柠条种子萌发对不同温度和土壤含水量的响应
2022-02-03闫兴富
杨 慧, 张 泽, 张 兰, 闫兴富
(北方民族大学生物科学与工程学院/国家民委黄河流域农牧交错区生态保护重点实验室,宁夏 银川 750021)
种子萌发策略是干旱荒漠地区植物生活史对策的重要方面,对植物的种群更新及植被恢复均具有重要意义。植物在对干旱荒漠地区生境的长期适应过程中,为了保证适宜种子萌发和幼苗生长的时空条件,进化出一些特殊萌发策略[1]。植物为了逃避不利于幼苗生长与存活的恶劣环境条件,种子可在适宜条件下萌发建植幼苗,而在恶劣环境条件下保持休眠状态得以分散萌发风险[2-3]。温度和水分是影响植物种子萌发的重要环境因子,温度可直接影响种子萌发或通过改变种子休眠状态间接影响种子萌发,并影响植物幼苗的生长、种群实生苗更新及其动态变化。一定范围内适宜的温度可促进植物种子的萌发,过高和过低温度均不利于种子萌发[4-6]。不同植物种子萌发要求的适宜温度差异较大,例如合柱金莲木(Sinia rhodoleuca)种子萌发的最适温度相对较高(25 ℃)[7],而峨参(Anthriscussy lvestris)种子仅有5 ℃[8]。水分是影响荒漠地区植物种子萌发和实生苗更新的主要环境因子,水分胁迫可能是此类地区种子萌发和实生苗更新的重要瓶颈[9],水分胁迫不仅可降低种子萌发率[10-12],延迟种子萌发进程[12-13],而且可能通过破坏细胞渗透调节功能和保护酶系统而影响幼苗生长[12,14]。不同植物的种子萌发对土壤含水量要求有较大差异,例如沙冬青(Ammopiptanthus mongolicus)和沙棘(Hippophae rhamnoides)(15%~20%)[15]、四合木(Tetraena mongoli⁃ca)(20%)[16]种子萌发的适宜土壤含水量相对较高,而沙芥(Pugionium cornum)和斧形沙芥(P.cornum do⁃labratum)种子则较低[17],甚至有些植物种子在1%~2%土壤含水量即可萌发[8-9]。干旱地区植物的种子在低土壤含水量下快速萌发可增加幼苗建植的机会,但降水的不确定性和频繁土壤水分胁迫可能导致幼苗建植后期大量死亡[8]。因此,种子萌发策略是植物对分布区气候条件和生境长期适应的结果[18],干旱地区植物种子通常采取持续性萌发和出苗策略以降低集中萌发造成的幼苗死亡风险[19-20]。
柠条(Caragana korshinskii)是豆科锦鸡儿属旱生灌木,主要分布于沙地草场、荒漠或半荒漠地区的流动沙地和半固定沙地,是我国西北荒漠地区水土保持和防风固沙的主要灌木树种之一[21]。已有研究在实验室条件下证实了柠条种子在15%土壤含水量时萌发率最高(72%),但在10%含水量时仍在65%以上[22],模拟降水研究还发现,每3 d 供水5.0 mm时柠条种子出苗率为59.0%[23],但在单次供水量2.5 mm 时仅有不足10%的种子能够出苗[24]。曾彦军等[25]通过PEG-6000模拟干旱胁迫验证了柠条种子萌发对水分条件的较强适应性。尽管已有研究结果表明,温度与PEG模拟干旱胁迫的交互作用显著影响柠条种子萌发[26],但PEG溶液模拟干旱胁迫难以模拟低土壤含水量对种子萌发的胁迫作用,尤其是柠条种子萌发对不同温度和土壤含水量及其交互作用的响应方面还缺乏深入了解。本文在人工气候箱控制的6 个恒温条件下,每一温度下各设置6 个梯度的土壤含水量处理,研究了不同温度和土壤含水量及其交互作用对柠条种子萌发的影响,研究结果可为西北干旱荒漠地区柠条灌丛群落的人工辅助实生苗更新及退化植被的恢复提供重要参考。
