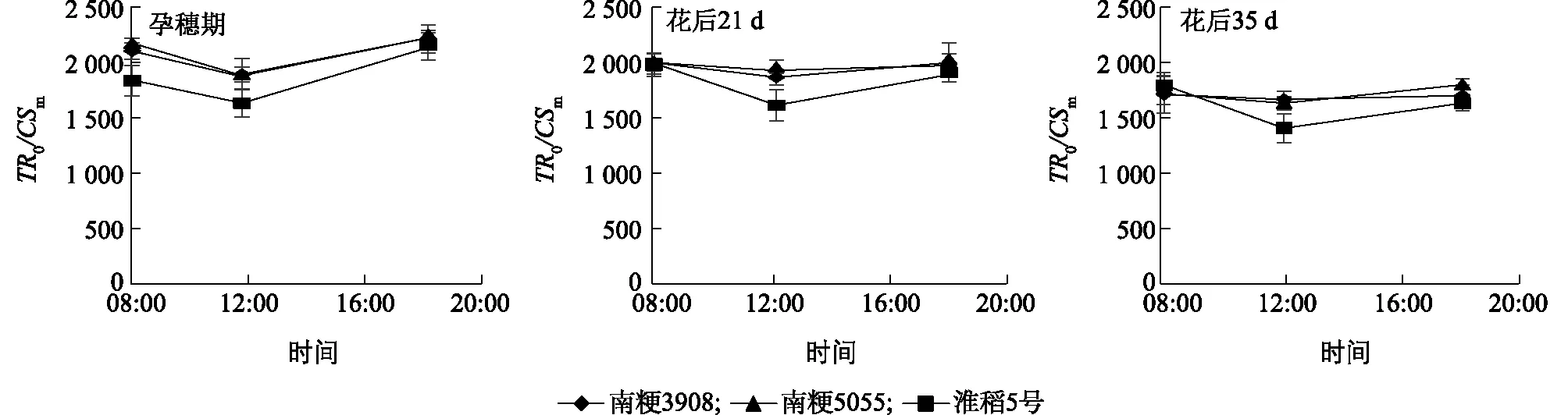
TR0/CSm:单位截面捕获的光能。图9 南粳3908及对照品种从孕穗期到开花后35 d剑叶单位截面捕获的光能(TR0/CSm)的日变化Fig.9 Diurnal variation of light energy captured by unit cross section (TR0/CSm) of flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering
由图10可知,各水稻品种单位截面用于电子传递的能量(ET0/CSm值)随着水稻生育进程的推进而逐渐降低,且在08:00-18:00呈现先降低后升高的趋势,淮稻5号在各个时期12:00 的ET0/CSm值最低,与其他品种间差异明显,且在花后21 d、35 d18:00的ET0/CSm值明显低于其他品种,在孕穗期和花后21 d 08:00的ET0/CSm值明显低于其他品种。在各个时期,南粳5055和南粳3908的ET0/CSm值间无明显差异。
由图11可知,各水稻品种单位截面用于热耗散的能量(DI0/CSm值)在各时期均呈现先升高后降低的趋势,不同水稻品种间的DI0/CSm值在孕穗期08:00-12:00差异明显,其他时期无明显差异,孕穗期淮稻5号的DI0/CSm值在12:00增加得最多,南粳5055次之,南粳3908增加得最少,三者间差异明显。
φE0表示用于电子传递的量子产额,由图12可知,各水稻品种的φE0在花后35 d有所降低,各时期的φE0在08:00-18:00均呈现先降低后升高的趋势,其中淮稻5号在各个时期12:00的φE0均小于其他品种,在花后21 d 08:00-18:00的φE0都小于其他品种,各时期南粳5055和南粳9108的φE0无明显差异。
M0表示OJIP荧光曲线诱导的初始斜率,由图13可知,各水稻品种在不同时期的M0有所不同,花后21 d的M0最低,花后35 d的M0最高。各水稻品种的M0在不同时期的日变化基本呈现先升高后降低的趋势,其中淮稻5号在各时期12:00的M0明显高于其他品种;在孕穗期和花后35 d,淮稻5号的M0在08:00-18:00均高于其他品种,南粳5055和南粳3908的M0在各时期、各时间段无明显差异。

ET0/CSm:单位截面用于电子传递的能量。图10 南粳3908及对照品种从孕穗期到开花后35 d剑叶单位截面用于电子传递的能量(ET0/CSm)的日变化Fig.10 Diurnal variation of energy used for electron transport per unit cross section (ET0/CSm) of flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering

DI0/CSm:单位截面用于热耗散的能量。图11 南粳3908及对照品种从孕穗期到开花后35 d剑叶单位截面用于热耗散的能量(DI0/CSm)的日变化Fig.11 Diurnal variation of energy used for heat dissipation per unit cross section (DI0/CSm) of flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering

φE0:用于电子传递的量子产额(当t=0时)。图12 南粳3908及对照品种从孕穗期到开花后35 d剑叶用于电子传递的量子产额(当t=0时)(φE0)的日变化Fig.12 Diurnal variation of quantum yield (t=0) for electron transport (φE0) in flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering

