微小RNA-16对高糖诱导人视网膜内皮细胞凋亡的抑制作用
2022-01-27刘志涛刘刚陈霞王岁月
刘志涛,刘刚,陈霞,王岁月
糖尿病性视网膜病是成年人失明的主要原因[1-2],据报道,60%以上的2型糖尿病病人在发病数年后出现视网膜病变[3]。目前,尽管相关学者已经就导致糖尿病引起的视网膜损伤的潜在原因进行了大量工作,但糖尿病性视网膜病的机制仍未完全了解。众所周知,高血糖是导致糖尿病性视网膜病发作和发展的关键危险因素。慢性高血糖症会损害多种细胞类型,并导致包括眼睛在内的许多器官功能障碍和衰竭[4]。对高葡萄糖相关的病理学途径的了解越来越重要,了解高葡萄糖反应相关细胞途径对于糖尿病性视网膜病治疗至关重要。
微小RNA(microRNA,21至23个核苷酸的小的非编码RNA小分子)作为基因表达的新兴关键调控因子在调节多种细胞活动(包括细胞发育、增殖、分化和凋亡)方面具有重要作用[5-6]。目前,部分miRNA已经作为诊断某些疾病的潜在生物标志物进行临床应用[7-9]。miRNA表达及其功能具有组织和细胞特异性。糖尿病与胰岛素靶组织以及胰岛素分泌细胞中几种miRNA的表达改变有关。其他研究发现 miR-19a、miR-1740、miR-126、miR-200等 miRNA在糖尿病性视网膜病变中发生异常表达[10-14]。然而,迄今为止,只有很少的研究考察了miRNA的下游细胞信号传导,且关于miRNA如何影响糖尿病性视网膜病常见的病理机制尚未完全揭示。最近,有研究发现在人脐静脉内皮细胞(HUVEC)中miR-16的过表达在抑制血管生成中发挥重要作用[15]。另一项研究发现miR-16在链脲佐菌素(STZ)诱导的糖尿病大鼠视网膜内皮细胞(REC)中表达增加[11]。然而,关于miR-16在人视网膜内皮细胞中的表达及其在与糖尿病性视网膜病发病机制相关的下游细胞信号传导中的潜在作用知之甚少。
其他研究报道,在高葡萄糖条件下,视网膜内皮细胞中的肿瘤坏死因子-α(TNF-α)水平升高。TNF-α可激活细胞因子信号转导负调控因子(SOCS3)信号,以及胰岛素受体底物1(IRS-1)和胰岛素受体(IR)的磷酸化,导致在高葡萄糖条件下正常胰岛素信号传导的抑制[16]。本研究自2019年2―8月考察miR-16在高糖诱导人视网膜血管内皮细胞凋亡中的分子机制。
1 材料与方法
1.1 细胞培养人视网膜内皮细胞购自美国ATCC。细胞在含有微血管生长补充剂、10 μg/mL庆大霉素和0.25 μg/mL两性霉素B的M131培养基中培养(美国Invitrogen公司)。将细胞在正常葡萄糖培养基(5 mmol/L)或高葡萄糖培养基(30 mmol/L)(美国Invitrogen公司)中培养3 d。通过在无生长补充剂的高葡萄糖或正常葡萄糖培养基中孵育20 h使细胞静止并用于后续实验。
1.2 microRNA模拟物转染细胞按照制造商的说明,使用Oligofectamine(美国Invitrogen公司)将miRNA模拟物[miR-16-mimic,生工生物工程(上海)股份有限公司合成]转染视网膜内皮细胞,终浓度为50 nmol/L。用等浓度的模拟物阴性对照(miRNC)处理阴性对照组。通过实时荧光定量逆转录聚合酶链式反应(qRT-PCR)验证miRNA表达水平。
