抚育间伐对塞罕坝地区云杉人工林碳储量及固碳速率的影响
2022-02-06张乃暄王韵頔许中旗付立华
张乃暄,王韵頔,许中旗,付立华,张 菲,程 顺
(1.河北农业大学 林学院,河北 保定 071000;2.河北省塞罕坝机械林场总场,河北 围场 068466)
森林在调控大气CO2浓度方面发挥着重要作用[1-3]。目前我国陆地碳汇强度为0.20 ~0.25 PgC/a,预计2060 年在0.15 ~0.52 Pg C/年之间[4],森林碳汇贡献了其中约80%的碳汇[5]。在我国“双碳”目标视角下,作为中国陆地碳汇的主体,森林碳汇是实现“碳中和”目标的主要途径[4]。中国森林植被的碳贮量由20 世纪 70 年代的4.38 Pg 增长到 1998 年的 4.75 Pg,平均年积累率为0.021 Pg/年[6]。不同森林类型因建群种的不同,固碳能力会有很大差别。小兴安岭地区红松(Pinus Koraiensis)林、兴安落叶松(Larix gmelinii)林、樟子松(Pinus sylvestrisvar.mongholica)林、云冷杉(Abies fabri)林、白桦(Betula platyphylla)林蒙古栎(Quercus mongolica)林和山杨(Populus davidiana)林的成熟林的碳密度分别为130.2、113.3、92.6、130.3、88.5、68.9、76.6 t/hm2[7];山西省5 种主要乔木林的固碳速率分别为落叶松[6.02 t/(hm2·年)]、山杨[10.02 t/(hm2· 年)]、 油 松(Pinus tabulaeformis)[17.23 t/(hm2·年)]、刺槐(Robinia pseudoacacia)[18.31 t/(hm2·年)]、辽东栎(Quercus wutaishansea)[20.56 t/(hm2·年)][8]。华北平原地区毛白杨(Populus tomentosa)、国槐(Sophora japonica)和松树组贡献的净碳汇量占总净碳汇量的84.41%[9]。同时,林分的固碳能力还与林分的密度直接相关[10]。甘肃地区,林分密度是影响冷杉天然林和栎类天然林碳密度最重要因素,可解释冷杉天然林碳密度差异53.00%的信息量[11]。
抚育间伐是一种常见的森林经营措施,它会通过影响林分结构而对人工林的碳汇能力产生影响[12]。轻度间伐后油松人工林乔木层总碳储量和固碳速率分别比对照组增加47.15%和52%,中度间伐后固碳速率则增加57%[13]。塞罕坝地区华北落叶松林随间伐强度增加,林木的树干、树枝、树根、树叶和树皮生物碳储量以及总生物碳储量均呈逐渐增加的趋势,38.6%为最适间伐强度[14]。南方的杉木人工林间伐5 年后则表现为乔木层碳储量减少,林下植被碳储量增多的变化特征[15]。
