西藏玉米田养鹅模式下的土壤碳排放特征
2022-01-28柳文杰刘合满曹丽花
杨 红,柳文杰,刘合满,曹丽花
(1.西藏农牧学院资源与环境学院,西藏 林芝 860000;2.信阳农林学院,河南 信阳 464000)
土壤呼吸作为大气CO2浓度升高的主要因素,所指为土壤中产生CO2的所有代谢过程,包括土壤生物(动物、植物、微生物等)在生命活动过程中所进行的呼吸作用以及土壤有机碳矿化分解等作用产生的CO2,是陆地生态系统土壤碳排放的主要途径[1]。土壤呼吸每年的CO2排放量是全球化石燃料排放量的10倍[2]。在全球气候变化背景下,土壤呼吸碳排放量可能会进一步增加,西藏作为全球气候变化的敏感区,在本世纪中叶增温幅度可达到2.4℃~3.2℃[3],这将激发西藏高原敏感区土壤温室气体的排放,从而加速全球变暖。
农田生态系统作为受人为干预和支配较为频繁的半自然陆地生态系统,农耕方式、水肥管理措施及作物种类均可通过改变土壤的结构、质地及肥力等因素,影响土壤碳排放强度[4]。西藏农田主要集中在“一江两河”流域,是西藏人民赖以生存的主要粮食来源,然而,由于气候恶劣、土壤贫瘠等原因,施肥成了提高作物产量的主要措施,施肥的残留物质,通过淋溶污染地下水,产生的氮氧化物增加大气温室气体浓度,而这可能是导致青藏高原脆弱生态环境被破坏的“致命一击”。更值得注意的是随着全球气候变化的进一步加剧,青藏高原作为气候变化的敏感区,农田土壤碳排放将积极响应于这一变化。
玉米田养鹅(Raising geese in corn field,RGC)是遵循生态学原理,在同一块土地内同时进行种养结合的一种可持续农业生产模式,这种模式可有效提高农业经济收入[5]、维持农田生态系统多样性[6]和杂草防控[7]。然而,“玉米田养鹅”模式在提高农业经济效益的条件下,对土壤碳排放的影响如何尚鲜见报道,从而制约对玉米田养鹅模式生态经济效益的有效评估。青藏高原高寒农田生态系统十分脆弱,正在经历着明显的气候变暖,因此,研究青藏高原玉米田养鹅模式下高寒农田生态系统土壤碳排放,对预测未来大气CO2浓度、改变传统农业措施造成的面源污染及有效评价玉米田养鹅模式的利弊具有重大意义。故本研究以西藏高寒区域玉米田养鹅农业生产模式下的农田土壤为研究对象,分析玉米田养鹅对土壤呼吸速率的影响,以期为科学阐明高寒气候条件下玉米田养鹅模式的固碳效应和为研究青藏高原高寒农田生态系统土壤呼吸速率提供科学依据。
1 材料与方法
1.1 研究区概况
试验区位于藏东南林芝市巴宜区八一镇章麦村,29°33′N,94°21′E,海拔3 100 m,全年均温8.6℃~8.8℃,7月(当地最热月)均温可达18℃;≥10℃的年有效积温2 225.7℃,年平均降雨量634.2 mm,降水主要集中在5~9月,占全年降水的79.1%,年日照时数1 988.6 h左右,年总辐射量为6.1×109~70×109J·m-2,光合有效辐射为2.5×109~3.0×109J·m-2,无霜期177 d。试验地为高寒农田,无遮阴,土壤类型为沙质土,沙粒含量较多。试验地的耕作制度为油菜—冬小麦或油菜—春青稞轮作。2012年种植青稞,于2013年开始长期定位试验。
1.2 试验设计