1.3 qRT-PCR使用TRIzol方法(美国Invitrogen公司)分离和纯化总RNA。对于polyA尾法逆转录酶PCR,将5 μg总RNA在室温下用DNase I[中科瑞泰(北京)生物科技有限公司]处理15 min,然后使用polyA聚合酶[生工生物工程(上海)股份有限公司]在37℃下处理1 h。最终的反应混合物用苯酚/氯仿萃取,异丙醇沉淀,然后重新溶于25 μL焦碳酸二乙酯(DEPC)处理的水中。使用Superscript Ⅱ逆转录酶(美国Invitrogen公司)和oligo-dT锚定引物5'GCGACGCAATAACTCTGGTTTGACTTAATTTAGTATTTAACTTTVN3'将 PolyA 尾 RNA(6 μL)逆转录成第一链cDNA。对于PCR,使用SYBR Green I在LightCycler 480实时PCR系统(美国Roche Applied Science公司)上进行PCR。反应中使用的引物序列如下 :miR-16,正向 :CAAAAAGCGGCTACTTTAAGCACTC,反向 :TCGAAGTTACACCGGTTCCAGAAGGA;U6,正向:GCTTCAGCTCATGGGATTCCAAC,反向:TGCATCCCTCATGTAGCTTGGG。U6用作内源对照。采用比较阈值循环(2−ΔΔCt)方法计算mRNA相对表达量。
1.4 蛋白质印迹法(Western blotting)分析用冷PBS冲洗细胞后,将视网膜内皮细胞收集在含有蛋白酶和磷酸酶抑制剂的裂解缓冲液中(上海碧云天生物技术有限公司)。在预制的tris-甘氨酸凝胶(美国Invitrogen公司)上分离等量的蛋白质,然后将其印迹到硝酸纤维素膜上。在TBST(10 mmol/L Tris-HCl缓冲液、pH 8.0、150 mmol/L NaCl、0.1%Tween 20)和5%(W/V)BSA中封闭1 h后,将膜用相应的一抗在4℃过夜孵育,然后用辣根过氧化物酶标记的二抗室温孵育1 h。通过增强的化学发光试剂盒(Cell Signaling Technology公司)检测免疫条带。GAPDH作为内部对照。使用的一抗包括TNF-α、胰岛素受体底物-1丝氨酸307(IRS-1 Ser307)、磷酸化的 IRS-1 Ser307、SOCS3、胰岛素受体 Tyr1150(IR Tyr1150)、磷酸化的IR Tyr1150、蛋白激酶B(Akt)、磷酸化的Akt、活化的胱天蛋白酶-3(cleaved caspase 3)和1-甘油醛-3-磷酸脱氢酶(GAPDH)。所有抗体全部购自美国Cell Signaling Technology公司。
1.5 统计学方法使用SPSS 17.0软件进行统计分析。数据以±s表示。使用Studentt检验进行两组间数据比较,实验采用单因素方差分析和LSD法进行多组间数据比较。P<0.05表示差异有统计学意义。
2 结果
