牡丹根腐病病株和健株根内微生物菌群的差异
2022-02-02姬俊华杨瑞先王奥一李静宜孙高峰何艳红
姬俊华,杨瑞先,刘 萍,王奥一,李静宜,孙高峰,何艳红,杜 凡
(洛阳理工学院 环境工程与化学学院,河南 洛阳 471023)
牡 丹(Paeonia suffruticosa)属 芍 药 科(Paeoniaceae)芍 药 属(Paeonia)牡 丹 组(Sect.Moutan)多年生落叶灌木,是我国传统名花,兼具观赏和药用价值[1]。近年来,连作导致牡丹种植区土传病害发病日益严重,一定程度上制约了牡丹产业的发展。其中,牡丹根腐病(又称烂根病)是典型的土传病害,在牡丹的整个生育期均可造成危害。在牡丹主要种植区,根腐病发病率一般为20%,严重地块可达40%以上,影响牡丹观赏价值和药用价值,制约牡丹产业可持续发展[2]。前人对牡丹根腐病的研究主要集中在病原菌的分离及防治[3-4]方面,认为引起根腐病的病原菌种类主要为腐皮镰刀菌(Fusarium solani),同 时 认 为,尖 孢 镰 刀 菌(F.oxysporum)和丝核菌属(Rhizoctoniasp.)、腐霉属(Pythiumsp.)、拟盘多毛孢属(Pestalotiopsissp.)真菌作为常见土壤习居菌,在根腐病的发生和发展过程中,往往与腐皮镰刀菌混合侵染,加重根腐病的发生[5]。这表明牡丹根腐病的发生与根际其他腐生菌也密切相关。
目前,植物健康状况与其根际或根部微生物群落结构之间的关系已成为研究热点。植物病害的发生与植物根际、根面和根内微生物种群结构的改变存在一定的相关性,尤其是随着高通量测序技术的快速发展,研究者已能从微生物群落水平分析影响植物病害发生的主要微生物种类。例如,WU等[6]通过高通量测序对比研究三七健康植株及根腐病植株根内微生物发现,健康与患病植株根内细菌和真菌种群组成不同,患病植株根部组织中假单胞菌属(Pseudomonas)细菌和土赤壳菌属(Ilyonectria)真菌丰度显著高于健康植株,表明三七根腐病的发生与根内微生物种群结构的改变密切相关。ZHANG等[7]研究发现,感染柑橘黄龙病植株样本和健康植株样本中根面细菌种群丰度不同,且细菌功能基因表达水平不同,其中,慢生根瘤菌(Bradyrhizobium)和伯克霍尔德菌(Burkholderia)在病株根面显著富集,而土壤杆菌(Agrobacterium)和假单胞菌的相对丰度较健株根系明显降低。
牡丹根腐病作为一种土传病害,其发生与根部组织微生物群落结构的关系目前尚无相关研究。探究牡丹健株和根腐病病株微生物群落结构差异等,能够揭示根腐病发生与根部组织微生物群落之间的关系,也可为进一步获取对牡丹根腐病有抑制作用的有益微生物提供新资源。基于此,拟以牡丹健株和根腐病病株根部组织为研究对象,利用Illumina MiSeq 高通量测序技术分析牡丹根部组织中细菌及真菌群落结构的变化,以探究牡丹健株与根腐病病株根部组织中细菌与真菌的种群结构组成及差异,揭示根腐病发生与微生物群落结构变化之间的关系,为生防菌筛选和利用微生态手段防控牡丹根腐病的发生提供理论基础,促进牡丹产业可持续发展。
1 材料和方法
1.1 样本采集与处理