北半球中高纬度的森林植被是一个重要的碳汇[16],位于该地区的中国森林植被的碳汇主要来自于人工林[17]。云杉是华北地区分布较广的针叶树种之一,主要在海拔和纬度相对较高的地带形成纯林,也是塞罕坝地区的主要造林树种之一。研究抚育间伐下云杉人工林的固碳速率变化趋势,对于提高云杉人工林的固碳增汇能力具有重要意义。本研究以塞罕坝地区的云杉林为研究对象,研究抚育间伐强度对云杉人工林碳储量及固碳速率的影响,为该地区云杉林的合理经营,和碳储量以及碳汇作用的评估提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于河北省塞罕坝机械林场(41°22′~42°58′N,116°53′ ~118°31′)所 属 的 千 层 板 林 场和北漫甸林场。塞罕坝地区位于冀北山区及内蒙古高原的交汇地带,内蒙古高原南缘,与内蒙古克什克腾旗和多伦县部分接壤,坝上地区平均海拔高度1 500 ~1 940 m。该区冬季漫长而寒冷,每年积雪时间长达7 个月,春秋两季短暂且干燥,年平均气温-1.2℃,昼夜温差大,平均日照时数为2 546 h。该地区年均降水量460 mm,多风沙天气,年平均大风天气80 d 左右。该地区植被以人工林为主,包括华北落叶松(Larix principis-rupprechtii)林、云杉(Piceameyeri)林、樟子松(Pinus sylvestrisvar.mongolica)林等,还有天然次生白桦(Betulaplatyphylla)林和蒙古栎(Quercus mongolica)林等。
1.2 样地设置及调查
于2018 年8 月,在对塞罕坝机械林场的云杉人工林进行全面踏查的基础上,在千层板林场和北漫甸林场选择不同抚育强度的云杉林设置面积为20 m×30 m的调查样地11 块,其中未抚育间伐3 块,高强度抚育间伐5 块,低强度抚育间伐3 块,共11 块样地。间伐林分初始密度在3 000 ~3 300/hm2株之间,分别在20 年生、25 年生和30 年生时进行过抚育间伐,每次间伐强度分别30%(高强度)和15%(低强度),均为株树采伐强度。对各样地进行每木检尺,调查内容包括坐标、坡向、坡度、胸径、树高、冠幅等。胸径测量采用胸径尺、冠幅采用激光测距仪、树高采用勃鲁莱测高器进行测定。各林分概况见表1。