2.1 高糖抑制视网膜内皮细胞中miR-16的表达水平本研究检测了高糖培养基培养的视网膜内皮细胞中miR-16表达的变化。通过在高葡萄糖培养基(25 mmol/L)中培养视网膜内皮细胞,并从细胞中分离出总RNA,然后进行qRT-PCR。qRT-PCR结果显示,与正常葡萄糖组相比,高葡萄糖培养可显著降低miR-16的水平[对照组(1.07±0.08)比高糖组(0.35±0.03),t=14.43,P=0.011]。由于高葡萄糖导致miR-16的表达降低,本研究通过对视网膜内皮细胞转染miR-16模拟物(50 nmol/L)48 h来增加miR-16的表达。qRT-PCR证实了转染后细胞内miR-16的表达显著增加[高糖+miR-NC组(1.01±0.07)比高糖+miR-16组(3.54±0.27),t=18.55,P=0.004]。
2.2 上调miR-16降低视网膜内皮细胞中的TNF-α水平及IRS-1 Ser307的磷酸化为了确定miRNA-16是否参与胰岛素信号传导,本研究考察了改变miRNA-16的表达对糖尿病性视网膜病变相关下游信号通路的影响。其他研究者已经证明TNF-α水平在糖尿病性视网膜病变中明显升高[16]。本研究发现,与对照相比,用miR-16转染的视网膜内皮细胞的TNF-α水平显著降低(图1A和表1)。因此,miR-16的过表达可降低高葡萄糖诱导的视网膜内皮细胞中的TNF-α水平。另外,敲低TNF-α会导致IRS-1 Ser307的磷酸化降低,从而促进正常的胰岛素信号转导[16]。在本研究中,高葡萄糖条件下培养的细胞中的IRS-1磷酸化水平明显增加,而上调miRNA-16可抑制IRS-1 Ser307的磷酸化(图1B和表1)。这表明miR-16可能在视网膜内皮细胞中通过抑制TNF-α来抑制IRS-1Ser307的磷酸化。

图1 上调miR-16降低视网膜内皮细胞中的蛋白磷酸化:A为TNF-α;B为胰岛素受体底物-1(IRS-1)Ser307
表1 各组肿瘤坏死因子-α(TNF-α)蛋白相对表达量和胰岛素受体底物-1(IRS-1)磷酸化情况/±s
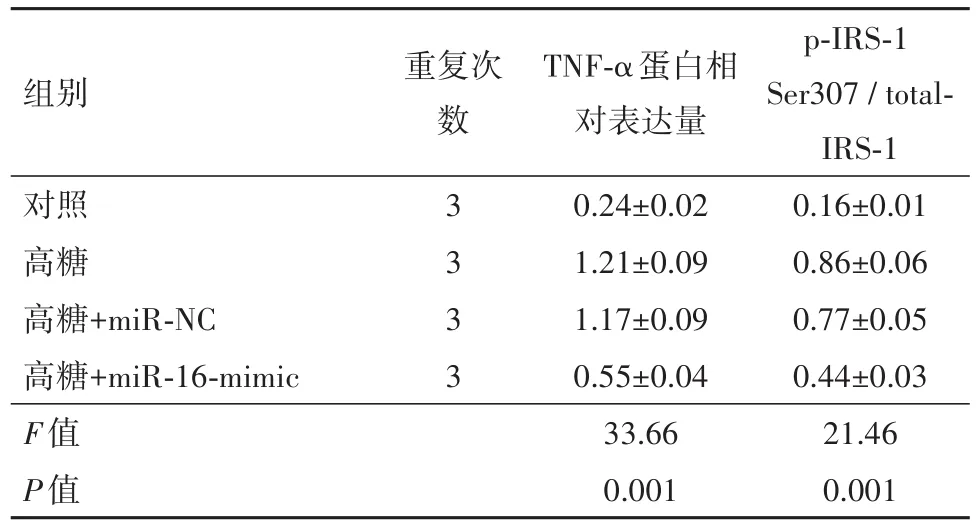
表1 各组肿瘤坏死因子-α(TNF-α)蛋白相对表达量和胰岛素受体底物-1(IRS-1)磷酸化情况/±s
组别 重复次对照高糖高糖+miR-NC高糖+miR-16-mimic F值P值数 3 3 3 3 TNF-α蛋白相对表达量0.24±0.02 1.21±0.09 1.17±0.09 0.55±0.04 33.66 0.001 p-IRS-1 Ser307/total-IRS-1 0.16±0.01 0.86±0.06 0.77±0.05 0.44±0.03 21.46 0.001
2.3 上调miR-16降低视网膜内皮细胞中的SOCS3水平前人研究发现,TNF-α可激活视网膜内皮细胞中的SOCS3信号[16]。本研究考察了miR-16是否会改变高葡萄糖诱导的视网膜内皮细胞中的SOCS3水平。Western blotting数据表明,高葡萄糖处理显著增加了视网膜内皮细胞中的SOCS3水平,而miR-16的过表达则显著降低了SOCS3的水平(图2和表2)。因此miR-16在高葡萄糖诱导的视网膜内皮细胞中抑制SOCS3信号传导。

图2 上调miR-16降低视网膜内皮细胞中的细胞因子信号转导负调控因子(SOCS3)水平