健株和根腐病病株根部组织样本于2018 年3月采集自河南省洛阳市隋唐城遗址植物公园牡丹种植区(34°64′21″N,112°45′36″E),采样品种为洛阳红,已种植10 a。当地年平均气温约15 ℃,年降雨量约630 mm。2018 年3 月发现该种植区中部分牡丹植株生长衰弱,叶片变小、发黄或泛红,枝条细弱,呈根腐病典型发病症状。在种植区中选择典型根腐病病株15 株,采集其发病根部组织,作为牡丹根腐病病株样本(PDR),并于管理一致的种植区内采集健康牡丹根部组织样本(距离采样根腐病植株样本500 m 以外)作为牡丹健株样本(PHR)。采用5点取样法,每5 棵牡丹记为1 次重复,根腐病病株和健株样本各3 次重复,分别标记为PDR1—3 和PHR1—3。将采集的样本装入无菌袋内,带回实验室用于后续试验,根部组织表面用无菌水冲洗干净,用70%乙醇浸泡30 s,用5%次氯酸钠溶液消毒3 min,再用无菌水冲洗5 次,晾干,用于组织样本DNA的提取。
1.2 样本DNA提取与测序
采 用FastDNA®SPIN Soil for kit 试 剂 盒(MP Biomedicals,Solon,USA)提取牡丹根部组织DNA;利用Nanodrop 2000(Thermo Scientific,Wilmington,USA)检测DNA 样本的浓度和纯度;利用1%的琼脂糖凝胶电泳进行DNA 质量检测。检测合格的DNA用于后续PCR扩增。
以牡丹根部组织样本DNA 为模板,使用引物799F(5′-AACMGGATTAGATACCCKG-3′)和1193R(5′-ACGTCATCCCCACCTTCC-3′)对细菌16S rRNA基因的V5—V7 可变区进行扩增;使用引物ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2R(5′-GCTGCG TTCTTCATC GATGC-3′)对真菌ITS基因进行扩增。PCR 扩增反应体系为20 µL,反应程序:95 ℃预变性3 min,27 个循环(95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s),最后72 ℃延伸10 min。扩增后取3µL 利用2%琼脂糖凝胶电泳进行检测,PCR 产物纯化后利用Illumina MiSeq PE300平台对扩增产物进行双端测序。测序工作委托上海美吉生物医药科技有限公司完成。
1.3 数据质控与分析
利用Trimmomatic 0.33 软件和FLASH 1.2 软件对测序原始数据进行质控,过滤掉低质量的Read序列,并将得到的双端序列数据拼接成Tag,利用UCHIME 8.1 软件去除低质量的Tag 和嵌合体。使用UPARSE 7.1 软件对有效Tag 在97%的相似度水平下聚类成为操作分类单元(Operational taxonomic units,OTU)。基于Silva 细菌分类数据库和Unite 真菌分类学数据库,利用RDP Classifier 2.11 软件对OTU 进行分类学注释,置信度阈值为0.7,得到每个OTU 对应的物种分类信息,进而在各水平统计样本群落组成。利用Mothur 1.30 软件评估样本的Alpha多样性指数,用QIIME 1.9.1软件进行样本Beta多样性分析,利用FUNGuild 软件对真菌分类进行注释[8]。以上分析利用上海美吉生物医药科技有限公司I-Sanger 生信云网站平台(http://www.i-sanger.com/project/index.html)完成,所有统计分析均采用R3.1.2(http://www.r-project.org/)软件完成。
2 结果与分析
2.1 牡丹根部组织样本微生物基因序列深度分析和数据质控
本研究采集了6 个牡丹根部组织样本,经过数据质控,从6 个样本中获得细菌16S rRNA 基因V5—V7 区高质量序列(Clean tag)136 078 条,每个样本中Clean tag 数为15 866—24 869 条,序列平均长度为377 bp;从6个样本中获得真菌ITS基因高质量序列313 753 条,每个样本中Clean tag 数为38 636—66 509条,序列平均长度为238 bp。