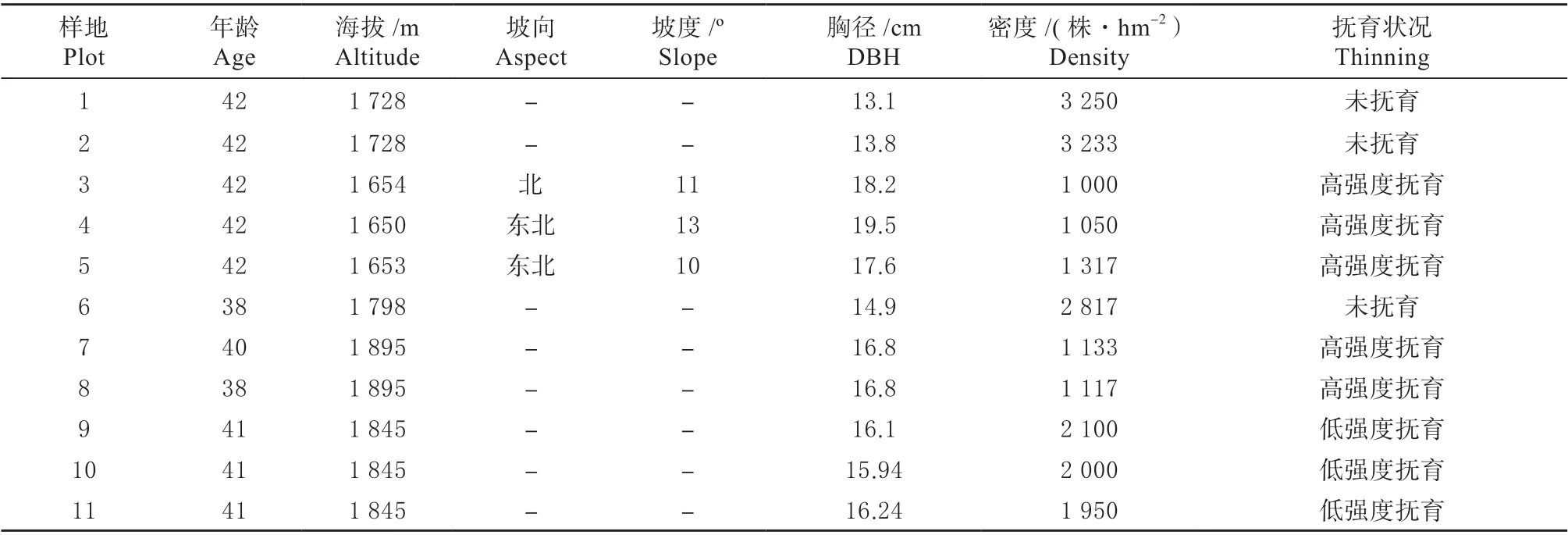
表1 各样地概况Table 1 Conditions of the plots
1.3 生物量和生物碳储量测定
根据样地调查结果,在调查样地中选取不同径级林木进行树干解析和地上部分生物量的测定,共30 株。生物量按照枝、干、叶分别进行调查。其中,枝和叶生物量采用标准枝法进行测定,树干生物量采用全部称重法进行测定。同时,从侧枝、叶片和主干中,取一部分样品,采用烘干法测定含水率,由各部分的鲜重及含水率换算得到各部分生物量。根系生物量根据IPCC 给出的根冠比进行计算,根冠比取0.39。生物碳储量依照国内外惯例,取50%的含碳率计算乔木生物碳储量[18]。
1.4 单株地上生物量模型
本研究以解析样木的实测数据为基础,使用目前国际上比较公认的两种生物量模型进行拟合,建立云杉的单株地上生物量方程[19],并计算林分地上部分总生物量。模型形式分别为:

式中:W 为立木地上生物量;D、H分别为测树因子胸径、树高;a、b 为模型参数。
1.5 林分固碳速率的计算
1.5.1 标准木固碳速率的计算 固碳速率的计算一般采用“时间序列法”[20]。由各林分标准木的树干解析数据得到其各年龄的树高及胸径总生长量,将各年龄的树高和胸径带入云杉生物量模型(2 式),得到各年龄标准木的生物碳储量,由标准木相邻两年的生物碳储量之差得到当年的固碳速率:

Pt为林分标准木的t年的生物碳储量,Wt为t年的标准木的单株生物碳储量。
1.5.2 云杉林固碳速率计算 由林分标准木的固碳速率乘以林分密度得到林分固碳速率:

其中,Pft为t年的林分固碳速率,Pt为林分标准木的固碳速率,N为林分密度。
2 结果与分析
2.1 云杉生物量模型
两种地上生物量及各器官生物量的回归模型的拟合度都处于较高水平,决定系数均在0.882 0 以上,最高达0.986 4,均达到显著水平(表2)。总体来看,相对于W=aDb模型,W=a(D2H)b生物量模型的误差平方和(SSE)、均方误差(MSE)和均方根误差(RMSE)均较低,且确定系数(R2)更高,因此,W=a(D2H)b生物量模型的拟合效果更好。云杉地上生物量实测值与模拟值之间具有较好的相关性和一致性(图1)。另取3 株云杉生物量的实测数据对该生物量模型进行检验,检验结果显示模拟值与实测值没有显著差异(P=0.11),说明该模型拟合效果较好,可用来对云杉的生物量进行估算。
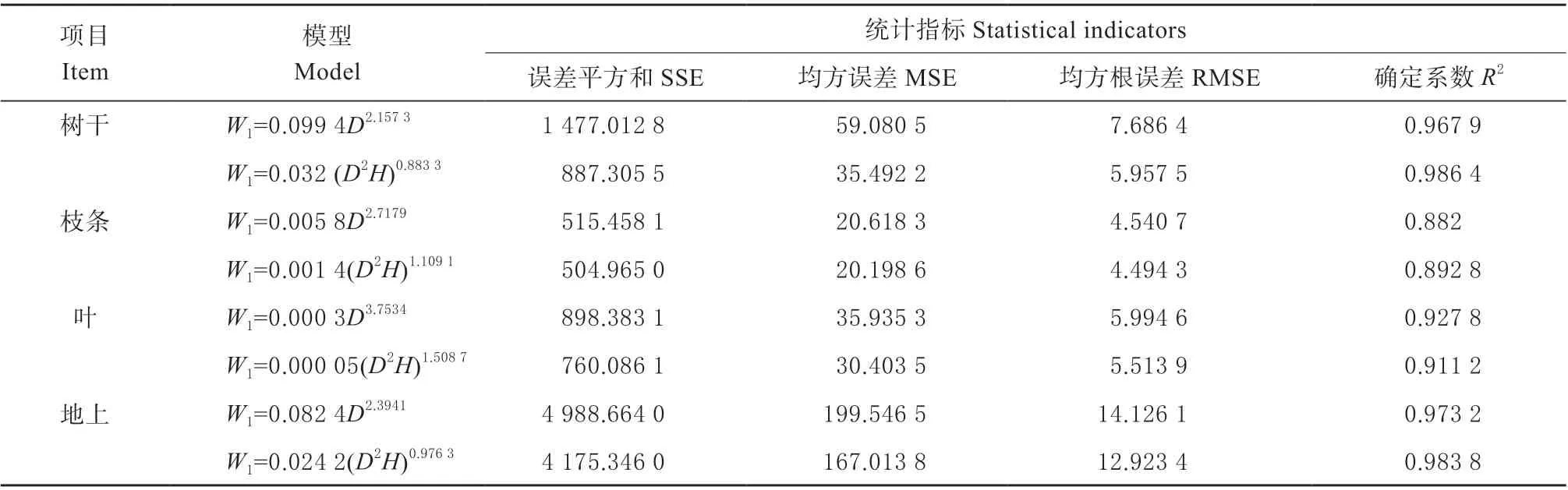