于2013年5月9日试验组进行玉米播种,所播玉米品种为酒单4号,玉米株、行间间隔分别为30 cm和70 cm,设置保护行。底肥施用复合肥(组成:N33%,P2O517%,K2O17%,有机质20%),用量为240 kg·hm-2。设置玉米田养鹅(RGC)和常规玉米种植(CK),重复3次,每个试验小区20 m2,总面积为120 m2,随机排列,各小区之间间隔1 m,养鹅处理的农田不除杂草,各小区间用0.5 m高的网围住。综合考量小区内杂草的持续供应性和玉米的正常生长,3个试验处理小区的养鹅总数量为10只,并在3个小区内轮换放牧,每个小区方放牧时间相等。于8∶00左右放入试验地,确保鹅的饮水供应充足,19∶00左右收回鹅圈,并一次性投加饲料1 kg。对照使用除草剂玉雕(成分:锈去津,含量:90%;硝磺草酮,含量:10%)来进行杂草管理,施用量为1.65 kg·hm-2,利用背式喷雾器进行近地面喷施,共喷施2次(第1次在播种后,第2次与第1次间隔50 d)。除此之外,RGC和CK的其他管理措施相同。2013年开始试验至2017年期间处理相同。本试验数据采集于2017年5—8月。
1.3 测定项目及方法
采用开路式土壤碳通量测定系统(Li-8100,Li-COR,USA)进行土壤呼吸速率的测定,为降低土壤表面植物生理活动对土壤呼吸速率的影响,测量前24 h在每个小区玉米行间随机选择3个样点,并安装直径为20 cm,高10 cm的土壤环(呈对角线安装),将土壤环高度的2/3插入土壤,环内的植物齐地面剪除。于2017年5月29—31日、6月12—14日、6月28—30日、7月16—18日、7月28—29日、8月6—8日进行土壤呼吸速率及10 cm土壤深度的温度、5 cm深度的体积含水量测定;仪器测定时间步长设为3 min,土壤呼吸、体积含水量和温度的变化取该时段内20次(1 h)测定值的平均数,并将24 h的指标变化作为日变化特征进行统计。
土壤有机质、全氮、全磷和全钾含量测定分别采用重铬酸钾-外加热法、半微量开氏法、HClO4—H2SO4法和NaOH熔融—火焰光度法。土壤速效氮、速效磷、速效钾含量测定分别采用碱解扩散法、0.5 mol·L-1碳酸氢钠浸提—钼锑抗比色法、NH4OAc浸提—火焰光度法,土壤pH值测定采用5∶1水土比,即搅拌1 min,静置30 min后,采用pH测定仪(IQ150,美国IQ公司)进行测定,采用土壤紧实度仪(SC900)测量土壤紧实度。
1.4 数据分析
采用Excel 2007进行数据处理,处理间土壤呼吸、土壤温度和土壤体积含水量之间的差异分析采用单因素方差分析(One-way ANOVA)法,CK与RGC处理土壤呼吸之间的差异分析采用配对样本T检验。作图及相关性分析采用Origin 9.0(Originlab公司,美国)进行。
土壤呼吸温度敏感性Q10值采用指数模型拟合,计算公式为:
Rs=aebT
(1)
Q10=e10b
(2)
式中,Rs为土壤呼吸速率,T为土壤温度,a是当土壤温度为0℃时的土壤呼吸速率,b为温度反应系数,Q10为土壤呼吸温度敏感性系数。
2 结果与分析
2.1 玉米田养鹅模式下土壤理化性质的变化特征
由表1可知,较试验农田初建时的养分状况,经过5 a试验后,RGC处理和CK处理的土壤pH值均有所降低,但RGC处理降低趋势较CK显著(P<0.05)。有机质、全氮、碱解氮、速效磷、速效钾、紧实度和土壤容重均表现为RGC>CK。
2.2 玉米田养鹅模式下土壤呼吸日变化动态
由图1可知,CK土壤呼吸速率日变化特征呈“单峰型”变化态势,其中幼苗期、拔节期、抽雄期、开花期、吐丝期和成熟期土壤呼吸速率分别分布在0.59~1.86、0.69~2.40、1.02~2.79、1.28~2.50、1.92~3.14 μmol·m-2·s-1和1.09~2.83 μmol·m-2·s-1,各生长期最大值分别出现在16∶00、16∶00、17∶00、19∶00、18∶00和17∶00。幼苗期、抽雄期和吐丝期最小值出现时段一致,在06∶00—07∶00,拔节期和开花期最小值出现时段一致,在07∶00—08∶00,成熟期最小值出现在06∶00。RGC处理幼苗期、拔节期、抽雄期、开花期、吐丝期和成熟期土壤呼吸速率分别在1.34~2.40、1.75~3.54、2.26~3.20、2.65~4.33、2.39~4.21 μmol·m-2·s-1和2.38~4.13 μmol·m-2·s-1,日变化趋势与CK一致,但极值出现时间存在差异。
