现行黄河口保护区典型湿地植被时空动态遥感监测
2022-01-22张晨宇陈沈良李鹏刘清兰
张晨宇,陈沈良*,李鹏,刘清兰
( 1. 华东师范大学 河口海岸学国家重点实验室,上海 200241)
1 引言
湿地是地球上重要的自然生态系统,具有蓄水调洪、调节气候、保持水土、保护生物多样性等重要作用[1-2]。其中滨海湿地处于海陆相互作用的特殊地带,生态和环境条件十分复杂,是最为脆弱和最具经济价值的生态系统之一[3]。滨海湿地生长有多种水生、沼生和耐盐植物群落,这些植被是滨海湿地生态系统的重要组成部分,也是表征生态系统健康状况的重要指标[4]。黄河三角洲湿地是世界上暖温带保存最广阔、最完善、最年轻的湿地生态系统。然而,受入海水沙和海洋动力的综合作用,黄河口滨海湿地处于动态变化中,生态系统不稳定,极易受到各种因素的破坏[5-6]。此外,黄河三角洲滨海湿地分布着大量的芦苇(Phragmites australis)、碱蓬(Suaeda salsa)和互花米草(Spartina alterniflora)等盐沼植被,这些湿地植被多处于水陆交界地带和半水生、水生环境下。植被混合交错分布且界线模糊,在遥感影像上因多种植物光谱混合而造成的“异物同谱”现象较为明显,对其进行精确分类的难度较大[7]。因此,采用先进的技术手段准确快速地提取湿地植被信息,监测湿地植被的长时序动态变化,对于生态保护和管理具有重要的科学价值和现实意义。
湿地野外调查存在很多困难及不确定性,而遥感技术具有覆盖范围广、获取速度快、信息量大、动态性强等优势[8-9],可为湿地研究提供强有力的技术支持。在过去10年中,高分辨率影像如Worldview-2/3、SPOT-5/6和GF-1/2等已被用于提取湿地植被信息[10-12],然而这些影像成本较高且无法实现长时序动态监测。具有中等分辨率的Landsat影像为湿地研究提供了长期连续的数据支撑,在大范围、长时序湿地植被监测等方面发挥着越来越重要的作用[13-15]。近年来,已有大量学者采用基于像素的方法对盐沼湿地植被开展信息提取和识别,如最大似然分类[16]、人工神经网络[17]、决策树[18]、支持向量机[19]和随机森林[20]等,分类精度达75%~90%。