表2 单株及各器官生物量模型及统计指标Table 2 Biomass regression models of individual plants and organs
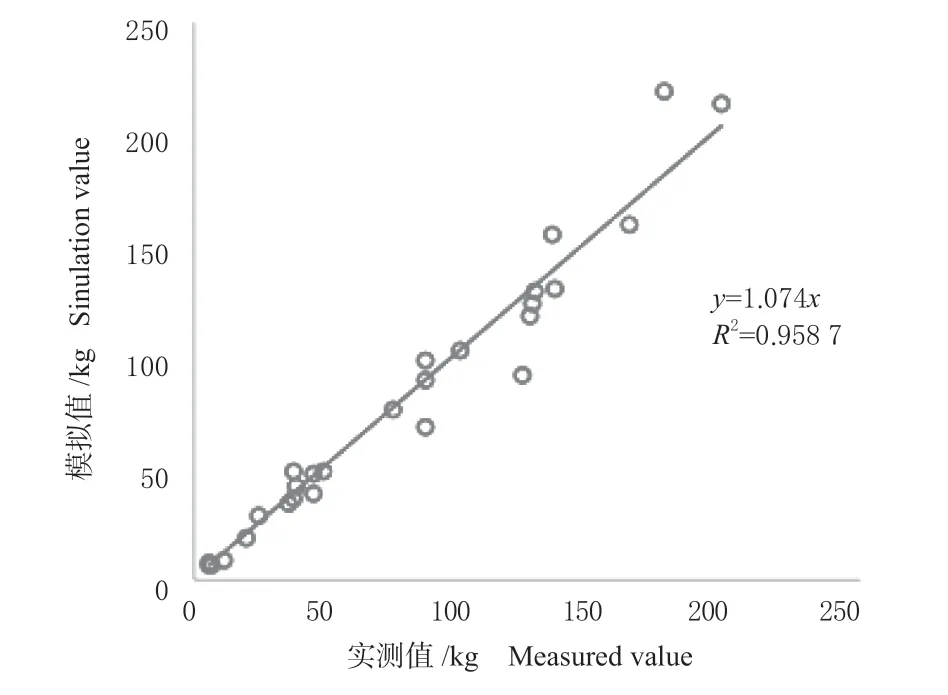
图1 地上生物量拟合值与实测值的关系Fig. 1 The relationship between the simulation values and measured values of aboveground biomass
2.2 不同抚育强度云杉林单株生物碳贮量及其分配特征
为消除年龄因素对不同抚育强度生物碳储量研究结果的影响,各样地均按照树干解析表以及生物量方程计算得出38 年生云杉单木的生物量,进而获得生物碳储量来进行比较。
由表3 可见,不同抚育强度下,平均木和优势木的各组分生物碳储量以及地上总生物碳储量均表现为随抚育强度的增大而增加的趋势。其中,高强度抚育林分单株地上生物碳储量分别是未抚育和低强度抚育的2.27 倍和1.68 倍,其中树干生物碳储量分别是1.84 倍和1.40 倍,枝生物碳储量分别是3.13倍和2.13 倍,叶生物碳储量分别是3.94 倍和2.70 倍。另外,不同抚育强度下优势木的单株地上碳储量同平均木表现相同,皆为高强度抚育后的云杉林单株碳储量最大,低强度次之,未抚育的林分最低。因此,抚育采伐对云杉林单株碳储量具有明显影响,各器官生物碳储量均随抚育强度的增加而增加,同时,枝和叶生物碳储量对抚育采伐的响应更为明显。
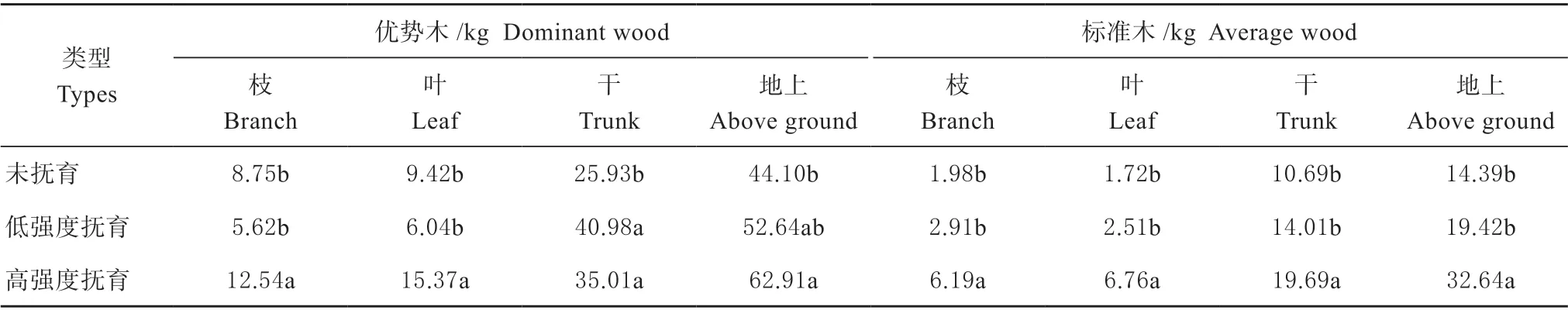
表3 不同抚育强度云杉单株生物碳储量Table 3 Vegetation carbon storage of spruce individual tree under different thinning intensities
云杉不同器官的生物碳储量均表现为干所占比重最大,占地上总生物碳储量的59.8%~74.3%;枝和叶生物碳储量所占比重较小, 分别为13.74% ~19.08% 和11.93% ~21.12%( 图2)。在不同抚育强度下,各器官生物碳储量的分配比例存在一定差异。枝、叶生物碳储量的百分比均随抚育强度的增大而增加,而主干生物碳储量则相反,未抚育、低强度抚育和高强度抚育云杉林主干生物碳储量百分比分别为74.3%、71.60% 和59.8%。云杉优势木的各器官生物碳储量在未抚育和低强度抚育时为干>叶>枝;云杉标准木的各器官生物碳储量在未抚育和低强度抚育时为干>枝>叶,而高强度抚育下则表现为与优势木相同的干>叶>枝。由此可知,抚育采伐不仅会改变云杉林单株林木的生物碳储量,还会对生物碳储量的分配产生明显影响。