1 材料与方法
1.1 种子采集与处理
试验用种子于2018 年6 月下旬采自宁夏灵武白芨滩国家级自然保护区内的龙坑林区(106°30′E,37°53′N)人工栽植的树龄20 a 以上的成年柠条植株。摘取外形饱满的黄褐色成熟荚果带回实验室自然风干,人工清除果荚和虫蛀的种子,将完好种子装入透气布袋内,于室温下贮藏3 个月后用于萌发试验。
1.2 试验设计和播种方法
有关温度[27-28]、土壤含水量[17,22]对于柠条及其他种子萌发影响的研究中,试验设置的温度范围为5~30 ℃,土壤含水量的范围为1%~30%,而温度与土壤含水量双因素影响下种子萌发情况的研究鲜有报道。因此,本研究在智能人工气候箱控制的6 个恒温条件(5 ℃、10 ℃、15 ℃、20 ℃、25 ℃、30 ℃)下,将取自宁夏灵武白芨滩国家级自然保护区内龙坑林区柠条人工林表层1~5 cm的风沙土作为基质,每一温度分别设置6 个土壤含水量(4%、8%、12%、16%、20%、30%),共36 个处理,每一处理设置4 次重复,各重复分别取大小基本一致的柠条种子50粒播于直径为90 mm的培养皿中。播种时将种子置于培养皿中湿润沙层表面,将播种后的培养皿分别称重记录后置于不同温度的人工气候箱中,设置14 h光照/10 h 黑暗(光照强度约11500 LUX)的光周期。为避免试验期间水分因种子吸收和蒸发引起土壤含水量的波动对试验结果的影响,在试验期间每24 h将所有培养皿取出用1/100电子天平称重,根据培养皿重量变化补充蒸发损失的水分。
1.3 数据记录和萌发参数计算
以胚根伸出种皮2 mm为萌发标准,每24 h观察记录种子萌发情况1次,记录萌发种子数,并测定已萌发种子的胚根长(mm),记录后取出已萌发种子,观察记录至连续14 d 不再有种子萌发为止。分别按下面公式计算种子萌发率(germination percent⁃age, GP)、萌发速率(germination rate,GR)、萌发指数(germination index,GI)和活力指数(vigor index,VI)等萌发参数。
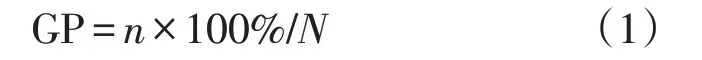
式中:n和N分别为萌发种子数和试验用种子总数。

式中:n为每一处理使用的种子数;Gi为ti(ti=0,1,2,3,···)天萌发的种子数;萌发速率越大,表示萌发越快[29]。

式中:Gt为时间t天的萌发种子数;Dt为萌发试验天数。
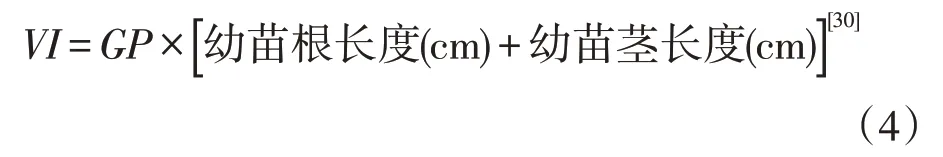
本研究只计算幼苗根长度即胚根长度。
1.4 数据处理
采用双因素方差分析(Two way ANOVA)的方法分析温度和土壤含水量及其交互作用对各种子萌发参数的影响,用单因素方差分析(One way ANO⁃VA)的最小显著差法(LSD)分析各萌发参数在不同温度间和同一温度的不同土壤含水量处理间的差异显著性。为使数据符合样本方差齐性的要求,将全部试验数据在进行统计分析前进行平方根转换,所有统计分析均在SPSS 25.0软件中进行。
2 结果与分析
2.1 温度对柠条种子萌发的影响