此外,基于对象的分类方法也被广泛地应用于湿地植被解译,毕恺艺等[21]基于Sentinel-2A时序数据利用面向对象决策树方法提取了陈家湾流域的植被信息,分类总体精度达到89.7%。Lantz和Wang[22]比较了基于对象和基于像素的最大似然分类,发现基于对象的方法具有更高的分类精度。在黄河口湿地植被监测方面,Cao等[23]基于单时相SPOT影像应用面向对象方法对2005年黄河三角洲湿地植被信息进行提取,总体分类精度为73.6%。徐洋[24]同样也基于面向对象方法利用2018年Sentinel-2A影像进行黄河口湿地植物分类,总体精度仅在63%左右。杨俊芳等[25]利用国产高分影像对2015年黄河三角洲互花米草分布情况进行了统计和分析。刘康等[26]采用监督分类的方法提取了1984-2014年期间4景黄河三角洲影像中碱蓬盐沼的信息,分类精度在80%~90%之间。
总的来看,现有针对黄河三角洲湿地植被信息提取的研究大多基于单时相遥感影像,分类精度有待提高。并且湿地植被制图的时间序列较短,难以准确反映植被长期的动态演变过程。因此,本文基于长时序Landsat卫星数据,结合植被的物候特征,采用面向对象和随机森林模型结合的方法对湿地植被进行分类,以期提高现有相关研究的制图精度,并分析近20年来现行黄河口保护区3种典型湿地植被(芦苇、盐地碱蓬和互花米草)的时空变化特征。
2 研究区与数据
2.1 研究区概况
现代黄河三角洲是中国三大河口三角洲之一,也是中国沿海地区年度新增土地面积最大的三角洲[27]。黄河三角洲国家级自然保护区成立于1992年,植被覆盖度为53.7%,是中国沿海植被覆盖度最大的地区[28]。该保护区分为现行黄河口保护区和刁口河口保护区两部分。本研究选择现行黄河口保护区内的自然滨海湿地作为研究区(图1),该区域包含的湿地类型主要为水体、光滩、草本沼泽和灌木沼泽,其中优势物种主要有芦苇、碱蓬和互花米草,且这3种植被的物候特征存在差异。具体表现为芦苇在5月初开始绿化并快速生长,而其他两种植被群落还没有开始生长;夏季生长旺季时,芦苇群落和互花米草群落的反射光谱特征差异不大,但到10月,芦苇的生长期已经基本结束,而互花米草的生物量达到最大;此外,由于9月土壤中的含盐量增加,此时盐地碱蓬呈现深红色状态[29]。