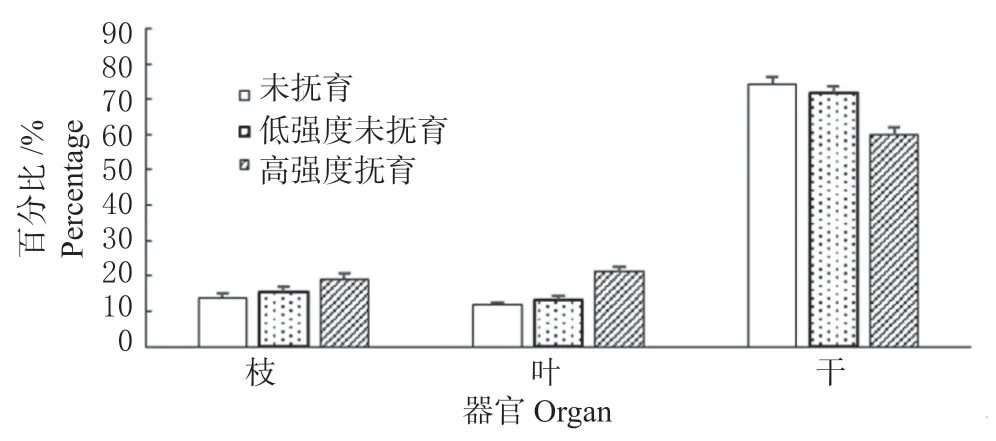
图2 各器官生物碳储量占地上总生物碳储量百分比Fig. 2 Percentage of vegetation carbon storage of different organ in the total above ground biological carbon storage
2.3 不同抚育强度云杉林生物量及其碳贮量的比较
为了消除年龄差异带来的影响,不同抚育强度云杉林均取38 年生林分进行比较(表4)。由表4 可知,林分总生物量及各器官生物量均随抚育强度的增加而逐渐降低,其总生物量分别为123.98、108.62 和100.77 t/hm2,高强度抚育林分总生物量分别比未抚育和低强度抚育林分低18.73% 和12.40%,树干生物量低34.05%和15.87%。
但是,枝、叶生物量与总生物量的变化趋势不同,表现为:高强度抚育>未抚育>低强度抚育,其中枝生物量分别为13.63、12.09 和11.93 t/hm2,叶生物量则分别为15.01、10.60 和10.26 t/hm2,高强度抚育林分的枝生物量分别比未抚育和低强度抚育林分的高12.69%和14.22%,叶生物量分别高41.59%和46.29%。
随着抚育强度的增加,林分碳储量呈现下降的趋势。如表4 所示,云杉林分碳储量变化趋势为未抚育>低强度抚育>高强度抚育,其林分碳储量分别为61.99、54.31 和50.39 t/hm2。相比于未抚育的对照组,低强度抚育的林分碳储量降低了7.68 t/hm2,高强度抚育的林分碳储量降低11.60 t/hm2。未抚育和低强度抚育林分的碳储量分别比高强度抚育林分高23.02%和7.78%。
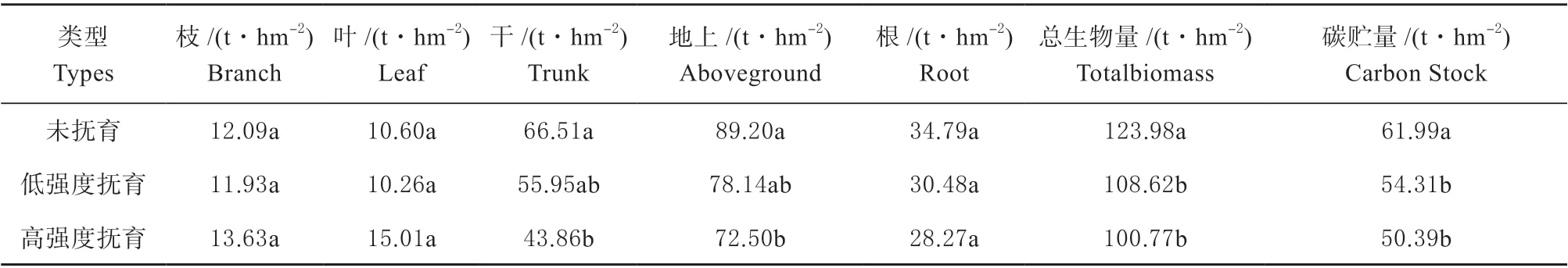
表4 不同抚育强度云杉林生物量及碳储量Table 4 The biomass and vegetation carbon storage of the spruce forests under different thinning intensities
2.4 不同抚育强度云杉林的固碳速率的比较