从表1可以看出,温度、土壤含水量以及两者的交互作用对柠条种子的萌发率、萌发速率、萌发指数和活力指数均具有极显著影响(P<0.01)。以温度为例,种子萌发率在5 ℃最低,温度为10 ℃时达最大值后又随温度升高逐渐降低,在10 ℃、15 ℃、20 ℃和25 ℃种子萌发率均显著高于5 ℃和30 ℃(P<0.05)。萌发速率和萌发指数也均在5 ℃最小,在25 ℃最大,其中萌发速率在20 ℃、25 ℃和30 ℃显著高于5 ℃与10 ℃、15 ℃,5 ℃、10 ℃和15 ℃三者间差异显著(P<0.05);萌发指数在25 ℃和30 ℃均显著大于3 个较低温度处理(5 ℃、10 ℃和15 ℃)(P<0.05),15 ℃和20 ℃处理显著大于5 ℃和10 ℃处理(P<0.05)。5 ℃的活力指数显著小于其他温度处理,随温度升高呈先升高后降低趋势,在15 ℃最大,显著高于20 ℃外的其他温度处理(P<0.05)。
2.2 不同温度和土壤含水量处理下种子的萌发率
土壤含水量显著影响柠条种子的萌发率(P<0.01),其与温度间的交互作用对种子萌发率的影响也达显著水平(P<0.01)(表1)。从图1可以看出,种子萌发率在5 ℃随土壤含水量增加而逐渐提高,12%及其以上的土壤含水量处理均显著高于8%处理(P<0.05),4%土壤含水量处理种子萌发率为0。在10 ℃时,土壤含水量为4%时的萌发率(53.50%)显著低于其他处理(P<0.05),当土壤含水量由4%增至20%时,萌发率逐渐升高,但土壤含水量增加到30%时开始降低,土壤含水量20%显著高于8%和30%处理(P<0.05)。萌发率在15 ℃和20 ℃随土壤含水量变化的趋势与10 ℃基本一致,均在土壤含水量20%时最大;且与8%、12%和16%处理均显著高于4%和30%处理(P<0.05)。在25 ℃和30 ℃,种子萌发率随土壤含水量升高表现为类似的变化趋势,均在16%处理最高;其中25 ℃,8%、12%、16%和20%均显著高于4%和30%土壤含水量(P<0.05),30 ℃,30%处理的最小值显著低于其他土壤含水量(P<0.05)。

图1 不同温度和土壤含水量处理柠条种子的萌发率Fig.1 Germination percentage of Caragana korshinskii seeds under different temperatures and soil water contents
2.3 不同温度和土壤含水量处理下种子的萌发速率
土壤含水量及其与温度间的交互作用对柠条种子的萌发速率均具有显著影响(P<0.01)(表1)。从图2 可以看出,萌发速率在5 ℃随土壤含水量增加逐渐提高,16%、20%和30%均显著大于8%和12%处理(P<0.05),8%和12%处理间差异也显著(P<0.05)。萌发速率在10 ℃随土壤含水量增加呈先升高后下降的趋势,土壤含水量为20%时,显著大于其他处理(P<0.05),30%略有降低,但仍显著大于4%的最小值(P<0.05)。在15 ℃和20℃,萌发速率均在土壤含水量4%时最小,20%时最大,其中在15 ℃不同土壤含水量处理间均差异显著(P<0.05),在20 ℃,20%处理显著大于16%外的其他处理(P<0.05),4%时显著小于其他处理(P<0.05)。在25 ℃,萌发速率在土壤含水量为12%和16%时均显著大于其他处理(P<0.05),20%时显著大于4%、8%和30%处理(P<0.05)。在30 ℃,萌发速率在16%土壤含水量处理的最大值与次之的12%处理均显著大于其他处理(P<0.05),在30%处理的最小值与4%处理均显著小于其他处理(P<0.05)。
