陆地棉泛素结合酶基因GhUBC4 调控开花时间机制探究
2022-02-06张子豪李会敏张竞予阎媛媛马峙英
张子豪,李会敏,张竞予,阎媛媛,马峙英
(河北农业大学 华北作物改良与调控国家重点实验室/河北省作物种质资源重点实验室/农学院,河北 保定 071000)
棉花被认为是研究多倍体植物的模式物种,为了解植物基因和基因组的进化模式和机制提供了组织框架和系统发育的视角[1]。棉花异源四倍体栽培品种通过亚洲棉Gossypium arboreum(Ga, A2)和雷蒙德氏棉Gossypium raimondii(Gr, D5)2 个二倍体物种的种间基因组融合和加倍形成5 个异源四倍体种[2],其中陆地棉(Gossypium hirsutumL.)是目前种植最广泛的棉种。
泛素- 蛋白酶体途径(Ubiquitin-proteasome pathway)在调节和维持细胞内稳态中起着重要作用,选择性降解异常蛋白和短命调节蛋白,包括细胞周期蛋白、信号转导器、细胞生长调节剂和转录因子[3]。泛素结合酶E2(Ubiquitin-conjugating enzyme E2)是泛素-蛋白酶体系统中关键组成部分之一,具有特殊的意义和功能[4]。拟南芥AtUBC2响应脱水、高盐和脱落酸(ABA)处理,正向调节渗透胁迫的耐 受 性[5],绿 豆VrUBC1[6]、大 豆GmUBC9[7]和CmUBC[8]、花生AhUBC2[9]具有相同的功能。AtUBC32、AtUBC33和AtUBC34可以与干旱胁迫负调控因子PUB19相互作用,通过脱落酸的介导在干旱胁迫中发挥负调控作用[10],并且抑制种子萌发和初生根生长[11]。此外,AtUBC18通过改变AtERF1的丰度和下游基因的表达,负调控干旱和盐胁迫的响应[12]。橡胶树Rad6和水稻OsRad6与拟南芥中AtUBC2的相似性超过96%,其表达可被乙烯(ET)和茉莉酸甲酯(MeJA)显著诱导[13]。AtRCE1与人UBC12基因同源,其与泛素类蛋白RUB1 结合,影响SCF E3s 的功能,从而影响生长素应答[14]。AtUBC24(pho2) 受microRNA399(miR399) 调控,控制无机磷酸盐(Pi)的稳态以及Pi 的易位和再活化[15]。
本实验室前期通过全基因组关联分析在陆地棉D03 染色体上挖掘到一个与棉花开花时间相关的候选基因GhUBC4[16],本研究对该基因进行了克隆与功能分析,由于不同陆地棉品种中单核苷酸多态性(SNP)位点的改变,GhUBC4具有不同的序列,将其命名为GhUBC4Ref和GhUBC4Alt,以期解析GhUBC14位点变异促进棉花开花的机制,为早熟棉育种提供理论和分子基础。
1 材料与方法
1.1 生物信息学分析