云杉单株林木固碳速率,在34 年之前呈逐渐上升的趋势,在34 年达到最高值,之后虽有波动,总体呈逐渐下降的趋势(图3)。同时,抚育对云杉单木固碳速率具有明显影响,单木固碳速率总体表现为未抚育<低强度抚育<高强度抚育,即随着抚育强度的增加,单木固碳速率呈逐渐增加的趋势。以34 年为例,未抚育、低强度抚育和高强度抚育的单株固碳速率分别为1.45、1.80 和3.28 kg/年,抚育为未抚育的2.26 倍。同时,从图3 可以看出,固碳速率的差异随年龄的增加而逐渐增大。在15 年生之前,各林分都未进行抚育间伐,因此单木固碳速率没有明显差异,抚育间伐之后,其效应逐渐显现,各林分之间的差异逐渐增大。
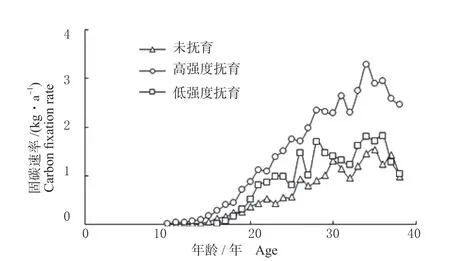
图3 不同抚育强度云杉单株固碳速率Fig. 3 The carbon fixation rate of individual spruce tree under different tending intensities
不同抚育强度云杉林林分固碳速率与单株不同,以38 年生为例,云杉林固碳速率以未抚育林分最高,为4 058.83 kg/(hm2·年),其次为高强度抚育,为3 173.26 kg/(hm2·年),低强度抚育为最低,为2 908.00 kg/(hm2·年),但高强度抚育与低强度抚育之间没有明显差异(P>0.05)(图4)。
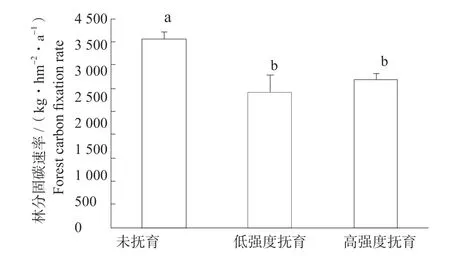
图4 不同抚育强度云杉林固碳速率Fig. 4 The carbon fixation rate of the spruce forests under different thinning intensities
3 讨论
森林生物量是反映森林生态系统固定CO2能力的重要指标。本研究建立了塞罕坝地区云杉地上生物量模型,从均方误差(MSE)和均方根误差(RMSE)及决定系数(R2)综合比较来看,以W=a(D2H)b模型的拟合效果较好,与多数生物量估算模型相同[21-22]。
抚育采伐对云杉的生物碳储量有明显影响。抚育采伐明显促进了单株生物碳储量的增长,且抚育强度越大,增长量越大。这一结果与当前大部分研究结果一致[13,15]。不同间伐梯度对单株生物碳储量都具有促进作用,抚育间伐降低了林分密度,保留木的生长空间扩大,因此单株生物碳储量明显增加。但抚育强度不同,其对碳储量的影响也不同。北京地区油松人工林,轻度间伐对单株生物碳储量的促进最大,而后随着间伐强度的增加碳储量减少[13]。