2.2 牡丹根部组织样本微生物OTU聚类分析
在97%相似度水平对样本序列进行OTU 聚类,从健康牡丹根部组织中共鉴定获得细菌21个门,40个纲,106 个目,196 个科,316 个属,476 个种,632 个OTU;感染根腐病的根部样本中共鉴定获得细菌20个门,31 个纲,97 个目,169 个科,279 个属,424 个种,564 个OTU。Venn 图分析结果(图1A、C)表明,在OTU 和属水平,健株样本与感染根腐病样本之间细菌种类较为接近,共有的OTU 和属的种类远高于2组样本独有的种类。
从健康牡丹根部组织中共鉴定获得真菌9 个门,31 个纲,64 个目,122 个科,184 个属,223 个种,360 个OTU;感染根腐病的根部样本中共鉴定出真菌7 个门,25 个纲,51 个目,84 个科,112 个属,136个种,215 个OTU。Venn 图分析结果(图1B、D)表明,在OTU 和属水平,健株样本与感染根腐病样本之间真菌种类差异较大,共有的OTU 种类低于2 组样本独有的种类,在属分类水平上,健康组织样本中独有种类也远高于2组样本共有的种类。
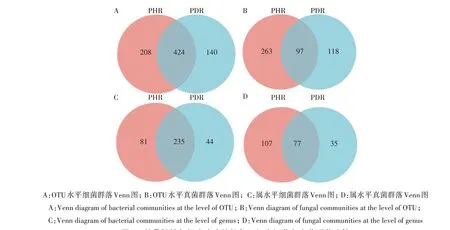
图1 牡丹健株与根腐病病株根部组织中细菌和真菌群落比较Fig.1 Comparison of bacterial and fungal communities in the root tissues of healthy and root rot-diseased tree peony
2.3 牡丹根部组织样本微生物Alpha多样性分析
对健株根部组织和根腐病发病组织样本中的细菌、真菌Alpha 多样性指数ACE、Chao、Shannon、Simpson 及Sob 进行分析,结果见表1。其中,ACE、Chao和Sob指数用于表示样本细菌及真菌群落丰富度;Shannon、Simpson指数用于表示样本中细菌及真菌群落多样性。上述指数中,Simpson 指数值越大,表明样本中群落多样性越低,其余指数值越大,说明样本中群落丰富度和多样性越高。由表1 可知,健株中细菌的ACE、Chao 和Sob 指数均高于病株,表明健株根部组织中细菌群落相对丰富,但差异不显著;Shannon、Simpson指数与病株差异较小。以上表明,健株与病株组织样本中细菌种群丰富度及多样性差异均不明显。同时,从表1可知,健株根部组织样本中真菌的ACE、Chao 和Sob 指数均明显高于病株,其Shannon 指数(P=0.04)、Simpson 指数(P=0.05)与病株具有显著性差异,表明健株根部组织样本中真菌丰富度和多样性均高于病株。以上表明,健株与病株之间的细菌种群多样性无显著差别,但在真菌多样性方面存在显著差异,相比健株,病株中真菌种群多样性显著降低。
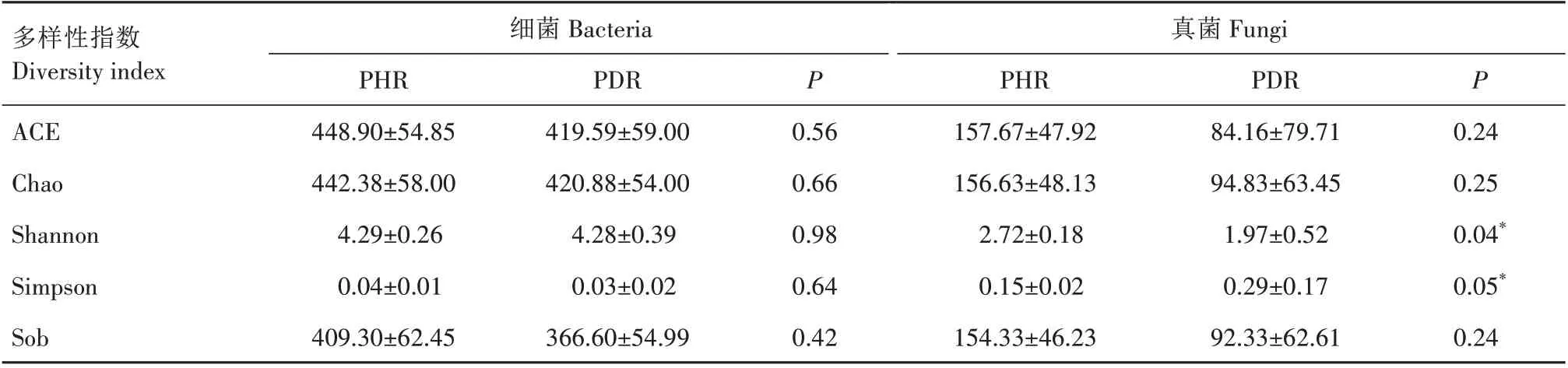
表1 牡丹健株与根腐病病株根部组织中细菌和真菌Alpha多样性Tab.1 Alpha diversity of bacterial and fungal communities in the root tissues of healthy and root rot-diseased tree peony
2.4 牡丹根部组织样本细菌群落基本组成和结构分析