在线网站SWISS-MODEL(https://swissmodel.expasy.org/interactive/Vh8qXc/models/) 预 测 蛋 白三级结构;MEGA7.0 进行系统进化树分析;在线网 站CDD Search(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)用于保守结构域分析;在线网站Plant CARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)用于启动子顺势作用元件分析;NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)和DNAMAN 软件对氨基酸序列进行比对。
1.2 植物培养及取样方法
拟南芥种植于光照培养室(23 ℃,16 h 光照/8 h 黑暗),在子叶展平后7 d 进行取样,用于基因表达分析。陆地棉TM-1 种植于河北农业大学温室(28 ℃,12 h 光照/ 8 h 黑暗),两片真叶期开始取样,到五片真叶期结束,每次选取长势均匀一致的棉株叶片及顶端分生组织用于基因的实时定量表达分析及相关基因克隆。所取材料立即投入液氮中冷冻,于-80 ℃冰箱保存备用。
1.3 基因克隆及拟南芥的遗传转化
以‘Babshaw1’和‘中棉所16 号’的叶片cDNA为模板扩增GhUBC4Ref和GhUBC4Alt的CDS 序列。扩增产物利用双酶切法构建植物超表达35S:pGreen载体。利用农杆菌浸染转化法进行拟南芥的遗传转化。本研究中使用的引物如表1 所示。

表1 本研究中所用引物序列Table 1 Primer sequences used in this study
1.4 亚细胞定位
构建目的基因与GFP 融合表达载体35S:GhUBC4Ref-GFP 和35S:GhUBC4Alt-GFP,采 用 电击法将重组质粒转化农杆菌感受态GV3101,28℃震荡培养。当烟草(Nicotiana bethamiana)长到五片真叶时,将农杆菌培养至OD600=1.0 ~1.2,离心收集菌液,将菌体重悬于缓冲液(10 mmol/L MES,10 mmol/L MgCl2, 100 mmol/L AS),使用1 mL 无针头注射器,将农杆菌注射到烟草叶片背面。侵染后黑暗培养48 h,利用DAPI 进行染色,使用智能激光共聚焦显微镜(FV1000)观察目标蛋白表达位置。
1.5 病毒诱导基因沉默
扩增GhUBC4目的基因的特异性片段,并连接到pTRV2 载体中。当棉花植株2 片子叶展平时,将含有目的基因的pTRV2 农杆菌与pTRV1 农杆菌等体积混合后注射早熟棉花品种‘CCRI50’叶片背部。pTRV2:CLA1和pTRV1 分别用作阳性对照和阴性对照。当阳性对照出现白化表型时,收获叶片以评估基因的沉默效率,检测开花促进子GhFT、GhSOC1、GhLFY表达量,之后统计第一果枝节位、现蕾天数和开花时间。
1.6 RT-PCR