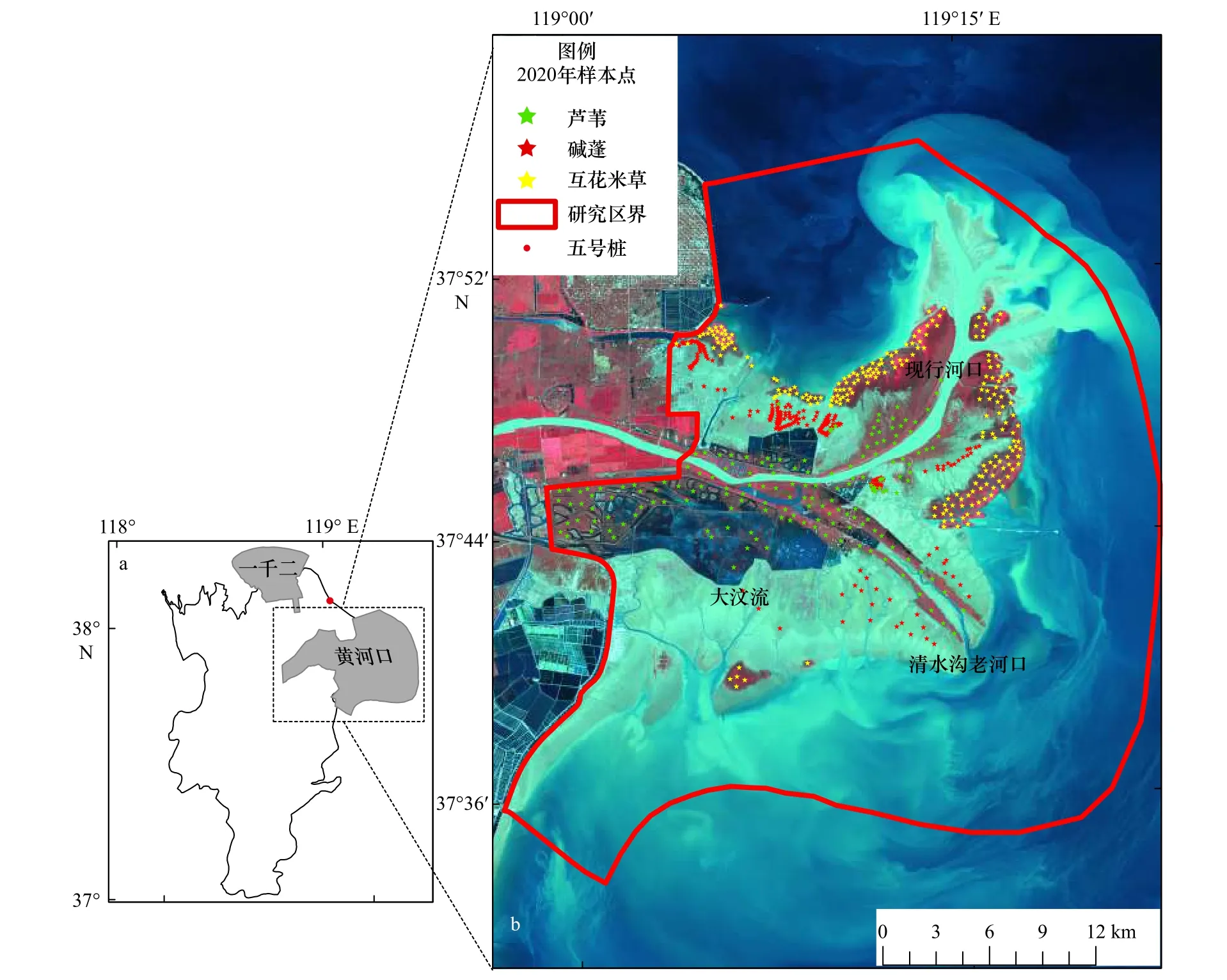
图1 研究区位置Fig. 1 Location of the study area
2.2 数据源
2.2.1 遥感数据
根据3种典型湿地植被的物候特征差异,可以判定黄河三角洲地区5月、9月和10月分别是提取芦苇、碱蓬和互花米草群落的重要时期。因此,为了准确提取湿地植被信息并进行长时间序列的监测,本文在2000-2020年期间每隔1年选择3景对应月份成像的Landsat系列影像,共33景(表1)。并使用ENVI软件对影像进行辐射定标和FLAASH大气校正等预处理操作。
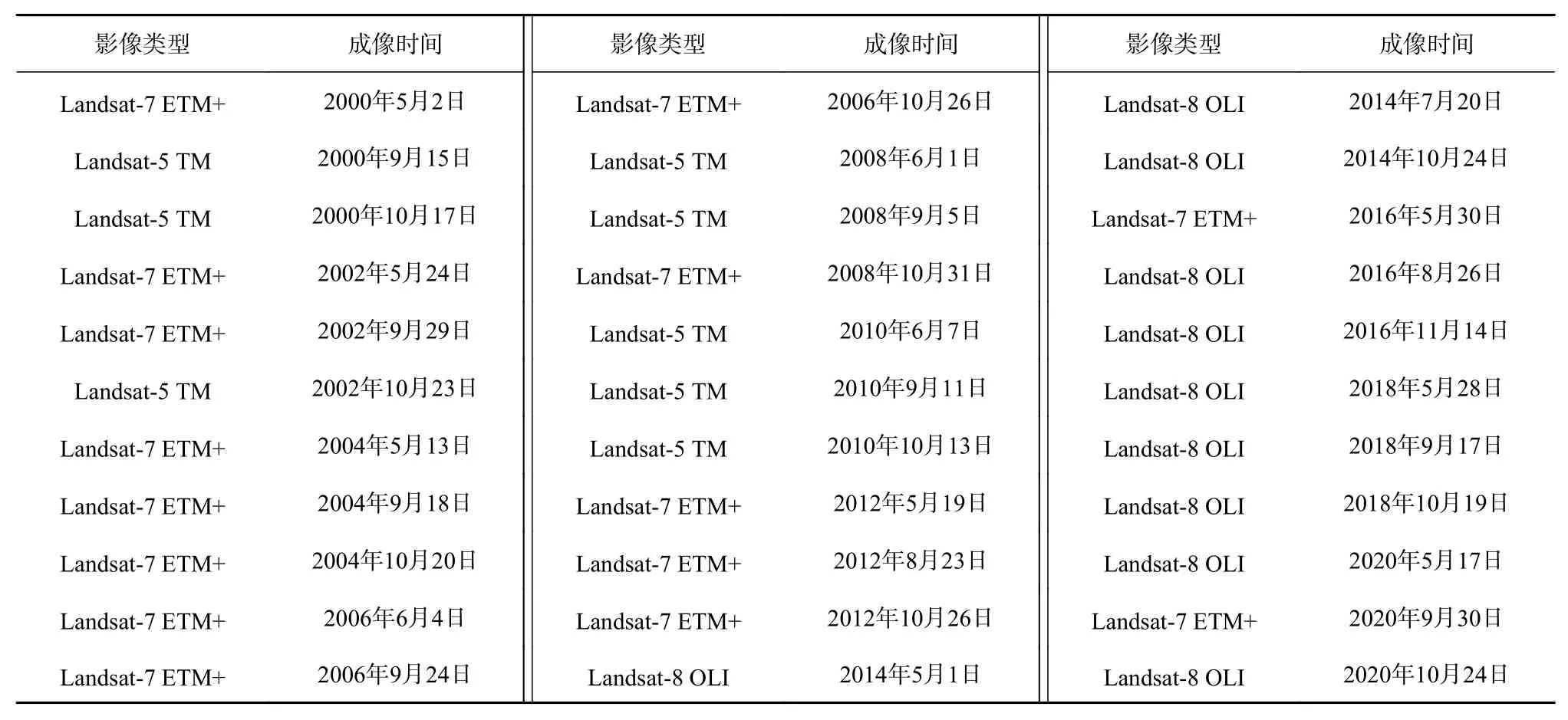
表1 卫星数据信息Table 1 Satellite data information
2.2.2 样本与验证数据
课题组于2020年9月在现行黄河口保护区开展实地考察调研,共获取248个样本观测数据(芦苇22个、碱蓬196个、互花米草12个、光滩15个、柽柳3个)。此外,还通过Google Earth、天地图等途径采集历史年份高分辨率遥感影像,根据各地物的遥感解译标志,同时考虑Landsat影像提供的物候差异信息,通过目视解译建立各历史年份的样本点。以其中2/3的样本点作为参考,在最佳分割结果的基础上选择分割对象单元作为训练样本;剩余作为验证样本,进行分类后精度验证。研究区2000-2020年各期训练样本和验证样本的数量分布见表2。
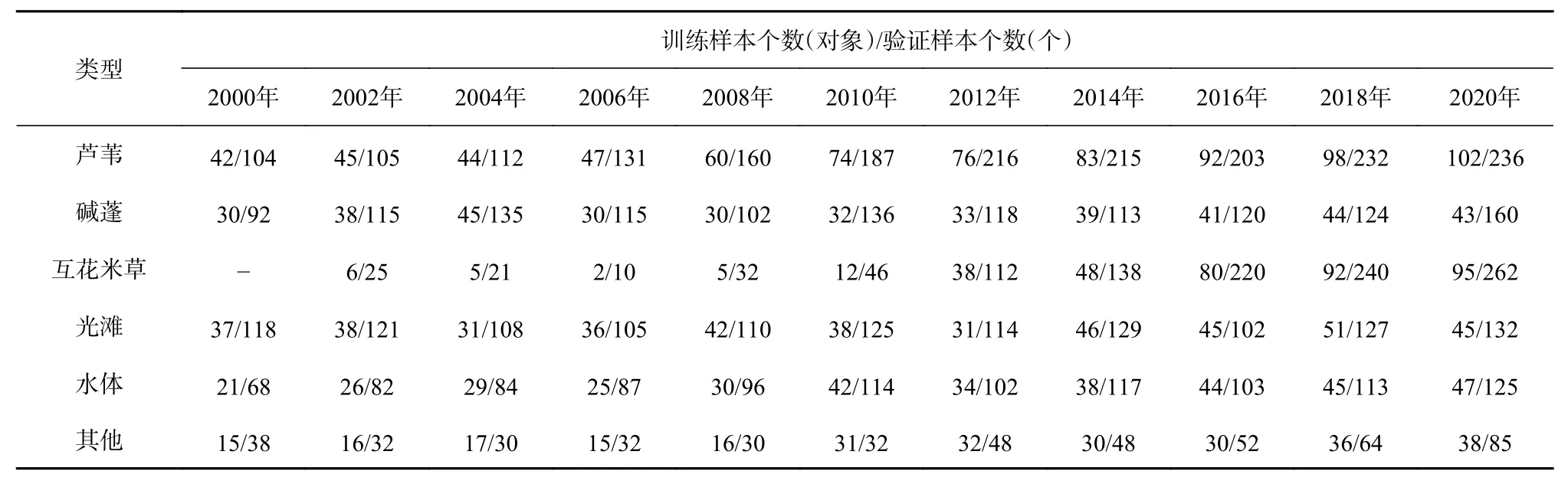
表2 2000-2020年不同地物类型的样本信息Table 2 Sample information of different feature types from 2000 to 2020
3 研究方法
3.1 面向对象分析