由图2可知,在门水平上,健康牡丹根部组织样本中细菌种群主要分布在放线菌门(Actinobacteria,58.55%)、变形菌门(Proteobacteria,36.87%)、拟杆菌门(Bacteroidetes,0.92%)、绿弯菌门(Chloroflexi),0.81%)、酸杆菌门(Acidobacteria,0.74%)和厚壁菌门(Firmicutes,0.70%);根腐病发病根部组织样本中细菌种群主要分布在变形菌门(70.9%)、放线菌门(21.16%)、拟杆菌门(6.08%)、酸杆菌门(1.04%)、绿弯菌门(0.09%)和厚壁菌门(0.05%)。细菌群落组成表明,与健株相比,患病根部组织中变形菌门、拟杆菌门、酸杆菌门的相对丰度增加,分别增加34.03、5.16、0.30 个百分点;而放线菌门、厚壁菌门和绿弯菌门的相对丰度降低,分别降低37.39、65.00、0.72个百分点。
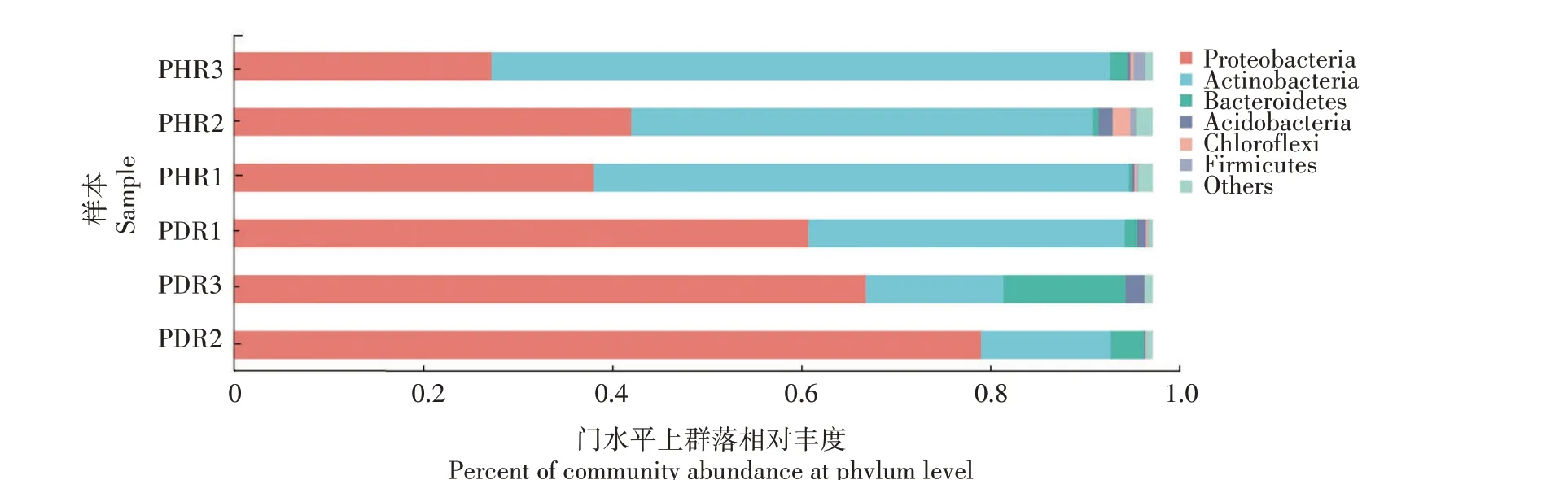
图2 牡丹根部组织样本细菌门水平上群落的相对丰度Fig.2 Relative abundance of bacterial communtiy in tree peony root tissues samples at phylum level
由图3可知,在属水平上,健康牡丹根部组织中细菌优势属主要为假诺卡氏菌属(Pseudonocardia,14.90%)、植物栖居菌属(Phytohabitans,8.50%)、分枝 杆 菌 属(Mycobacterium, 6.49%)、Norank_f__Rhodobacteraceae(5.39%)、弗兰克菌属(Frankia,4.58%)、固 醇 杆 菌 属(Steroidobacter, 2.84%)、Unclassified_f__Burkholderiaceae(1.94%)、链霉菌属(Streptomyces, 1.41%) 和 Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium(1.01%)。根腐病发病根部组织样本中细菌优势属主要为Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium(12.14%)、假单胞菌属(Pseudomonas,6.56%)、新鞘氨醇杆菌属(Novosphingobium,5.63%)、链霉菌属(4.97%)、Unclassified_f__Burkholderiaceae(4.57%)、鞘脂菌属(Sphingobium, 3.90%)、固醇杆菌属(3.22%)、Burkholderia-Caballeronia-Paraburkholderia(2.23%)、Unclassified_f__Microscillaceae(1.74%)、假诺卡氏菌属(1.48%)。健株与根腐病发病组织样本中共有细菌优势属5个,分别为假诺卡氏菌属、固醇杆菌属、Unclassified_f__Burkholderiaceae、链霉菌属 和 Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium。其中,健康组织样本中独有的属为植物栖居菌属、分枝杆菌属、弗兰克菌属和Norank_f__Rhodobacteraceae;患病组织样本中独有的属为假单胞菌属、新鞘氨醇杆菌属、鞘脂菌属、Burkholderia-Caballeronia-Paraburkholderia 和 Unclassified_f__Microscillaceae。以上表明,与健株相比,患病根部组织中的Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium、链霉菌属、Unclassified_f__Burkholderiaceae和固醇杆菌属等细菌属的相对丰度有所增加,分别增加11.13、3.56、2.63、0.38个百分点;而假诺卡氏菌属的相对丰度显著降低,降低了13.42 个百分点。同时推测病株样本中独有的属如假单胞菌属、新鞘氨醇杆菌属、鞘脂菌属的出现可能与牡丹根腐病的发生关系密切。