使用RNA prep Pure Plant Kit(天根,北京,中国)提取总RNA。使用PrimeScriptTM1st strand cDNA Synthesis kit(索莱宝,美国)进行反转录。RT-PCR反应体系为10 μL,其中包括前后引物各0.5 μL,5 μL 2×AugeGreen qPCR Master Mix (US Everbright),1 μL 模板cDNA,2.9 μL ddH2O,0.1 μL ROX。反应条件:95 ℃ 2 min;95 ℃ 5 s,60 ℃ 5 s,40 个循环;72 ℃ 50 s。棉花以GhUBQ14基因为内参,拟南芥以AtTUB2基因为内参,采用2-△CT法计算相对表达量。每个样品进行3 个生物学重复和3 个技术重复。
2 结果与分析
2.1 开花相关基因的鉴定
本课题组前期对419 份陆地棉核心种质进行了重测序,基于366 万个SNPs 的全基因组关联分析发现与开花天数相关的SNPs(1 199 个)位于强信号的D03 染色体上,在25.21 ~25.22 Mb 的位点发现1 个UBC 基因与开花时间显著关联。该基因在早熟棉品种中基因型为GGA,而在晚熟棉品种中基因型为TTT,与同样晚熟的陆地棉参考基因组TM-1基因型相同。
2.2 GhUBC4 基因克隆及生物信息学分析
为了分别克隆2 种基因型,以‘Babshaw1’和‘中棉所16 号’的叶片cDNA 为模板进行扩增,得到450 bp 左右的特异性条带,测序结果显示2 序列氨基酸序列相似度高达99%,仅有1 个氨基酸发生改变(图1A),分别命名为GhUBC4Ref和GhUBC4Alt。C 端氨基酸序列的差异导致2 个蛋白的结构明显不同(图1B)。GhUBC4不同基因型并不影响基因结构,均包含5 个外显子和4 个内含子。在NCBI 中对GhUBC4 蛋白进行氨基酸多序列比对,发现GhUBC4 蛋白与多个物种均有较高的相似性,其生物功能存在一定的保守性。系统进化树分析显示,GhUBC4 蛋白与锦葵科棉属的雷蒙德氏棉、海岛棉、亚洲棉以及锦葵科木槿、榴莲的亲缘关系最近,说明这些物种的GhUBC4蛋白进化于共同祖先(图1C)。利 用Plant CARE 对GhUBC4上 游3 000 bp 序列进行顺式作用元件预测(图1D)。结果显示,GhUBC4除了含有TATA-box 和CAAT 核心启动子元件外,还检测到多种光反应顺式作用元件AT1-motif、GTGGC-motif、GATA-motif、G-box、AE-box、GT1-motif、Box-4、TCT-motif,茉莉酸响应元件CGTCA-motif、TGACG-motif,脱落酸响应元件ABRE,赤霉素响应元件GARE-motif、TATCbox,低温响应的顺式作用元件LTR 及厌氧诱导元件ARE,分生组织表达相关的顺式作用调控元件CAT-box,及参与昼夜节律控制的顺式作用调节元件Circadian,暗示该基因受多种信号通路调控。