影像分割是面向对象提取植被信息的一个重要步骤,本文采用多尺度分割算法对遥感影像进行分割。它是一种基于异质性准则合并相邻像素或区域的分割算法[30]。对影像应用多尺度分割算法时,有3个重要的分割参数:形状因子、紧致度因子、分割尺度参数。形状因子越小,分割对象越复杂;紧致度因子越小,生成对象的边缘越平滑。本文在进行影像多尺度分割时,形状因子和紧致度因子参数的设置参考了Moffett和Gorelick[31]关于图像分割的研究结果,即Landsat影像设置形状因子/紧致因子为0.3/0.5时能更好地识别不同湿地植被斑块。
适当的分割尺度参数是获得最优分类结果的基础,其值越大,内部异质性越高,每个对象的像素数量也会增加[32]。因此,客观选择最优分割参数可提高分类的精度。本文利用基于eCognition Developer二次开发的ESP2尺度评价工具来确定最优分割尺度。该工具通过计算每个影像层对象异质性的局部变化(Local Variance,LV)作为分割对象层的平均标准差,ROC-LV表示从一个对象层到另一个对象层的LV变化率值,ROC呈现波峰状态时对应的尺度值即最优分割尺度[33]。结合ROC-LV曲线特征,并考虑到提取湿地植被信息的分割尺度不宜过大,且保护区内碱蓬斑块呈破碎化,经过反复测试,选择合适的分割尺度对影像进行分割。
3.2 随机森林算法及特征空间优化
随机森林算法是一种基于决策树的新型机器学习算法,随机抽取原始训练集2/3的样本作为一个新的训练集,然后从M个特征中随机抽取m个以完全分裂的方式生产决策树,最后集成多棵决策树组成随机森林分类器对特征数据集进行分类,新样本的类别由所有决策树以投票的方式来决定,每次抽样未被抽到的1/3数据称为袋外样本(Out of Bag,OOB),利用OOB数据通过交叉验证的方法可以估计内部误差[34]。