M0:OJIP荧光曲线诱导的初始斜率。图13 南粳3908及对照品种从孕穗期到开花后35 d剑叶OJIP荧光曲线诱导的初始斜率(M0)的日变化Fig.13 Diurnal variation of initial slope induced by OJIP fluorescence curve (M0) in flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering
Sm表示PQ库的大小,由图14可知,在不同时期,各水稻品种的Sm日变化值呈现先降低后升高的趋势,其中淮稻5号的Sm在孕穗期12:00下降得最多,与其他品种间差异明显,且其花后21 dSm的日变化均小于其他品种,各品种的Sm在不同时期间差异不大。
PIabs表示PSⅡ光能吸收的光合性能指数,各水稻品种在花后35 d的光合性能指数较早期有所降低,淮稻5号在各时期12:00的PIabs均明显低于其他品种,南粳3908、南粳5055在各时期的PIabs无明显差异(图15)。

Sm:标准化后的J-P相和直线F=Fm之间的面积。图14 南粳3908及对照品种从孕穗期到开花后35 d剑叶标准化后的J-P相和直线F=Fm之间面积(Sm)的日变化Fig.14 Diurnal variation of area between normalized J-P phase and straight line F=Fm (Sm) of flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering

PIabs:PSⅡ光能吸收的光合性能指数。图15 南粳3908及其对照品种从孕穗期到开花后35 d剑叶PSⅡ光能吸收的光合性能指数(PIabs)的变化Fig.15 Changes of photosynthetic performance index of PSⅡ light energy absorption (PIabs) in flag leaves of Nanjing 3908 and the controls from booting stage to 35 days after flowering
2.3 南粳3908及对照剑叶光合相关蛋白质表达的日变化特性
针对上述日变化的生理特性,对南粳3908、南粳5055、淮稻5号在花后21 d、35 d 08:00、12:00、18:00的光合相关蛋白质CP43、CP47的表达情况进行检测。结果表明,各水稻品种剑叶CP43、CP47蛋白的表达量都随叶片衰老表现出降低趋势,花后35 d的表达量低于花后21 d。光合相关蛋白质CP43、CP47在12:00的表达量较上午有降低趋势,但各品种的表现不一样。在花后21 d的08:00,各水稻品种CP43、CP47蛋白的表达量不尽相同,南粳3908中CP43、CP47蛋白的表达量明显高于南粳5055、淮稻5号;在12:00,各水稻品种CP43、CP47蛋白的表达量降低;在18:00,南粳3908 CP43、CP47蛋白的表达量明显高于其他品种。在花后35 d,南粳3908在08:00、18:00CP43、CP47蛋白的表达量明显高于其他品种,CP47蛋白在12:00的表达量和其他时间相比差异不大,可能由于此时期的正午温度和光照较花后21 d有所降低(图16)。