图1 陆地棉GhUBC4 基因克隆及生物信息学分析Fig. 1 Cloning and bioinformatics analysis of GhUBC4 in G. hirsutum
2.3 GhUBC4 基因表达模式分析
利用已发表的南京农业大学转录组数据库[17]对GhUBC4表达特征进行分析,发现该基因在棉花的各个组织中均表达,且表达量不具有组织特异性(图2A)。为了探究GhUBC4是否参与棉花的生长发育过程,检测该基因在陆地棉TM-1 二叶期到四叶期的顶端分生组织和叶片中的表达变化(图2B)。结果显示,随着棉苗的生长发育,GhUBC4基因的表达量在叶片Leaf 和顶端分生组织SAM 中上调表达,该基因表达模式与开花促进子一致。同时,选取了10 个早熟棉品种和10 个晚熟棉品种,在四叶期进行取样,检测GhUBC4基因的表达量(图2C),发现该基因在早熟棉花品种中的表达量普遍高于晚熟品种,表明该基因参与棉花开花调控。
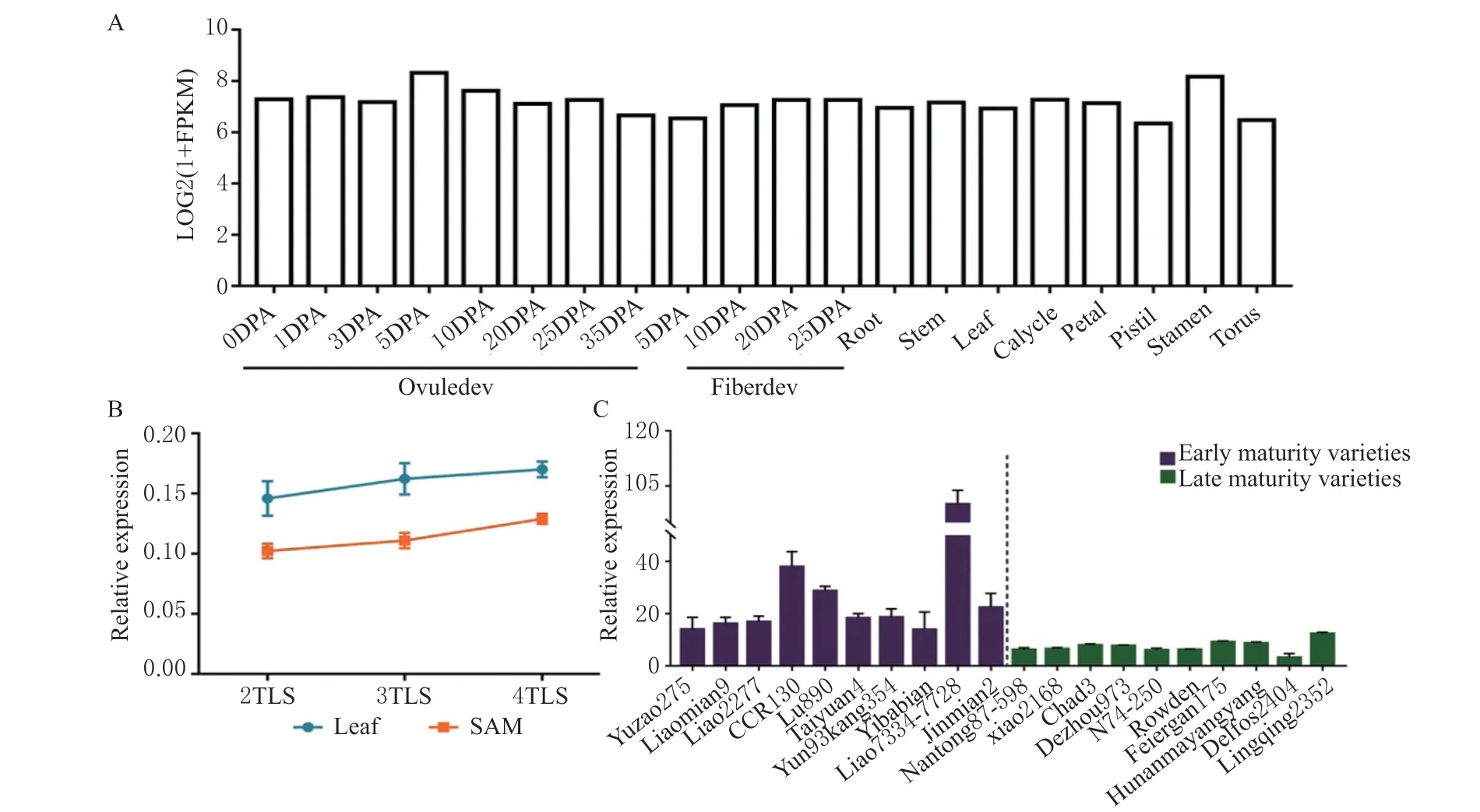
图2 陆地棉GhUBC4 基因表达模式分析Fig. 2 Expression analysis of GhUBC4 in G. hirsutum
2.4 GhUBC4 亚细胞定位
为了确定GhUBC4 在细胞中行驶功能的位置,我们将不含终止密码子的编码序列与GFP 载体N端融合,并在烟草表皮细胞中瞬时表达,结果显示GhUBC4Ref-GFP 和GhUBC4Alt-GFP 两 种 基 因 型的融合蛋白绿色荧光信号均定位在细胞核中,表明GhUBC4 在细胞核中行使功能。
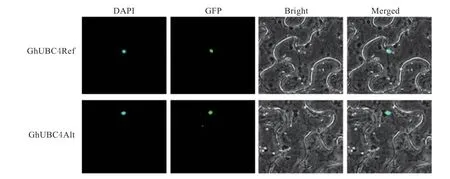
图3 GhUBC4 亚细胞定位Fig. 3 Subcellular localization of GhUBC4 in tobacco leaves
2.5 沉默GhUBC4 抑制开花
利用病毒诱导基因沉默技术在早熟棉‘CCRI50’中沉默GhUBC4,结果显示当沉默效率在80%以上时,沉默植株的果枝始节位、现蕾天数、开花天数显著高于对照植株,沉默植株第一果枝位于第9 节,现蕾时间延迟约6 d,且对照植株开花时,沉默植株不开花(图4)。表明GhUBC4表达的降低延迟了陆地棉开花时间,该基因具有促进棉花开花的功能。
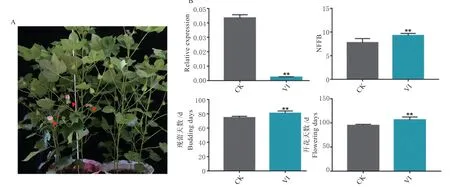
图4 沉默GhUBC4 抑制开花Fig. 4 Silencing GhUBC4 inhibits flowering
2.6 超表达GhUBC4 促进拟南芥早花