表2 各组细胞因子信号转导负调控因子(SOCS3)蛋白相对表达量/±s
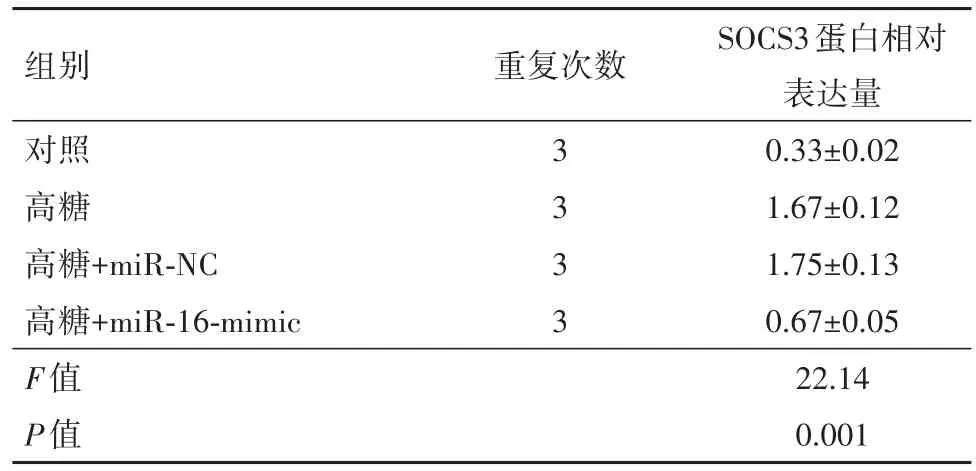
表2 各组细胞因子信号转导负调控因子(SOCS3)蛋白相对表达量/±s
组别对照高糖高糖+miR-NC高糖+miR-16-mimic F值P值重复次数3 3 3 3 SOCS3蛋白相对表达量0.33±0.02 1.67±0.12 1.75±0.13 0.67±0.05 22.14 0.001
2.4 上调miR-16增强视网膜内皮细胞中IR Tyr1150的磷酸化高血糖症中SOCS3和TNF-α的增加可通过促进IRS-1 Ser307磷酸化并抑制IR Tyr1150的磷酸化来抑制正常胰岛素信号[17]。在本研究中,高葡萄糖条件下培养的视网膜内皮细胞中IRTyr1150磷酸化水平明显降低,上调miR-16则会提高IRTyr1150磷酸化水平(图3和表3)。结果表明,胰岛素受体是视网膜内皮细胞中受miR-16调节的靶途径之一,而高水平的miR-16可在高血糖症中保护视网膜内皮细胞。

图3 上调miR-16增强视网膜内皮细胞中胰岛素受体Tyr1150(IR Tyr1150)的磷酸化
表3 各组胰岛素受体Tyr1150(IR Tyr1150)磷酸化/±s

表3 各组胰岛素受体Tyr1150(IR Tyr1150)磷酸化/±s
组别对照高糖高糖+miR-NC高糖+miR-16-mimic F值P值重复次数3 3 3 3 p-IR Tyr1150/total-IR Tyr1150 1.01±0.07 0.32±0.02 0.35±0.02 0.92±0.07 16.78 0.003
2.5 上调miR-16在视网膜内皮细胞中增加Akt磷酸化并减少细胞凋亡本研究考察了miR-16的上调对视网膜内皮细胞中Akt的磷酸化的影响。研究发现,高葡萄糖培养降低了Akt磷酸化,而miR-16模拟物处理则可促进Akt的磷酸化(图4A和表4)。

图4 上调miR-16在视网膜内皮细胞中增加Akt磷酸化(A)并减少cleaved caspase 3(B)的水平
表4 各组的蛋白激酶B(Akt)磷酸化和活化的胱天蛋白酶-3(cleaved caspase 3)蛋白相对表达量/±s
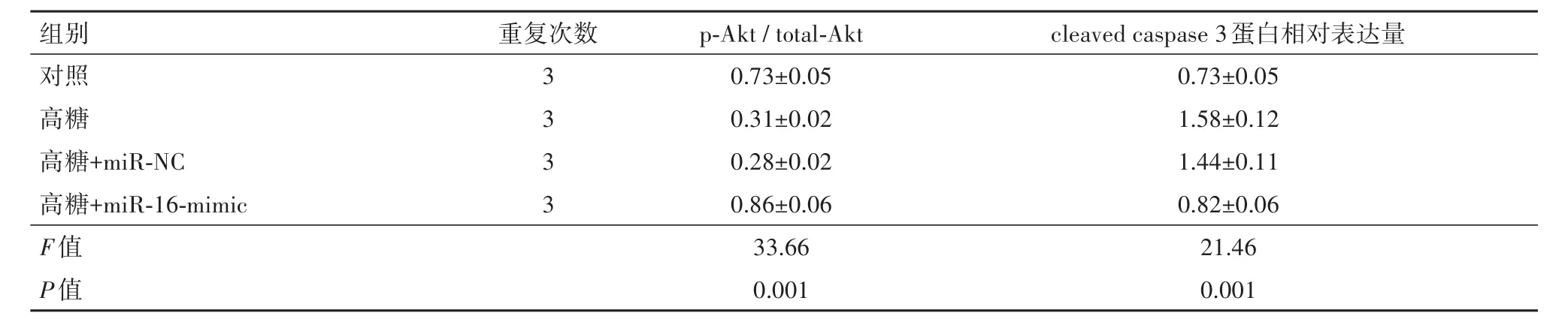
表4 各组的蛋白激酶B(Akt)磷酸化和活化的胱天蛋白酶-3(cleaved caspase 3)蛋白相对表达量/±s
组别 重复次数p-Akt/total-Akt cleaved caspase 3蛋白相对表达量0.73±0.05 0.31±0.02 0.28±0.02 0.86±0.06 33.66 0.001对照高糖高糖+miR-NC高糖+miR-16-mimic F值P值3 3 3 3 0.73±0.05 1.58±0.12 1.44±0.11 0.82±0.06 21.46 0.001
此外,miR-16的过表达显著降低了高葡萄糖中cleaved caspase 3的水平(图4B和表4)。这些结果表明,miR-16可以通过激活Akt信号途径来抑制高葡萄糖诱导的视网膜内皮细胞凋亡。