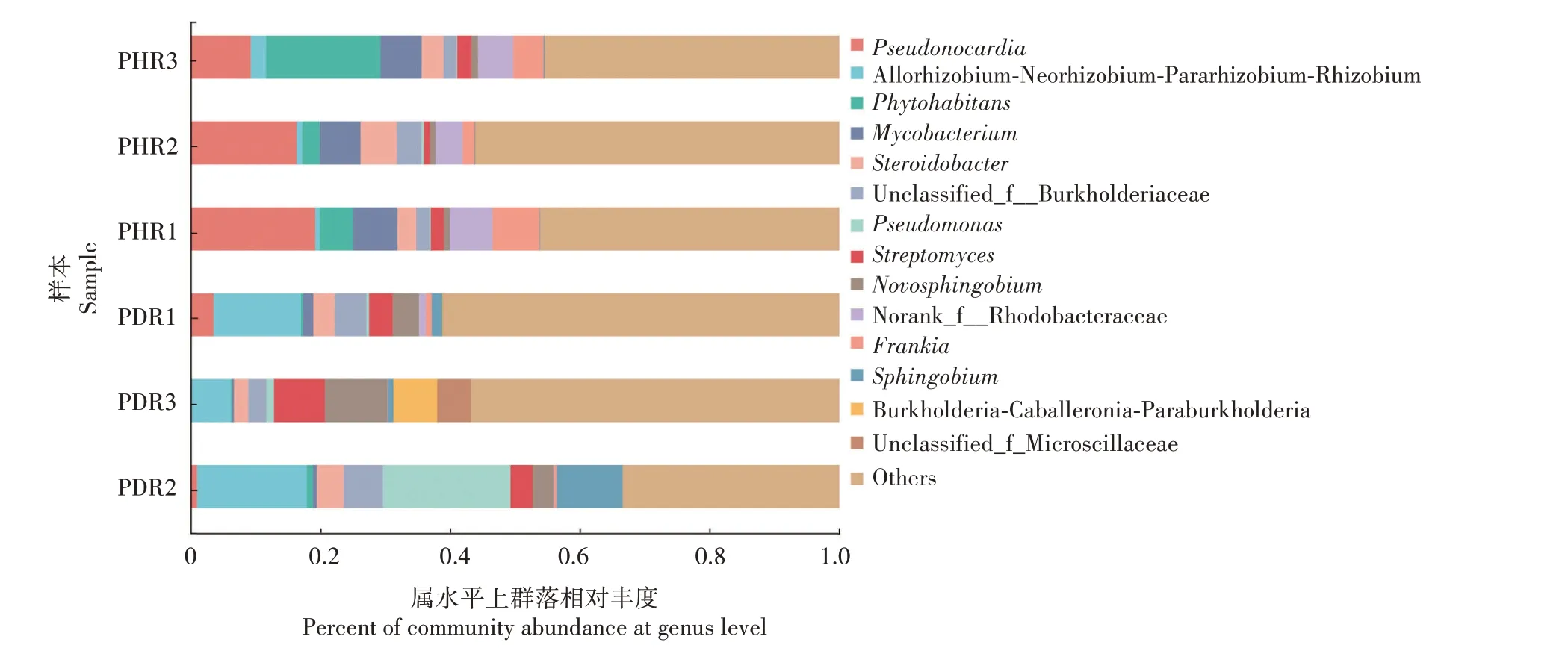
图3 牡丹根部组织样本细菌属水平上群落的相对丰度(相对丰度>0.05)Fig.3 Relative abundance of bacterial communtiy in tree peony root tissues samples at genus level(The relative abundance >0.05)
2.5 牡丹根部组织样本真菌群落基本组成和结构分析
由图4可知,在门水平上,健康牡丹根部组织样本中真菌主要分布在子囊菌门(Ascomycota,79.04%)、担 子 菌 门(Basidiomycota,12.60%)、Unclassified_k__Fungi (7.02%) 、罗 兹 菌 门(Rozellomycota,0.77%)和其他(0.54%);根腐病发病根部组织样本中真菌主要分布在子囊菌门(77.43%)、担子菌门(21.81%)、其他(0.48%)和Unclassified_k__Fungi(0.22%)。真菌群落组成表明,与健株相比,患病根部组织中担子菌门真菌的相对丰度明显增加,增加了9.21 个百分点;而子囊菌门真菌的相对丰度有所降低,降低了1.61 个百分点。
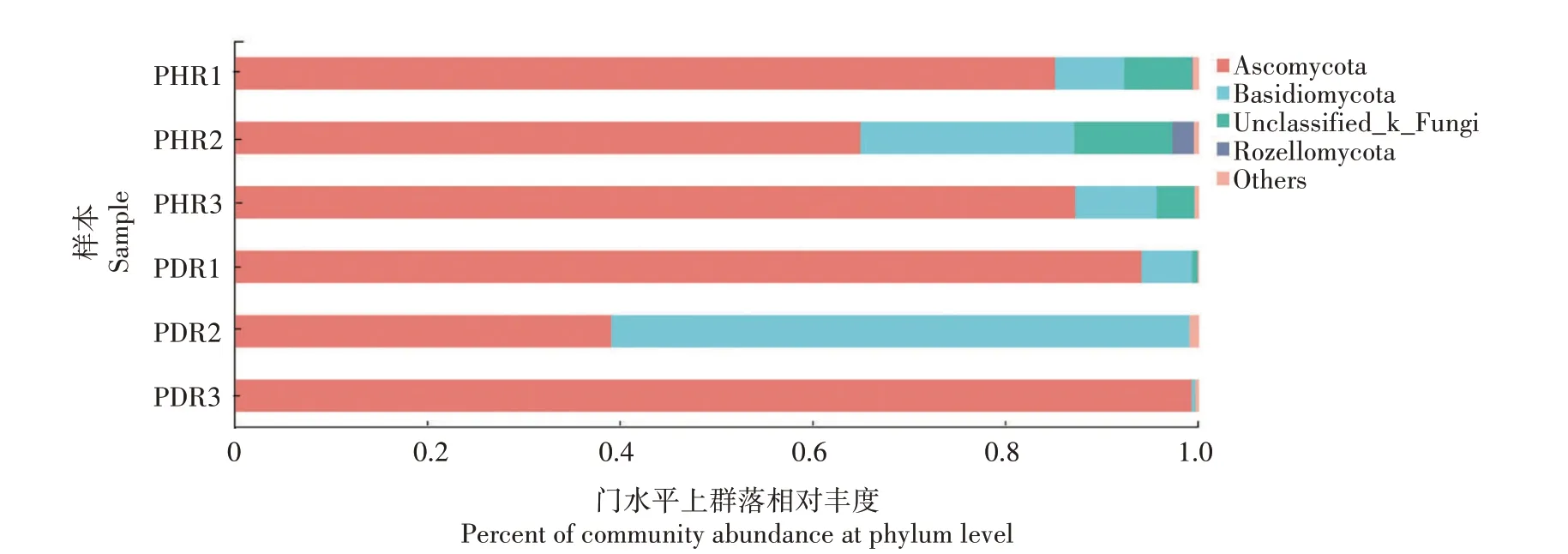
图4 牡丹根部组织样本真菌门水平上群落的相对丰度Fig.4 Relative abundance of fungal community in tree peony root tissues samples at phylum level
由图5可知,在属水平上,健康牡丹根部组织中真菌优势属主要为Unclassified-p-Ascomycota(21.46%) 、其 他(21.46%) 、拟 折 孢 属(Minimelanolocus, 13.03%) 、 Unclassified_c__Sordariomycetes(9.89%) 、Unclassified_k__Fungi(7.04%)、外 瓶 霉 属(Exophiala,5.93%)、Cutaneotrichosporon(5.82%)、交链孢属(Alternaria,3.82%)、Unclassified_o__Chaetothyriales(2.35%)、Hymenula(1.25%)、Dactylonectria(1.24%);根腐病发病根部组织中真菌优势属分别为Hymenula(31.96%)、其他(17.99%)、Kurtzmanomyces(15.58%)、Dactylonectria(12.93%)、Unclassified_f__Nectriaceae(9.36%)、Unclassified_o__Hypocreales(5.58%)、Cuniculitrema(4.04%)、Unclassified-p-Ascomycota(3.18%)、乳 突 赤 壳 属(Thelonectria,3.13%)、Unclassified_o__Sordariales(2.56%)、Unclassified_o__Helotiales(2.05%)、外瓶霉属(1.52%)。健株与根腐病发病组织样本中共有真菌优势属4 个,分别为Unclassified-p-Ascomycota、外瓶霉属、Hymenula和Dactylonectria。