2.3 玉米田养鹅模式下不同生长期土壤呼吸变化特征
由图2可知,随着生长期的推进,RGC和CK土壤呼吸速率均呈先增加后降低的变化趋势,即从幼苗期(RGC,1.75 μmol·m-2·s-1,CK,1.16 μmol·m-2·s-1,下同)开始,土壤呼吸速率逐渐增加,直至吐丝期(RGC,3.32 μmol·m-2·s-1,CK,2.32 μmol·m-2·s-1)达到最大,随后开始降低,至成熟期时,RGC和CK土壤呼吸速率分别为3.11 μmol·m-2·s-1和2.03 μmol·m-2·s-1。
从幼苗期至成熟期,土壤呼吸速率均表现为RGC>CK,RGC较CK分别高出51.74%、80.02%、81.61%、91.84%、43.19%和53.46%。在幼苗期至成熟期,RGC变异系数介于5.17%~26.48%,CK变异系数分布在3.16%~36.81%,各生长期均表现为CK>RGC,说明CK处理土壤呼吸更容易受到外界环境变化的干扰。
2.4 玉米田养鹅模式下土壤温度、体积含水量变化特征
2.4.1 土壤温度 由图3可知,从幼苗期至成熟期,RGC土壤温度变化范围分别为9.33~27.24、11.65~23.33、12.66~24.13、11.84~25.87、13.43~29.01、13.17~28.84℃,CK土壤温度变化范围分别为9.97~26.64、11.24~27.01、12.67~23.62、11.98~24.08、13.38~31.92、13.14~31.85℃,呈单峰型日变化特征。土壤温度快速升温期主要在07∶00—15∶00,快速降温主要在16∶00—19∶00。将幼苗期至成熟期土壤温度平均,土壤平均温度均表现为CK>RGC,且CK较RGC分别高出了2.58%、3.83%、0.41%、2.60%、0.64%和3.07%。

图3 不同处理下土壤温度日变化特征Fig.3 Diurnal variation characteristics of soil temperature under different treatments
2.4.2 土壤体积含水量 由图4可知,土壤体积含水量呈单峰型日变化态势。在6个生长期内,RGC处理土壤体积含水量变化范围分别在0.15~0.29、0.14~0.25、0.19~0.26、0.19~0.27、0.16~0.26 m3·m-3和0.17~0.26 m3·m-3;CK土壤体积含水量变化范围分别在0.15~0.23、0.13~0.23、0.18~0.28、0.17~0.29、0.19~0.27 m3·m-3和0.17~0.27 m3·m-3。各生长期土壤体积含水量最大值一般出现在06∶00—07∶00,最小值一般出现在14∶00—17∶00。平均而言,整个生育期内土壤平均体积含水量均表现为RGC>CK,幼苗期至成熟期,RGC分别较CK高出了6.41%、3.34%、0.79%、2.67%、3.04%和1.81%。
2.5 玉米田养鹅模式下土壤呼吸对温度和体积含水量的响应
2.5.1 土壤呼吸对温度的响应 土壤温度与土壤呼吸速率二者之间的对应关系呈近椭圆形分布特征,由图5可知,土壤温度与土壤呼吸速率二者并不是保持同步变化,而是在温度变化幅度大的一段时间内,土壤呼吸速率变化明显滞后于温度变化。处理RGC的幼苗期,土壤温度最小和最大值分别出现在05∶00和15∶00,而土壤呼吸速率最小和最大值出现在06∶00和16∶00,可见,土壤呼吸速率滞后于土壤温度变化1 h;CK处理土壤温度最小值和最大值出现时间分别为06∶00和15∶00,而土壤呼吸最大值和最小值分别滞后于土壤温度变化1 h和2 h。其他各生长期两者的变化特征与幼苗期相似,均存在土壤呼吸滞后现象。