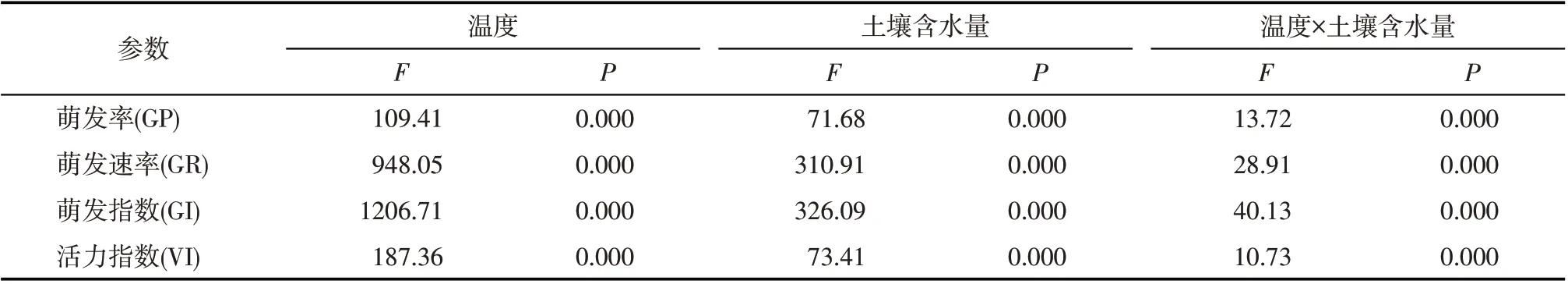
表1 温度和土壤含水量对柠条种子萌发影响的双因素方差分析Tab.1 Two way ANOVA of effects of temperature and soil water content(SWC)on the germination of Caragana korshinskii seeds
2.4 不同温度和土壤含水量处理下种子的萌发指数
土壤含水量及其与温度间的交互作用均显著影响柠条种子的萌发指数(P<0.01)(表1)。从图3 可以看出,萌发指数在5 ℃随土壤含水量增加逐渐增大,土壤含水量为16%、20%和30%时均显著大于8%和12%处理(P<0.05),8%和12%处理间也差异显著(P<0.05)。在10 ℃,土壤含水量4%时,种子的萌发指数显著小于其他处理(P<0.05),20%最高。萌发指数在15 ℃和20 ℃也均在4%时最小,20%最大,其中在15 ℃不同土壤含水量处理间均差异显著(P<0.05),20 ℃的12%及其以上土壤含水量均显著大于4%和8%(P<0.05),但30%显著小于16%和20%处理(P<0.05)。在25 ℃和30 ℃,萌发指数均随土壤含水量增加先升高后降低,且均在16%时最大,30%最小,其中25 ℃的最大值显著大于12%土壤含水量外的其他所有处理(P<0.05),30 ℃的最大值和次之的12%处理均显著大于其他处理(P<0.05);2 个温度下(25 ℃和30 ℃)萌发指数最小值均显著小于其他所有处理(P<0.05)。

图3 不同温度和土壤含水量处理柠条种子的萌发指数Fig.3 Germination index of Caragana korshinskii seeds under different temperatures and soil water contents
2.5 不同温度和土壤含水量处理下种子的活力指数
土壤含水量显著影响柠条种子的活力指数(P<0.01),且其与温度间的交互作用对柠条种子活力指数均呈显著影响(P<0.01)(表1)。从图4可以看出,柠条种子活力指数在5 ℃随土壤含水量增加而增大,3个较高含水量(16%、20%和30%)均显著大于8%和12%处理(P<0.05)。10 ℃土壤含水量为4%时显著小于其他处理(P<0.05),随土壤含水量增加先升高后降低,16%和20%显著大于8%和30%处理(P<0.05)。活力指数在15 ℃和20 ℃也均随含水量增加先升高后降低,含水量为20%时最高,其中15 ℃的12%、16%和20%处理均显著高于4%和30%处理(P<0.05);20 ℃的16%和20%处理显著高于4%、8%和30%处理(P<0.05)。随着土壤含水量升高,活力指数在25 ℃和30 ℃均呈先升后降趋势,分别在20%和16%时最高,且均显著大于其他处理(P<0.05)。

图4 不同温度和土壤含水量处理柠条种子的活力指数Fig.4 Vigor index of Caragana korshinskii seeds under different temperatures and soil water contents
3 讨论