为了探究不同基因型GhUBC4调控开花时间的功能,构建了超表达载体35S:GhUBC4Ref和35S:GhUBC4Alt,并将其转化到拟南芥中。在长日照条件下,超表达GhUBC4Ref和GhUBC4Alt均促进拟南芥开花(图5A),荧光定量PCR 检测发现GhUBC4Ref在转基因植株中表达丰富,且表达量越高,植株开花越早。但GhUBC4Alt的表达利用qPCR 检测CT 值较大,表明其在转基因拟南芥中表达较少,无法被qPCR 方法准确检测,因此我们用半定量法对其mRNA 进行扩增,发现转基因植株能完整的转录GhUBC4Alt,且转录相对较多的#1和#2 株系开花时间早于株系#3(图5B, 5C)。表明两种GhUBC4基因型均具有促进开花的功能,且GhUBC4促进开花功能具有剂量效应。
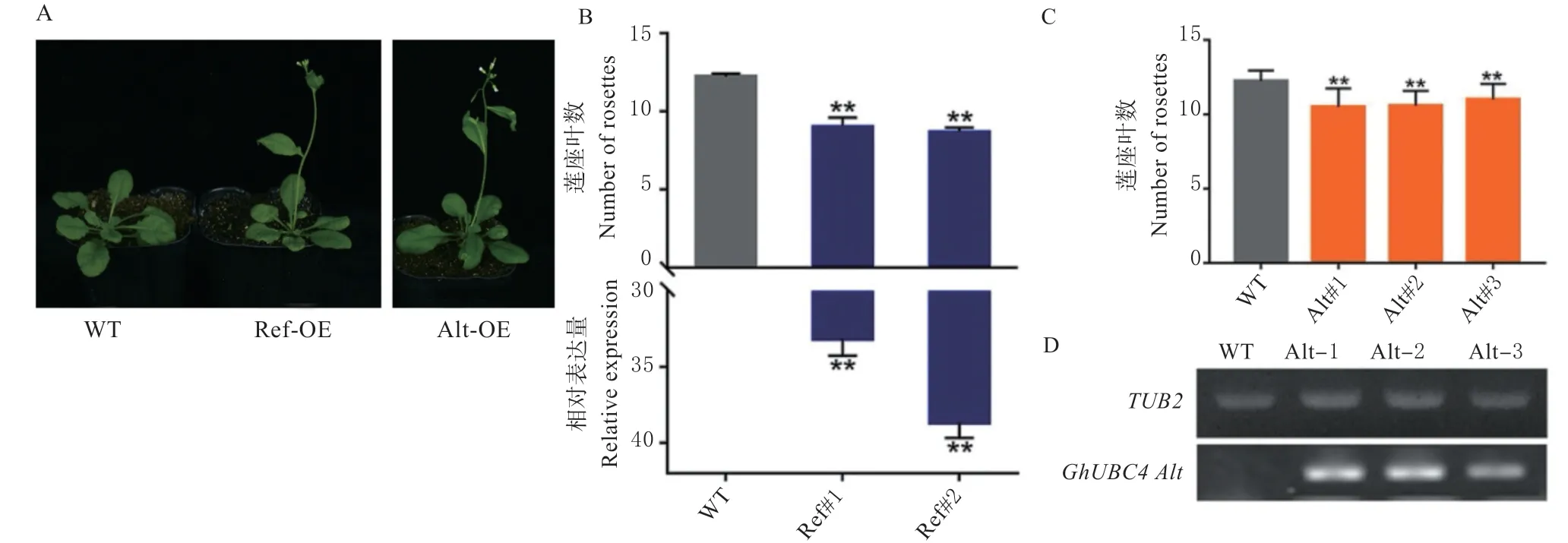
图5 超表达GhUBC4 促进拟南芥开花Fig. 5 Overexpression of GhUBC4 promotes flowering in Arabidopsis thaliana
2.7 GhUBC4 与开花促进子作用关系初步探究
在复杂的开花调控网络中,开花促进子FT、SOC1、LFY整合环境和内源的各种信号,最终激活花芽分化,开启生殖生长。因此我们进一步检测了转基因拟南芥和沉默植株中3 个重要开花促进子的表达。与野生型拟南芥相比,35S:GhUBC4Ref和35S:GhUBC4Alt拟 南 芥 中AtFT、AtSOC1、AtLFY的表达量均显著增加(图6A)。并且在35S:GhUBC4Alt植株中,开花促进子的转录水平均高于35S:GhUBC4Ref,而GhUBC4Alt的转录丰度明显低于GhUBC4Ref,说明GhUBC4Alt对开花促进子的促进功能更强。在早熟棉品种‘CCRI50’中沉默GhUBC4后,开花整合子GhFT、GhSOC1、GhLFY的表达量呈显著下调表达(图6B)。这些结果表明GhUBC4正调控开花整合子从而促进拟南芥和棉花早花。
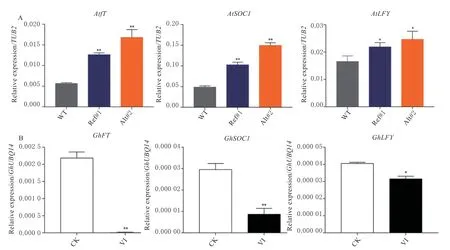
图6 GhUBC4 与开花促进子作用关系初步探究Fig. 6 Preliminary study on the relationship between GhUBC4 and flowering time integrons
3 讨论
水稻CsUBC24与花发育调控因子CsSEPs 和花药开裂调控因子CsICE1 互作,通过下调CsSEPs和CsICE1破坏花分生组织调节网络,导致花的发育异常及雄性不育[18]。番茄E2 基因LeUBC的转录水平在衰老叶组织和发育中的果实中增加,并在果实成熟开始时达到峰值[19]。GhUBC1/2选择性降解叶片和花衰老过程的蛋白[3]。本研究发现1 个调控开花时间的泛素化结合酶基因GhUBC4,该基因编码区的变异与棉花熟性相关,且在早熟品种中普遍高表达,这与其促进拟南芥早花的表型一致,说明该基因具有促进开花的功能。