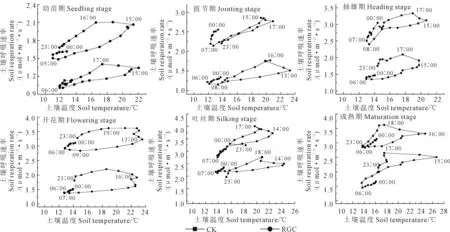
图5 不同处理下土壤呼吸速率对温度的响应Fig.5 Response of soil respiration rate to temperature under different treatments
2.5.2 土壤呼吸对土壤体积含水量的响应 土壤呼吸速率与土壤体积含水量的关系同样呈近椭圆形分布特征(图6),表现为随土壤体积含水量的增加土壤呼吸速率呈降低的趋势。RGC处理幼苗期、拔节期、抽雄期、开花期、吐丝期和成熟期的土壤呼吸速率最小值滞后于土壤体积含水量变化0、1、1、3、0 h和0 h,土壤呼吸速率最大值分别滞后于体积含水量变化1、0、0、3、2 h和1 h。CK幼苗期至成熟期,在最小值出现时间上土壤呼吸速率分别滞后于土壤体积含水量变化0、1、0、1、1 h和-1 h,在最大值出现时间上土壤呼吸速率分别滞后于土壤体积含水量变化0、1、0、4、4 h和1 h。

图6 不同处理下土壤呼吸速率对土壤体积含水量的响应Fig.6 Response of soil respiration rate to soil volumetric water content under different treatments
2.5.3 土壤呼吸速率与土壤温度、体积含水量的复合关系 土壤呼吸速率的变化并非受单一因子的影响,而是受多种因素的共同作用,其中,土壤温度和土壤体积含水量的影响尤为重要[11],且关系复杂。诸多研究者在进行回归分析时选择的关系拟合模型差异较大,笔者采用双变量回归模型y=at+bw+k(y为土壤呼吸速率,t为土壤温度,w为土壤体积含水量)对土壤呼吸速率与土壤温度及体积含水量的关系进行回归分析,结果见表2。CK幼苗期至成熟期,土壤呼吸速率与土壤温度及体积含水量之间均达到了极显著相关水平(P<0.01),土壤温度和土壤体积含水量两者拟合的双变量模型可共同解释土壤呼吸速率日变化的86.21%~98.12%;RGC处理幼苗期至成熟期,土壤呼吸速率与土壤温度及体积含水量之间亦均达到了极显著相关水平(P<0.01),土壤温度和体积含水量两者拟合的双变量模型可共同解释土壤呼吸速率日变化的58.32%~96.51%。相对于RGC处理,CK的土壤呼吸速率整体上更容易受到土壤温度和体积含水量变化的影响。可见,玉米田养鹅可能会通过提高土壤碳库的稳定性,从而降低土壤呼吸对外界环境温湿度变化的响应程度。

表2 不同处理下土壤呼吸与土壤温度、体积含水量的复合关系Table 2 Compound relation between soil respiration and temperature and volume water content under different treatments
土壤有机碳分解的温度敏感性(Q10)是指土壤温度每上升10℃,土壤有机碳分解速率所增加的倍数。Q10值越大,表明土壤有机碳分解对温度变化越敏感[8]。本研究中,各生长期CK温度敏感性系数Q10值分布在1.19~1.38,平均值为1.33;各生长期RGC温度敏感性系数Q10值分布在1.12~1.30,平均值为1.25,温度敏感性系数Q10值整体表现为:CK>RGC。其原因可能是鹅粪进入土壤改善了土壤结构,提高了土壤团聚体的稳定性,对土壤有机碳起到了保护作用,从而降低了土壤有机碳分解的温度敏感性。
3 讨 论
3.1 玉米田养鹅模式下土壤理化性状变化特征及影响因素分析