在多时相光谱特征的基础上加入植被指数和水体指数有利于提高分类精度。此外,纹理特征可以反映丰富的地物信息,几何特征能更好地识别具有明显形状特征的地物[35]。因此,本文选取了对象的光谱特征变量、植被指数、水体指数、纹理特征和几何特征来构建特征数据集(表3)。采用德国环境制图与分析计划项目组开发的EnMAP-BOX软件来优化特征空间及进行湿地植被信息提取。本文选取总特征个数的算术平方根作为抽取的特征变量个数;并通过实验发现决策树个数达到20时,Landsat数据的总体精度达到最大,因此本文选取N为20作为集成决策树的数量。随机森林算法可以对特征空间进行优化,通过计算特征变量的重要性得分,先将重要性得分最高的特征作为所选特征;然后按得分从高到低的顺序依次增加1个特征,计算每一次分类结果的精度,选出分类精度最高的一组作为最优特征组合。经过多次实验,本文选择了重要性较高的前26个特征变量组合的数据集进行分类,不同年份优选出的26个特征变量不完全相同。以2020年为例,最优特征数据集如表4所示。

表3 初始特征空间Table 3 The initial feature space

表4 优选特征分布Table 4 Preferred feature distribution
3.3 景观格局指数
本文基于类型水平,选取了斑块面积、斑块所占景观面积比例、最大斑块指数、斑块形状指数、平均斑块面积、聚合指数6种景观指数来表征3种典型湿地植被的景观格局演变特征,如表5所示。

表5 景观格局指数及其生态意义Table 5 Landscape pattern index and its ecological significance
4 结果与讨论
4.1 精度验证
本文利用混淆矩阵对2000-2020年的分类结果进行精度评价。从表6可以看出2000-2020年3种典型湿地植被的平均分类精度均达到80%以上,其中芦苇的分类精度最高,互花米草次之。而碱蓬的分类精度一般低于芦苇和互花米草,这是由于盐地碱蓬的植株高度和植株密度较低,其光谱特征受到土壤背景的强烈影响,很容易与光滩混淆[36]。此外,多年分类结果的总体精度处于84.74%~92.39%之间,Kappa系数处于0.81~0.91之间。可见本文基于多时相遥感影像结合面向对象和随机森林算法可以得到较高的分类精度,满足了湿地植被时空动态分析的需求。

表6 精度验证结果Table 6 Results of accuracy verification