一般认为,植物开花调控网络包括光周期途径、温度途径、春化途径、赤霉素途径和年龄途径。目前对UBC基因参与开花调控机制的研究均为其对FLC及其同源基因的调控。与野生型拟南芥相比,hub1和hub2单突变体,以及ubc1和ubc2双突变体中FLC 的表达显著降低,表明FLC位点组蛋白H2B 的单泛素化和去泛素化是FLC适当表达所必需的[20-21]。此外,在低温条件下,转录因子WRKY34结合到E3 连接酶复合体组分CUL3A的启动子上增加其转录,使FRIGIDA(FRI)降解,从而抑制FLC的表达[22]。GmUBC9正调控FLC基因导致转基因拟南芥晚花[7]。这些结果均表明UBC基因抑制开花时间。与之相似,对E3 基因参与开花时间调控的研究也表明泛素化抑制开花时间。RING 蛋白FRRP1 是HUB2 在水稻中的同源基因,FRRP1 通过泛素化组蛋白H2B 抑制Hd3a 的表达[23]。拟南芥光周期路径CO 蛋白的丰度受泛素化的周期调控,进而调控FT 的表达[24]。SCF E3s 连接酶复合物成员FKF1 蓝光下与COP1 互作,阻止其泛素化CO[25]。水稻环型E3 连接酶HAF1 通过26S 蛋白酶复合体降解OsELF3,从而调节OsELF3 蛋白昼夜节律性累积[26]。然而,GhUBC4促进开花,这拓宽了我们对泛素化功能的认识。与此类似,磷脂乙醇胺结合 基 因(Phosphatidyl ethanolamine-binding gene)家族中TFL1抑制AP1和LFY基因的表达[27],而陆地棉GhTFL1a基因作为TFL1的同源序列,其功能与TFL1相反,并参与了光周期路径[28]。突变型GhUBC4蛋白结构在C 端发生变化,可能影响其与蛋白的互作。功能分析表明,GhUBCAlt尽管在拟南芥中表达水平显著低于GhUBCRef,但其对开花促进子的促进作用更强,说明不仅在转录水平上,早熟棉中GhUBC4Alt大量积累,其蛋白的功能也相对增强。
多个棉花基因组的测序与组装拓宽了我们在基因组水平对棉花多倍体进化事件、变异与适应的认识,同时推进棉花研究进入后基因组时代。与模式植物不同,在棉花不存在春化途径,因此基因组中也缺失FLC基因[29],陆地棉UBC基因调控开花时间的机制不同。并且在棉花中,开花促进子GhFT、GhLFY在A、D 亚组各存在一个拷贝,然而SOC1和AP1同源序列较多,其功能的冗余性和多样性增加了棉花开花调控网络的复杂性。本研究发现超表达GhUBC4显著促进SOC1和FT的转录,而LFY表达量的升高不如FT和SOC1。表明GhUBC4可能通过降解调控FT和SOC1上游共同的调控因子进而调控开花。受限于棉花开花调控网络研究,目前对开花抑制子的认识尚浅,制约着我们对该基因调控机制的解析。挖掘其互作的蛋白及调控的靶基因对深入了解GhUBC4基因如何调控开花时间具有重要意义,也是今后研究的重点。