其中,健康组织样本中独有的属为拟 折 孢 属 、Unclassified_c__Sordariomycetes、Cutaneotrichosporon、Unclassified_k__Fungi、交 链 孢属和Unclassified_o__Chaetothyriales;患病组织样本中独有的属为Kurtzmanomyces、Unclassified_f __Nectriaceae 、Unclassified _ o __ Hypocreales 、Cuniculitrema、乳 突 赤 壳 属、Unclassified_o__Sordariales、Unclassified_o__Helotiales。以上表明,与健株相比,患病根部组织中的Hymenula和Dactylonectria属相对丰度显著增加,分别增加30.71、11.69 个 百 分 点,Unclassified-p-Ascomycota和外瓶霉属真菌相对丰度明显降低,分别降低为18.28、4.41 个百分点。同时推测病株样本中独有的属如Kurtzmanomyces、Unclassified_f__Nectriaceae 和Unclassified_o__Hypocreales 的出现可能与牡丹根腐病的发生关系密切。
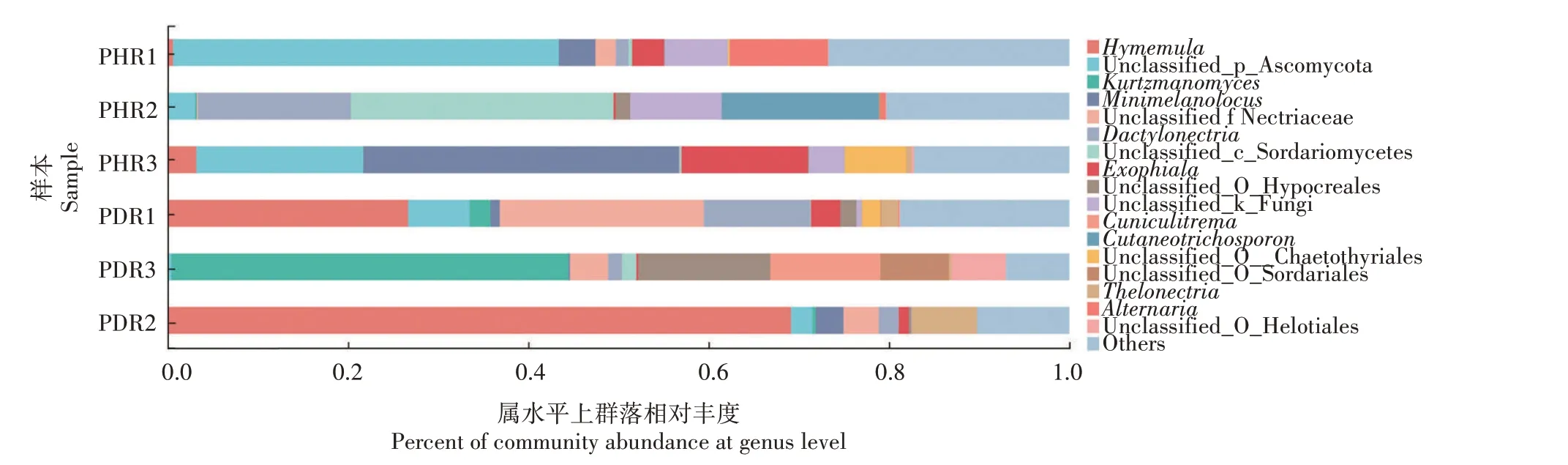
图5 牡丹根部组织样本真菌属水平上群落的相对丰度(相对丰度>0.05)Fig.5 Relative abundance of fungal community in tree peony root tissues samples at genus level(The relative abundance >0.05)
2.6 牡丹根部组织样本间微生物Beta多样性分析
样本距离Heatmap能够反映样本间群落组成的差异,基于Weighted unifrac 距离算法获得细菌距离Heatmap,由图6A 可知,健株与病株根部组织样本间最大距离值为0.500,表明健株与病株根部组织样本间细菌群落结构和多样性具有一定的差异;基于Weighted unifrac 距离算法获得真菌距离Heatmap,由图6B 可知,健株与病株根部组织样本间最大距离值为1.339,表明健株和病株根部组织样本间真菌群落结构和多样性存在显著的差异。以上表明,牡丹健株与病株根部组织样本间细菌、真菌群落结构和多样性均存在一定的差异,但真菌菌群结构差异明显,表明牡丹根腐病的发生对牡丹根部组织中细菌菌群结构的影响较小,但显著改变了牡丹根部组织中真菌的菌群结构。
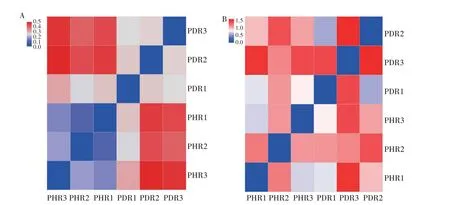
图6 牡丹根部组织样本细菌(A)和真菌(B)OTU水平距离HeatmapFig.6 Heatmap of tree peony root tissues samples at bacterial(A)and fungal(B)OTU level
主成分分析(PCA)能够反映样本间群落组成的差异。健株与病株根部组织样本间细菌群落结构主成分分析(图7A)表明,OTU 水平下第1 主成分(PC1)、第2 主成分(PC2)可以解释所有变量的44.16%、24.74%,2 个主成分方差累积贡献率为71.90%,说明其能够表征微生物群落组成的特征。进一步分析发现,健康根部组织样本主要位于第一、四象限,患病根部组织样本主要位于第二、三象限,健株与病株根部组织样本被明显区分开,表明健株与病株根部组织样本细菌群落结构存在差异。