4.2 近20年来3种典型湿地植被时空变化特征
图2为基于多时相Landsat系列影像提取的2000-2020年研究区典型湿地植被的空间分布。总体来看,芦苇主要呈条带状分布在黄河河道两侧及人工恢复区,受黄河淡水的调控,沿河道向现行河口新生湿地方向发展;碱蓬主要分布在研究区的滨海滩涂,时空分布变化较大。在现行黄河口以北和清水沟老河口北部的高潮滩地区由于土壤盐度适中,碱蓬呈大片分布。而大汶流区域由于盐度高于碱蓬适宜生态阈值,因此碱蓬生物量很小。互花米草主要生长在潮间带的中下部,在2002年时仅小片分布,2008年之后开始暴发式增长,逐步向陆和向新生湿地方向发展。
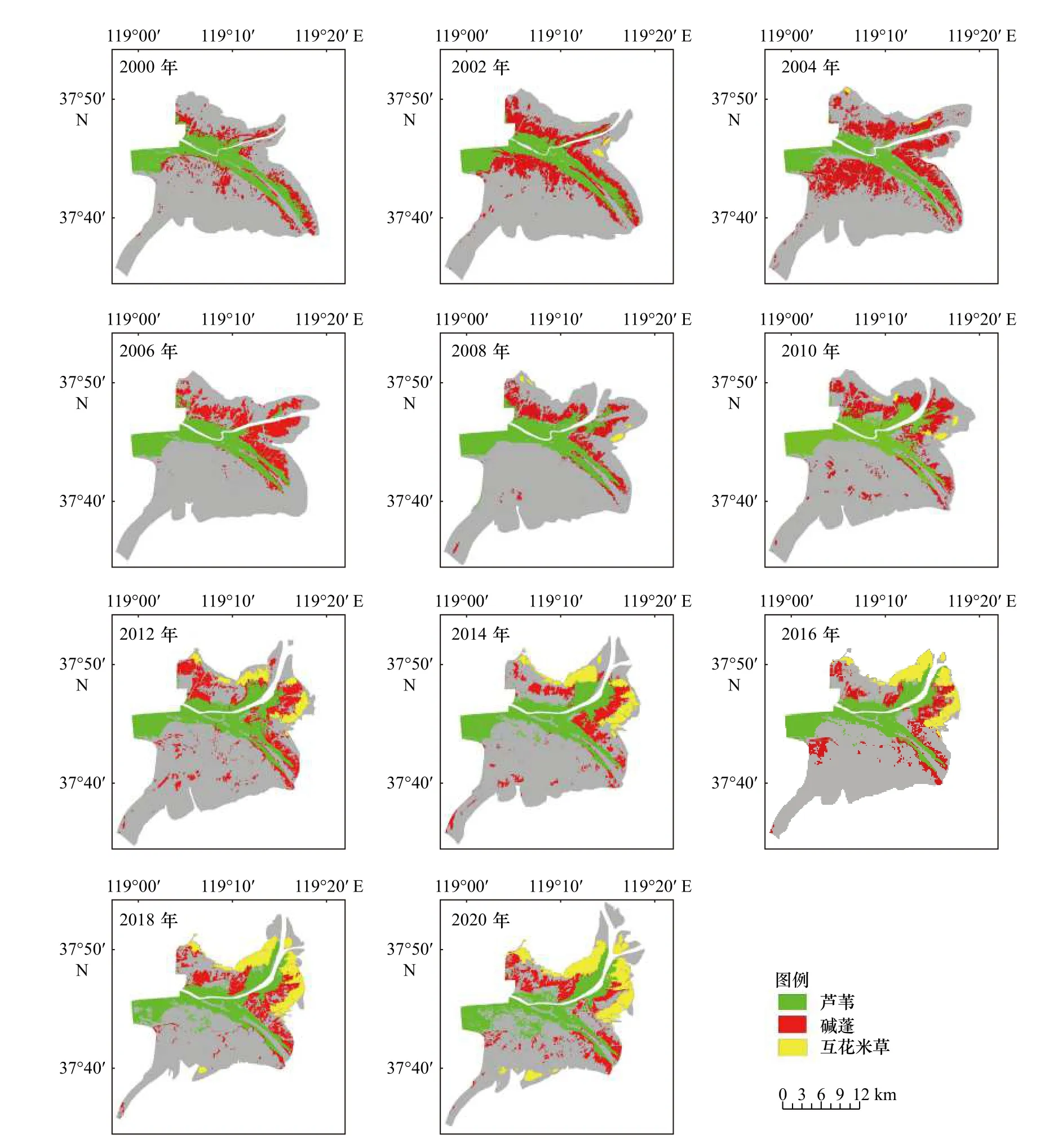
图2 2000-2020年现行黄河口保护区典型湿地植被分布Fig. 2 Distribution of typical wetland vegetation in the Current Huanghe River Estuary Reserve from 2000 to 2020
通过ArcGIS和Fragstats计算3种植被的6种景观指数(图3)。可以发现2000-2020年期间芦苇群落斑块面积总体呈平稳增长状态,斑块所占景观面积比例维持在6%以上,并且最大斑块指数在3种植被中最高。这是因为湿地恢复工程建设过程中,淡水对土壤的洗盐作用,使得芦苇成为此地区的优势物种[37]。芦苇的平均斑块面积在2008年之后呈下降趋势,景观开始呈现破碎化,主要是由于石油开采和旅游修建的道路将原本大的芦苇群落斑块切割为若干个小的斑块。
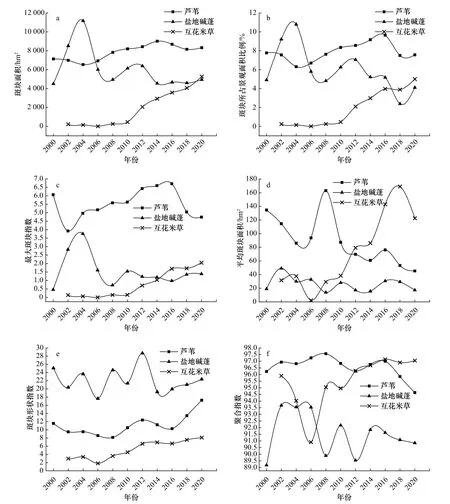
图3 2000-2020年3种典型湿地植被的景观格局指数变化趋势Fig. 3 Change trend of landscape pattern index of three typical wetland vegetations from 2000 to 2020