同时,抚育采伐降低了云杉林分生物碳储量,高强度抚育间伐和低强度抚育间伐林分生物碳储量分别为未抚育间伐的81.28%和88.07%。这与丹麦北部云杉林碳储量的研究结果相似[23],皆表现出云杉林的林分碳储量随着间伐强度的增加而减少的趋势。间伐林分生物碳储量低于未间伐林分表明间伐导致所引起的密度下降效应大于伐林对单株生物碳储量的促进效应。间伐后调查的间隔期时间会对研究结果造成影响,随着林分继续生长以及对单株生物碳储量单株增长效应的持续发挥,不同抚育间伐强度林分生物碳储量会逐渐趋同[24],抚育间伐对碳储量的促进作用会逐步增强[13,25]。
间伐不但对云杉生物碳储量具有重要影响,还对其生物碳储量分配具有重要影响。抚育间伐使枝叶生物碳储量所占比例明显增加,且有随抚育强度的增大而增加的趋势。未间伐林分的郁闭度大,导致云杉的自然整枝强烈,下层枝条大量死亡,使得枝叶生物量的比重较低。间伐后林分郁闭度下降,保留木能够获得充足的光照条件,一方面促进了侧枝的发育,另一方面也使得下部枝条的自然整枝得到缓解,使得枝叶的生物量增加。
抚育采伐导致云杉单株林木固碳速率明显增加,且总体上,抚育强度越大,单株林木的固碳速率越高。未抚育的云杉单木固碳速率与北京地区油松单木相近,而高强度抚育下云杉单木固碳速率要高于油松[13]。林分固碳速率则以未抚育林分为最高,高强度抚育略高于低强度抚育,但未表现出明显差异。与低强度抚育相比,高强度抚育下的云杉林的枝叶生物量更高,因为叶片是光合器官,所以高强度抚育下的云杉林的林分密度尽管低于低强度抚育云杉林,但林分固碳速率还略高于低强度抚育林分[15]。以上结果表明,抚育强度的增加,并不一定导致林分固碳速率的下降,这与多数研究结果一致,合理的抚育间伐能够提高林木单位固碳潜力[25]。在一定抚育强度下,保留木生长的资源空间得到明显扩大,枝叶生物量得到明显增加,从而使林分固碳速率能够达到一个较高水平[26]。本研究中两种抚育强度林分的固碳速率低于未抚育林分,其原因可能在于间伐后时间还相对较短,间伐后的正效应尚未完全发挥。另外,本研究中只有两种间伐强度,尚不能确定何种强度对于云杉林的固碳速率提高最为合理,需要进一步的研究来进行确定。
4 结论
(1)建立了塞罕坝地区云杉地上生物量模型,从均方误差和均方根误差及决定系数综合比较来看,以W=a(D2H)b模型的拟合效果较好。
(2)抚育间伐对云杉生物碳储量及其分配有明显影响,抚育间伐导致云杉单株生物碳储量明显增加,但林分生物碳储量减少;同时间伐导致云杉枝叶生物碳储量比例明显增加,而干生物碳储量所占比例明显下降。
(3)抚育采伐促进了云杉单株固碳速率的明显提高,同时使得林分固碳速率有所下降,高强度抚育间伐林分的固碳速率略高于低强度抚育间伐,但没有明显差异。