健株与病株根部组织样本间真菌群落结构主成分分析(图7B)表明,OTU 水平下第1 主成分(PC1)、第2 主成分(PC2)可以解释所有变量的42.84%、31.57%,2 个主成分方差累积贡献率为74.41%,说明其能够表征微生物群落组成的特征。进一步分析发现,健康根部组织样本主要位于第一象限,患病根部组织样本主要位于第三、四象限,健株与病株根部组织样本被明显区分开,表明健株与病株样本间真菌群落结构存在显著差异。
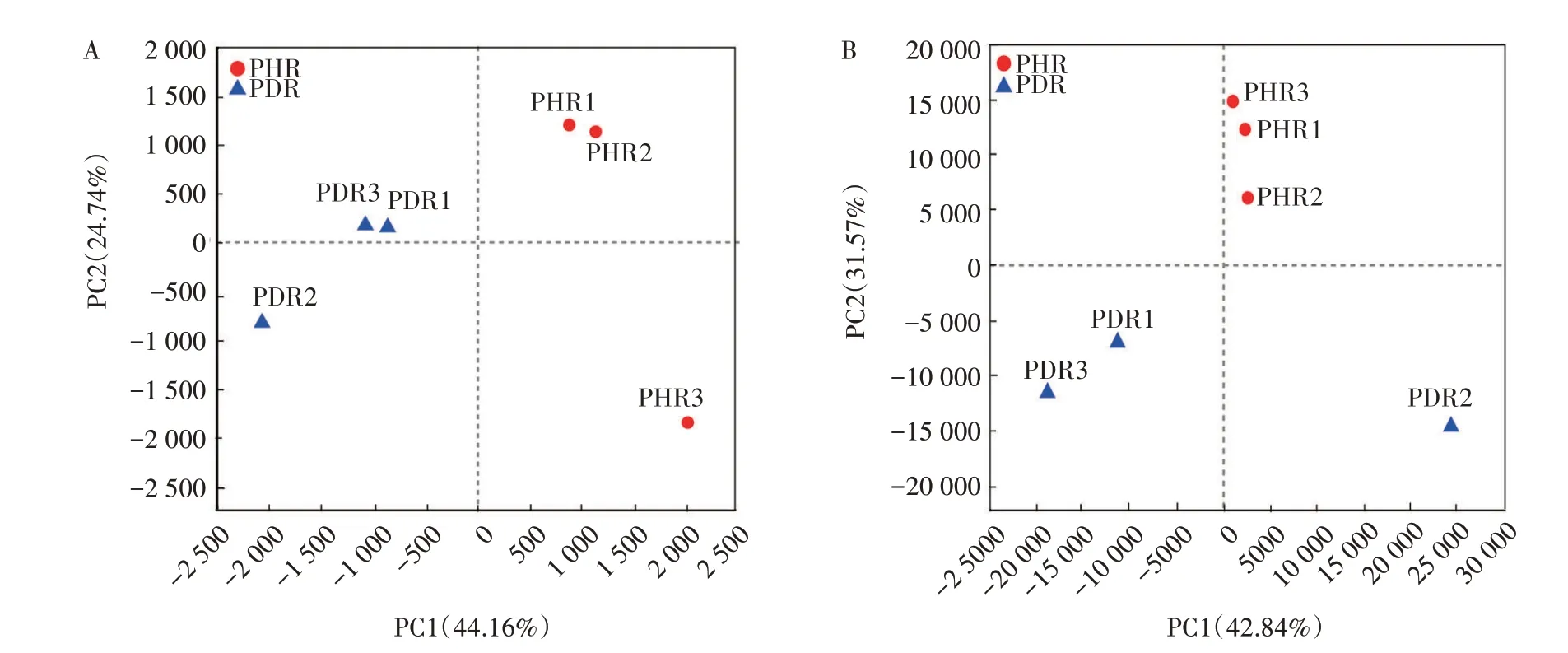
图7 牡丹根部组织样本细菌(A)和真菌(B)OTU水平主成分分析Fig.7 Principal component analysis of tree peony root tissues samples at bacterial(A)and fungal(B)OTU level
2.7 牡丹根部组织样本真菌功能类群预测分析
利用FUNGuild软件对真菌分类进行注释,结果见图8。由图8 可知,患病根部组织中植物病原菌(Plant pathogen)占所有真菌OTU 的27.41%,健康根部组织中植物病原菌仅占所有真菌OTU 的0.04%;患病根部组织中未定义的腐生菌(Undefined saprotroph)(27.38%)的OTU 丰度也显著高于健株(0.10%);健康根部组织中未知真菌(Unknown,45.20%)的OTU丰度显著高于病株(12.31%)。以上表明,牡丹发生根腐病后,其植物病原菌和腐生菌数量在根部组织中占据绝对优势,且真菌种群多样性显著减低。
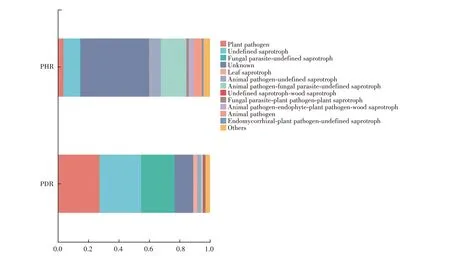
图8 牡丹健株与根腐病病株根部组织中真菌OTU水平功能注释Fig.8 Function annotation at fungal OTU level in the root tissues of the healthy and root rot-diseased tree peony
3 结论与讨论
研究表明,植物根际、根内微生物群落组成与植物土传病害的发生具有一定的关系,丰富的根际、根内微生物种类能显著提高植物抑制土传病害发生的能力[9-10]。本研究中所有样本OTU 聚类及Alpha 多样性分析表明,牡丹根腐病病株中细菌OTU 种类有所降低,种群的丰富度和多样性与健株差异不显著;但健株与根腐病病株间真菌种群多样性差异显著,健株中真菌OTU 种类和种群多样性显著高于病株,这个结果与前人研究相一致。如SHANG 等[11]研究发现,健康兰州百合根际土壤中的真菌多样性显著高于根腐病发病土壤;郑元仙等[12]研究发现,感染根腐病的烟株根际土壤真菌群落多样性水平显著低于健康烟株的根际土壤;文永均等[13]研究发现,健康三七根际土和根内生真菌多样性高于患根腐病三七,且根际土壤中真菌的种群多样性显著高于根内真菌的种群多样性。总体看来,植物根际、根内微生物群落是一个综合体,植物病害的发生与根际、根内微生物群落结构失衡关系密切。
细菌群落结构组成分析发现,病株与健株组织中细菌优势门类组成基本相似,其中患病根部组织中变形菌门相对丰度明显增加,但是放线菌门和厚壁菌门的相对丰度明显降低。杨光柱等[14]研究发现,苹果根腐病病株根际土壤中细菌变形菌门的相对丰度高于健株,其放线菌门的相对丰度低于健株,与本研究结果相一致。变形菌是一类适应性相对较强的细菌,既包含引起动植物生病的病原菌种类,也包含抑制致病菌的有益菌种类[15]。放线菌门细菌含量通常反映土壤的健康状况,其丰度越高,代表土壤质量较高,植物生长状况较好[16-17]。本研究中,健康牡丹组织样本中放线菌门的相对丰度显著高于患病植株,可推测相对于感染根腐病的植株而言,健株能够招募土壤中更多的放线菌进入根部组织,改善其生长状况,提高其抗病能力。同时研究表明,牡丹根系分泌物和脱落物成分复杂,其成分的微小变化可引起根际微生物区系组成上的巨大差异[18-19],因此,推测患病牡丹植株中变形菌数量的增加,可能与患病植株中牡丹根系分泌物的改变有关系。