相对于CK而言,RGC处理可有效提高土壤养分含量,但会降低土壤pH值,增加土壤容重和紧实度,其原因可能与RGC处理具有丰富的杂草多样性、较高的生物量以及鹅以杂草和作物基部叶片为食,养分以粪便形式还田,从而使RGC处理模式具有更加良好的物质循环和养分反馈特性,但鹅的活动会增加土壤紧实度和土壤容重,同时,鹅粪为生理酸性肥,可使土壤酸化。综上可知,RGC处理模式可降低化肥投入,减小环境污染,同时,土壤养分含量明显增加,土壤肥力有效性提高,另外,土壤pH值的降低,有利于盐渍化土壤理化性状的改善。
各生长期土壤温度和体积含水量均呈“单峰型”日变化趋势,且整个生育期内,土壤体积含水量表现为RGC>CK,这主要可能是鹅产生的粪便增加了土壤有机质含量,改善了土壤物理性状,提高了土壤保水保肥性能,从而使得RGC处理的体积含水量高于CK。本研究中,土壤温度表现为CK>RGC,其原因可能在于RGC处理地上生物量(386.30 g·m-2)是CK(5.67 g·m-2)的68.13倍,盖度(90.56%)是CK(7.22%)的12.54倍[9],这可使得RGC处理能有效阻挡太阳辐射,同时,RGC处理较高的体积含水量和有机质含量使得其比热容大于CK,导致在同一大气环境条件下,RGC处理的温度变化(升温或降温)小于CK。可见,RGC模式可有效提高土壤保水能力,降低土壤温度。
3.2 玉米田养鹅模式下土壤呼吸变化特征及影响因素分析
本研究对RGC模式下土壤呼吸速率的研究表明,CK和RGC土壤呼吸速率均呈“单峰型”日变化特征,但不同测定日期土壤呼吸峰值出现时间存在差异,这与大多数研究结果[10-11]一致。其原因可能是不同测定季节的作物生长状况[12]、测定时段的天气变化特征[13]、样地土壤的类型和质地[14]、作物根系分布及生物量[15]、土壤生物组成及分布[16]等不同有关。同时,CK和RGC的田间管理措施和外源养分输入等存在差异,这也可导致在同一测定条件下土壤呼吸峰值出现时间上存在位差。
在整个生长期内,CK土壤呼吸速率峰值出现在吐丝期,其值变化范围在1.92~3.14 μmol·m-2·s-1,均值为2.32 μmol·m-2·s-1。RGC处理土壤呼吸速率峰值亦出现在吐丝期,其值变化范围在2.39~4.21 μmol·m-2·s-1,均值为3.32 μmol·m-2·s-1,RGC处理较CK提高了43.10%(P<0.05)。相对于CK而言,其一,RGC处理具有更高的地上地下生物量,其二,鹅的活动为土壤增加了额外的碳源和氮源,增强了土壤微生物活性[17],提升了土壤微生物呼吸的碳排放速率,同时,由于养分的额外输入,使得RGC较CK更能满足作物生长的养分需求,使植物的根系生物量、土壤养分、微生物多样性和酶活性均有提高[18-19],从而提高了土壤自养呼吸,表现为RGC处理的土壤呼吸整体高于CK。CK处理为单施化肥,而化肥的施用会降低微生物多样性、活性及碳源的利用效率[20],从而抑制了土壤异养呼吸的能力,降低了土壤呼吸速率。其他生长期平均土壤呼吸速率均表现为RGC>CK。
从土壤呼吸空间变异性的角度,RGC处理的土壤呼吸速率空间变异性远小于CK,这可能是因为长期的鹅粪输入土壤后改善了土壤理化性状,增加了土壤团聚体对有机碳的物理保护,从而使得RGC模式的土壤碳库相对较稳定,土壤呼吸速率的空间变异性小;另外,RGC模式长期的鹅粪输入增加了土壤微生物的营养来源,改善其生存环境,提高土壤微生物的种类和数量,使得其在外界环境变化时具有更高的抵抗能力,故使得RGC模式下的土壤呼吸速率变异性较小。
3.3 玉米田养鹅模式下土壤呼吸速率与土壤温度、体积含水量的关系