3 讨论
微血管修饰是糖尿病性视网膜病变的重要改变之一。其他工作者已经证明,视网膜内皮细胞是在糖尿病性视网膜病中受到实质影响的关键细胞类型之一[13,18],但尚不清楚糖尿病性视网膜病的潜在细胞机制。糖尿病视网膜病变的新潜在因素是miRNA的调节。其他研究表明miR-200b在高葡萄糖条件处理的HUVEC中被下调[19]。此外,高糖可通过降低HUVEC中miR-146a的表达来调节糖尿病中细胞外基质蛋白的产生[20]。本研究旨在探讨miR-16对高葡萄糖引起的人类视网膜内皮细胞损伤的影响。据报道,miR-16在多种哺乳动物组织中表达,包括脑、心脏、骨骼肌、肝、肺、肾、脾脏等[21-23]。本研究证明了miR-16可在人视网膜内皮细胞中表达,并且高糖处理可抑制人类视网膜内皮细胞中miRNA的水平。
其他研究者发现,高葡萄糖会增加视网膜内皮细胞中的TNF-α和SOCS3蛋白水平,TNF-α和SOCS3水平的升高会导致IRS-1 Ser307磷酸化增强,以及IR Tyr115磷酸化降低,从而导致胰岛素抵抗和凋亡增加[16-17]。敲低 TNF-α会导致 IRS-1 Ser307的磷酸化降低,从而促进正常的胰岛素信号转导。此外,上调TNF-α可激活视网膜内皮细胞中的SOCS3信号[16]。miR及其靶标之间的调控网络很复杂,因为单个miRNA可以与数百个靶基因结合,而一个靶基因可以被多个miRNA调控[24]。各种miRNA靶向不同水平的胰岛素信号传导的分子。不同的miRNA参与信号蛋白的表达,例如胰岛素、IRS-1 和 IRS-2和 Akt[25]。本研究证实视网膜内皮细胞中miR-16的水平升高可抑制TNF-α和SOCS3的信号转导。此外,上调miRNA-16可抑制IRS-1 Ser307的磷酸化并促进IR Tyr 1150的磷酸化。这表明miR-16可能在视网膜内皮细胞中通过抑制TNF-α和SOCS3信号通路来调节下游信号分子如IRS-1 Ser307和IR Tyr 1150的表达,从而发挥视网膜内皮细胞保护作用。
据报道,TNF-α水平降低可导致Akt磷酸化增加,从而抑制细胞凋亡[26]。TNF-α水平升高导致cleaved caspase 3水平升高并引起细胞凋亡[27]。在高葡萄糖情况下,视网膜内皮细胞中Akt磷酸化水平的降低会显著增加促凋亡蛋白如cleaved caspase 3的表达[16]。因此,本研究考察了miR-16的上调是否可以增加视网膜内皮细胞中Akt的磷酸化。研究发现,高葡萄糖培养降低了Akt磷酸化,而miR-16模拟物处理则可促进Akt的磷酸化。此外,miR-16的过表达显著降低了高葡萄糖中cleaved caspase 3的水平。这些结果表明,miR-16可以通过激活Akt信号途径来抑制高葡萄糖诱导的视网膜内皮细胞凋亡。
综上所述,本研究表明,miR-16通过抑制TNF-α和SOCS3信号通路来抑制IRS-1 Ser307的磷酸化并促进IRTyr1150和Akt的磷酸化,从而在抑制胰岛素抵抗中发挥作用,并包括视网膜内皮细胞保护免受高葡萄糖诱导的细胞凋亡。这一结果表明,miR-16是糖尿病性视网膜病的潜在治疗靶标。