a:不同水稻品种剑叶CP43蛋白在花后21 d、花后35 d 08:00、12:00、18:00的相对表达量;b:不同水稻品种剑叶CP47蛋白在花后21 d、35 d 08:00、12:00、18:00的相对表达量。图16 南粳3908及对照品种开花后21 d、35 d剑叶CP43、CP47蛋白的相对表达量Fig.16 Relative expression levels of CP43 and CP47 in flag leaves of Nanjing 3908 and the controls at 21 d and 35 d after flowering
2.4 南粳3908及对照剑叶叶绿体超微结构的差异
叶绿体结构的完整和有序是植物进行光合作用的前提。利用透射电镜对3个水稻品种花后21 d、35 d剑叶的叶绿体结构进行观察。从图17可以看出,随着叶片衰老,各个水稻品种的叶绿体结构明显衰败,花后21 d,各个水稻品种叶绿体呈纺锤形或椭圆形,形态规则,类囊体基粒片层排列整齐,垛叠紧密,其中南粳3908的叶绿体类囊体基粒片层垛叠数多于南粳5055,淮稻5号基粒片层稍有模糊。在花后35 d,各个水稻品种的叶绿体形状相对不规则,类囊体结构开始变得松散,嗜锇颗粒增多、增大,从图17还可以看出,南粳3908叶绿体中的嗜锇颗粒较其他品种少,淮稻5号叶绿体中的嗜锇颗粒数最多,南粳3908叶绿体的内膜结构相对清晰,而南粳5055、淮稻5号叶绿体的内膜结构相对模糊不清,基粒和基质片层开始消融。
3 讨论
作物干物质的积累大都来自叶片的光合作用,光系统的高效运转是干物质积累的前提。稻谷产量的40%~60%来自剑叶的光合作用,而PSⅡ是光系统中重要的光能转化单位,其功能的好坏决定了水稻的光合性能,进而影响稻谷产量。叶绿素相对含量对光合作用有直接影响,一般呈正相关关系[15]。在本试验中,南粳3908的剑叶SPAD值与淮稻5号相当,低于南粳5055,但在净光合速率、产量上有明显优势,说明叶绿素含量不是光合速率的决定因素,光合色素的重要功能是光能转化,光系统的性能不仅与色素含量相关,还与光合反应中心的效率有关[16]。从叶绿素荧光动力学的数据可以看出,南粳3908 PSⅡ的光化学效率与南粳5055相差不大,但明显优于淮稻5号。在孕穗期各水稻品种荧光参数的日变化幅度较大,在不同时期各个水稻品种的荧光参数在12:00的差异较大,其中淮稻5号的变化幅度较大。反应中心失活会导致F0升高,而失活反应中心的增加会导致ABS/RC值、TR0/RC值、ET0/RC值升高,从结果上看,孕穗期淮稻5号在12:00的PSⅡ失活反应中心数最多,而南粳3908、南粳5055的PSⅡ反应中心相对稳定[17]。在热耗散方面,淮稻5号在12:00 PSⅡ热耗散的能量明显增加,南粳3908热耗散能量增加的幅度最小,说明南粳3908 PSⅡ具有良好的性能,能够充分利用光能、转化光能[18]。
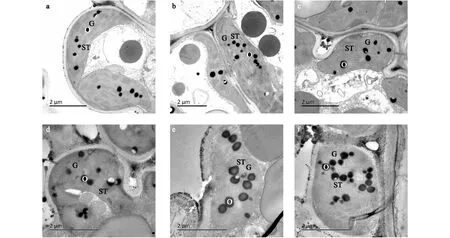
a:南粳3908 花后21 d;b:南粳5055花后21 d;c:淮稻 5号花后21 d;d:南粳 3908花后35 d;e:南粳 5055花后35 d;f:淮稻5号花后35 d。G:基粒类囊体;ST:基质类囊体;O:噬锇滴。图17 南粳3908及对照品种花后21 d、35 d剑叶叶绿体的超微结构Fig.17 Chloroplast ultrastructure in flag leaves of Nanjing3908 and the controls at 21 d and 35 d after flowering
随着叶片的衰老,各个水稻品种的Fm降低,通常最大荧光值的降低与放氧复合体失活有关,而淮稻5号在12:00的Fm较其他品种下降得多,说明其放氧复合体损伤得较多[19]。从单位截面数据来看,南粳3908、南粳5055单位面积吸收、捕获和用于电子传递的光能受光抑制的影响较小,与其他水稻品种相比,淮稻5号在12:00各参数下降得较多,说明单个反应中心的效率影响了单位面积的光能转化效率。φE0、M0、Sm通常表示PSⅡ受体侧的功能,南粳3908、南粳5055的φE0在12:00与08:00相比呈降低的趋势,但总体上高于淮稻5号,Sm的降低在一定程度上是由电子传递受阻造成的,中午强光高温导致各水稻品种的Sm降低,18:00则又升高。反应中心关闭会导致M0升高,从结果可看出,淮稻5号的M0在12:00上升得较多[17,19]。综合来看,各个水稻品种PSⅡ在中午均会发生一定程度的光抑制,反应中心存在一定程度的可逆失活,在此过程中,淮稻5号各个参数的变动幅度最大,其中孕穗期的变化幅度高于其他时期,南粳3908、南粳5055剑叶PSⅡ供体侧与反应中心、受体侧电子传递的性能优于淮稻5号,最终表现为光合性能指数(PIabs)显著高于淮稻5号。
光合蛋白是光合机构行使具体功能的重要物质,当发生光抑制时,光合蛋白也会发生不同程度的降解[20-22]。从研究结果来看,南粳3908的光合蛋白表达量高于南粳5055、淮稻5号,中午的强光高温会在一定程度上降低光合蛋白的表达量,因此午间各品种的光合蛋白表达量差异不大,各个水稻品种的光合蛋白表达都有“光合午休”现象。而南粳3908光合蛋白的表达量在18:00的恢复能力强于其他品种,这在一定程度上说明南粳3908光合蛋白的调节能力优于其他品种,能在一定程度上适应环境的变化,保持较高的光能转化能力。叶绿体功能和状态影响作物的光合作用。从叶绿体超微结构可以看出,南粳3908光合膜结构性能优于其他品种,光合膜垛叠程度高的品种的光合性能优于光合膜垛叠程度低的品种,嗜锇颗粒数目少的品种膜结构的过氧化程度较轻,其自身抗氧化能力优于其他品种,南粳3908的叶绿体衰败缓慢,基粒类囊体垛叠程度高,类囊体数目多,嗜锇颗粒数目少,这些结构特点保证了光合系统的高效运转,为光能转化提供了优良的载体,稳定的光合蛋白也能够减少光损伤和脂质过氧化[23-24]。
从本研究结果可以看出,南粳3908稳定高产的光合因素主要是光合速率高,PSⅡ供体侧、反应中心、受体侧受光合午休现象的影响较小,光合蛋白稳定,叶绿体衰败缓慢,膜脂过氧化程度轻,光合系统高效运转,能够为籽粒灌浆提供充足的光合产物。