土壤呼吸速率的变化主要是受土壤温度和土壤体积含水量的协同影响,而非单一因子的作用[21]。本研究表明,CK和RGC处理的土壤温度和体积含水量均可显著影响土壤呼吸速率的变化,二者可共同解释土壤呼吸变化的86.21%~98.12%(CK)和58.32%~96.51%(RGC),可见,相对于CK,RGC处理可能会通过改善土壤养分结构及物理性质,从而减弱土壤呼吸对外界环境的响应程度。相关研究表明,在没有水分胁迫的条件下,土壤温度与土壤呼吸之间呈显著正相关关系[22],而土壤水分只有在最低或最高时才会对土壤呼吸速率产生影响[23]。本研究中,土壤呼吸速率随土壤温度的变化呈正相关近椭圆形分布关系,与土壤体积含水量之间呈负相关椭圆分布关系,但在极值出现时间上,土壤呼吸速率略滞后于土壤温度和体积含水量的变化。Vargas等[24]研究得出,光合作用是造成土壤呼吸与土壤温度日滞后现象的主要因素,其原因可能是温度的增加导致植物的蒸腾作用增加,植物叶片水分减少,限制了光合作用和减少了光合产物向根部的输送,同时,光合产物从叶片输送至根系需要一定的时间,导致了增温与根呼吸之间产生了位差,从而影响了根呼吸,导致了呼吸作用的滞后现象;另外,土壤热传递符合傅里叶热传导定律[25],表层土壤是连接地上与地下热辐射和气体交换的关键层,土壤呼吸动态变化优先受近地表温度的影响,而深层土壤呼吸受到增温的影响较小,导致表层和深层土壤呼吸速率之间出现位差[26],使得土壤呼吸速率滞后于温度的变化。
3.4 玉米田养鹅模式下土壤呼吸温度敏感性
研究者常用温度敏感性系数(Q10)来反映土壤温度和土壤呼吸速率之间的关系,Zheng等[27]研究表明,不同农业管理措施下土壤温度敏感性系数(Q10值)的变化范围分别在1.28~4.75和2.35~3.49。在本研究中,CK和RGC处理的Q10值变化范围分别为1.19~1.38和1.12~1.30,均小于以上研究结果,但本研究结果与Raich等[28]的研究范围相似(Q10值范围为1.3~3.3)。有研究表明,相对于难分解有机质而言,易分解有机质的Q10值相对较小[29],本研究中,RGC处理的Q10值整体小于CK处理,这主要是因为本研究区域农田土壤类型为沙壤土,沙粒含量较高,肥力较低,相对于CK而言,RGC处理通过鹅粪投入和杂草腐殖化[30],增加了土壤有机质含量,改善了土壤物理结构、提高了土壤胶结、保肥和缓冲能力[31],同时,RGC处理有鹅的活动,提高了土壤紧实度,降低了孔隙度,从而使得本研究中Q10值表现为RGC 本文基于全球气候变化及青藏高原高寒环境条件的双重背景下,研究高寒区域玉米田养鹅模式下土壤碳排放特征,所得结论如下: 1)相对于CK而言,RGC处理可有效提高土壤养分含量,但会降低土壤pH值,增加土壤容重和紧实度; 2)土壤温度、体积含水量和碳排放速率均表现为“单峰型”日变化特征,但在峰值和谷值出现时间上存在差异,即土壤呼吸速率滞后于土壤温度和含水量的变化。双变量拟合模型显示,土壤呼吸速率的日变化极显著响应土壤温度与体积含水量的协同变化(86.21% 3)整个生育期内,土壤碳排放速率均表现为RGC>CK,说明玉米田养鹅模式可有效提高土壤碳排放效率,但从空间变异性(变异系数CK>RGC)和温度敏感性系数(Q10值表现为CK(1.19~1.38)>RGC(1.12~1.30))来看,玉米田养鹅生产模式可有效提高土壤呼吸抵抗外界干扰的能力和降低高寒区域农田土壤温度敏感性; 4)本研究测定期间,土壤呼吸速率表现为RGC>CK,但变异系数表现为CK>RGC,说明CK的土壤呼吸更容易受到外界环境变化的影响。因此,在高寒区域农田进行玉米田养鹅模式,有利于提高土壤肥力、增加土壤碳稳定性,降低土壤碳排放速率温度敏感性,这对高寒环境条件下玉米田养鹅模式的进行及土壤固碳减排具有一定指导意义。4 结 论