种子萌发策略在植物实生苗更新、种群分布格局和数量动态等方面具有重要作用,且对植物群落演替和退化植被恢复过程具有重要意义[31]。干旱荒漠地区植物的种子萌发因受多变的环境因素影响可能具有更大的不确定性[26]。温度是影响植物种子萌发的主要环境因子,一定范围内较高的温度促进种子萌发,超过这一特定范围的温度均不利于种子萌发[6-7,32]。大量研究结果显示,20~25 ℃是多数植物种子萌发的适宜温度范围[6]。尽管一些研究发现,相对较低的温度可通过降低萌发率和推迟萌发进程而抑制种子萌发[5,8],但种子在较低温度下萌发可为幼苗在干旱季节到来前赢得生长时间,从而保证幼苗根系充分发育以提高耐受干旱胁迫能力[31]。
本研究发现,温度显著影响柠条种子的萌发,在10 ℃萌发率最高(87.5%),在15 ℃、20 ℃和25 ℃萌发率也均在75%以上,萌发速率、萌发指数和活力指数均随温度的升高先升高后降低,且均在温度超过25 ℃时大幅降低。尽管种子在30 ℃条件下萌发较快,萌发指数较大,但萌发率和活力指数大幅降低。基于对4 个萌发参数的综合分析,笔者认为柠条种子萌发的适宜温度范围应为10~25 ℃;与相关研究报道的柠条种子萌发的适宜温度范围(15~20 ℃)相比[26],本研究中柠条种子萌发的适宜温度范围略宽,可能由于以土壤为萌发基质更接近于干旱荒漠区早春季节温差较大的环境条件,适宜种子萌发的较宽温度范围可能有利于抵消水分胁迫造成的负面影响。种子萌发行为是植物对分布区自然生境长期适应的结果,成熟散落于夏季高温季节的柠条种子如果遇有少量降水即可快速萌发成苗,但随之而来的土壤水分亏缺可能导致幼苗大量死亡[31]。因此,柠条种子在相对较高温度下的萌发抑制可能是保证少量种子进入土壤种子库并逃避幼苗死亡的保护性策略,而种子在春季低温下的萌发可在一定程度上对这一策略进行补偿[26]。
种子萌发和幼苗更新期间由降雨量时空分布的高度不确定性及单次降雨伴随的长时间剧烈干旱导致的土壤水分亏缺是限制荒漠地区植物种群实生更新的重要瓶颈[9]。土壤含水量不足可引起种子吸胀困难,从而导致胚根无法突破种皮抑制萌发[33],如果土壤水分不能维持胚根继续伸长和生长,即使已经吸胀的种子也会中断萌发[16],因为胚根和胚轴生长在萌发期间对水分亏缺的反应要比萌发起始阶段更为敏感[34]。荒漠地区植物对土壤水分亏缺的耐受能力相对较强,种子萌发要求的土壤含水量一般相对较低,甚至一些植物的种子在极端土壤含水量下也可完成萌发过程[17]。有研究结果显示,柠条种子可在较低供水量下完成萌发和出苗[24],而较高供水量可能引起部分种子缺氧死亡导致萌发率降低[23],另有研究认为,适度干旱可能对一些植物种子萌发具有一定促进作用[11,35-36],这些结论在本研究中也得到了一定程度验证。
土壤含水量显著影响了柠条种子的萌发。随着土壤含水量的增加,萌发率、萌发速率、萌发指数、活力指数均先增加后减小,且在土壤含水量超过20%时大幅降低。在16%~20%土壤含水量下柠条种子萌发率均在75%以上,其他3个参数也最大,土壤含水量为20%时,萌发率高达87.5%。这些结果表明,柠条种子萌发的适宜土壤含水量范围在16%~20%,这一土壤含水量范围与其他荒漠旱生灌木沙冬青和沙棘(15%~20%)[15]、长叶红砂和蒙古扁桃(Amygdalus mongolica) (16%)[17]和四合木(20%)[16]等种子萌发的最适土壤含水量范围较为接近,但高于荒漠草本植物沙芥和斧形沙芥种子萌发的适宜土壤含水量(6%~12%)[17]。在荒漠地区,种子遇少量降水后快速萌发既可有效利用转瞬即逝的萌发机会,又可为幼苗生长发育赢得充足的时间。