在细菌属水平上,牡丹根腐病病株与健株组织样本间也存在差异,患病植株中假单胞菌属和新鞘氨醇杆菌属为优势菌属。研究表明,假单胞菌属细菌既是根际促生菌和植物组织内生菌的主要种类,同时又是侵染多种植物的病原菌[20-21]。向立刚等[22]研究发现,感染青枯病的烟株根际土壤样本中假单胞菌属细菌的相对丰度高于健株。新鞘氨醇杆菌属细菌作为降解菌,也广泛存在于植物根际,如孙会忠等[23]在牡丹(凤丹)根际土壤中分离获得了1 株具有良好解磷活性的解磷菌Novosphingobiumsp.YF20。本研究中牡丹根腐病病株中假单胞菌属和新鞘氨醇杆菌属相对丰度的增加,是否为牡丹在感病条件下,应急招募更多的有益细菌,以缓解病害的发生,或这些细菌种群相对丰度的增加加重了根腐病的发生,仍需做进一步的研究和探讨。
真菌群落结构组成分析发现,病株与健株中真菌优势门类组成基本相似,其病株根部组织中担子菌门真菌的相对丰度有所增加,而子囊菌门真菌相对丰度降低,但降低幅度不明显。宋旭红等[24]研究发现,黄连感染根腐病后,其根际土壤中担子菌门真菌的丰度有所增加;另有研究者发现,三七感染根腐病后,其根内子囊菌的丰度有所降低[13],这与本研究结果基本一致。子囊菌门真菌包含了植物常见的内生菌种类,主要参与土壤中有机物质的分解和养分循环,其含量降低可能会导致土壤肥力的下降[25]。担子菌门真菌种类繁多,部分种类能够与植物共生形成菌根,同时又有一些种类作为植物病原菌导致植物病害的发生。本研究中担子菌门真菌丰度增加显著,推测其部分种类可能为牡丹根腐病病原菌,导致其在发病根部组织占据优势地位。
真菌属水平上,牡丹病株与健株样本之间存在显著差异,患病植株中Dactylonectria属为优势菌属,目前Dactylonectria属真菌作为植物根腐病的病原菌相继被报道:LI等[26]调查发现,D.torresensis为云南白芨根腐病的主要致病菌;ZHANG 等[27]报道,D.novozelandica可引起三七根腐病的发生。本研究中,Dactylonectria属真菌在病株根部组织中显著增加,推测Dactylonectria属真菌也可能引起牡丹根腐病的发生,但前期研究认为牡丹根腐病的病原菌主要为镰刀菌属真菌[2]。因此,仅采用传统分离方法对土传病害病原菌进行分离鉴定,并不能够深入了解牡丹根腐病的病原菌种类,高通量测序技术和传统平板分离法的有机结合,将有助于更好地解析牡丹根腐病的病原菌种类及发病机制。
本研究结果表明,牡丹感染根腐病后,其根部组织中细菌、真菌群落丰富度均减低,真菌种群多样性与健株存在显著差异,与根腐病发病相关的病原菌Dactylonectria属及丛赤壳科真菌相对丰度显著增加,表明病原菌数量的变化导致牡丹根部组织中真菌种群多样性的降低,真菌种群平衡被破坏,从而导致根腐病的发生。同时,本研究中健康牡丹根部组织中假诺卡氏菌和植物栖居菌相对丰度较高,假诺卡氏菌为一类稀有放线菌,可产生一些重要抗生素类活性次生代谢产物,可作为植物促生菌改善植物的生长状况[28];植物栖居菌主要是来自植物组织内部的一类放线菌,可产生多样化和新颖的次生代谢产物[29]。因此在后续试验中,可进一步分离纯化健康根部组织中的内生放线菌,并验证菌株及次生代谢产物对牡丹根腐病的生防效果,可为牡丹根腐病的防治提供具有生防潜能的微生物资源。