然而,一些种子即使在土壤含水量满足萌发要求时,仍有少量保持不萌发状态而进入土壤种子库,以适应不稳定的环境变化,这一策略有效避免了种子集中萌发后因缺乏连续降雨导致的幼苗大量死亡[19]。柠条种子在较低土壤含水量下大量快速萌发无疑将幼苗置于高死亡风险之下,因为单次降雨事件后往往伴随着长时间剧烈干旱,这一推测在孙毅等[31]在毛乌素沙地开展的柠条种子萌发和幼苗存活的跟踪研究中得到了证实。柠条种子萌发对不同土壤含水量的响应模式可能是其在野外生境实生苗缺乏和更新障碍的一种体现,较高土壤含水量对种子萌发的抑制有利于维持种子持续萌发和出苗,从而规避萌发风险[20]。
柠条种子萌发率、萌发指数和活力指数在不同温度下均随土壤含水量增加表现为先增后降趋势,除在5 ℃、30%土壤含水量处理最大外,其他温度下3 个参数均在20%或16%土壤含水量时最大,反映了温度和土壤水分2 个生态因子间的交互作用,即相对充足的土壤含水量能够缓解不适宜温度条件对种子萌发的负面影响,又反映了适宜温度下的低强度干旱对种子萌发的促进作用[11,14,36]。上述结果与吴漫玲等[11]以水杉(Metasequoia glyptostroboides)种子为材料的研究结果类似,即在较低温度下适度干旱可一定程度上促进水杉种子的萌发。柠条种子萌发速率也表现出与其他萌发参数的类似变化,在5 ℃随土壤含水量增加逐渐增大,而10 ℃、15 ℃和20 ℃在20%土壤含水量时最大,25 ℃和30 ℃在16%时最大。低温与低土壤含水量、高温与高土壤含水量均抑制了柠条种子的萌发;适宜的温度与土壤含水量可以有效促进柠条种子的萌发。当温度为15 ℃,土壤含水量为20%时,柠条种子的萌发率与活力指数最大。因此,柠条种子萌发的适宜温度与土壤含水量分别为15 ℃、20%。
柠条种子在相对较低的温度和土壤含水量下即可萌发,但相对较高的温度和土壤含水量抑制种子萌发,这一萌发策略有利于夏季成熟散落的种子进入土壤种子库而逃避死亡,受土壤种子库屏障保护的休眠种子在来年早春的较低温度下遇少量降水即可萌发,并保证充足时间发育出完善的根系,这一结果不仅可从春季较低温度下适度干旱促进种子萌发的结论[11]得以验证,而且与不同温度下PEG 模拟干旱胁迫下的研究结论[26]基本一致,即柠条种子萌发在低温下对干旱胁迫具有更强的耐性。对夏季成熟散落的种子来说,即使其在高土壤含水量条件下能够保持部分水合状态也不会萌发,有研究将其视为一种特殊类型的休眠种子[37],此类种子一旦错过雨季并再次脱水后也会成为土壤种子库的成分,并为来年温度和土壤含水量相对较低的早春季节萌发提供种源。因此,春季低温和低土壤含水量对柠条萌发的促进作用可能是夏季相对较高的温度和土壤含水量对萌发抑制的一种补偿作用,要深入了解这一补偿作用的机制,仍需结合野外自然生境的土壤含水量监测和室内模拟试验开展大量研究工作。
4 结论
柠条种子萌发对温度、土壤含水量及其交互作用具有极显著的反应。种子萌发的适宜温度范围为10~25 ℃,较宽温度范围可能有利于抵消干旱荒漠区早春季节温差较大的环境下水分胁迫造成的负面影响。土壤含水量在16%~20%之间,有效促进了种子萌发。不同温度下,萌发率等指数随土壤含水量的增加均呈先升后降趋势;15 ℃下20%土壤含水量处理有效促进了柠条种子的萌发。因此,夏秋季节较高的温度和土壤含水量对柠条种子萌发的抑制可能是保证部分种子进入土壤种子库并降低幼苗死亡风险的保护性策略,而种子在早春的较低温度和土壤含水量条件下萌发可能是对这种抑制作用的一种补偿。