2000-2020年期间现行黄河口保护区内盐地碱蓬群落的景观格局发生了极大的变化。盐地碱蓬群落斑块面积从2000年的4 503.33 hm2增长到2004年的11 163.51 hm2,碱蓬增长最为明显的区域为大汶流区域。2000年时此区域盐地碱蓬仅呈小片分布,2004年时已大面积分布,这与2002年开始的淡水恢复工程有关。引黄河淡水进入生态恢复区,降低了土壤含盐量,土壤肥力得到改善,因此碱蓬的生物量达到最高值。2006年时大汶流区域的盐地碱蓬大面积退化,碱蓬斑块面积下降到6 007.23 hm2。这是因为保护区内已完成了湿地恢复,人工恢复区内淡水比例过大,这种高水深及低盐分环境不适宜碱蓬生长。此后,碱蓬仍在进一步退化,到2020年碱蓬斑块面积仅为4 945.14 hm2。碱蓬生长需要一定的盐度空间,黄河上游淡水水量的减少,使得其生境中的土壤盐度超标,这是导致此时期碱蓬继续退化的主要原因。碱蓬的最大斑块指数和聚合指数自2004年之后开始呈下降趋势,并且平均斑块面积在3种植被中最小,说明该阶段碱蓬的优势度减小且破碎化程度加剧。此外,碱蓬的斑块形状指数远高于其他两种植被,碱蓬的景观结构最为复杂。
互花米草的斑块面积由2002年的221.85 hm2增加到2020年的5 267.79 hm2,最大斑块指数也在不断增加,表明互花米草在现行黄河口保护区内的优势度不断提高,由最初自然情况下传播的零散分布逐渐形成大片面状分布。2006-2018年期间,互花米草平均斑块面积由2.34 hm2增加到169 hm2,说明该时段内互花米草由小斑块生长合并为大斑块;2002-2020年期间其斑块形状指数呈现小幅度增加态势,互花米草的形状越来越复杂,呈现不规则化。聚合指数有波动增加的趋势,表明互花米草景观聚集度有所增强。
4.3 互花米草时空动态定量评价
作为入侵物种,互花米草的扩张变迁对研究区其他原生植被的景观格局产生着深远影响。从以上结果可知,近20年来互花米草的时空动态变化在3种典型湿地植被中最为显著,且在不同区域的变化存在较大差异(图2)。为此,本文根据互花米草的时空分布特征,将其分为6个分区(A-F),分别探讨各分区互花米草近20年的动态变化(图4)。
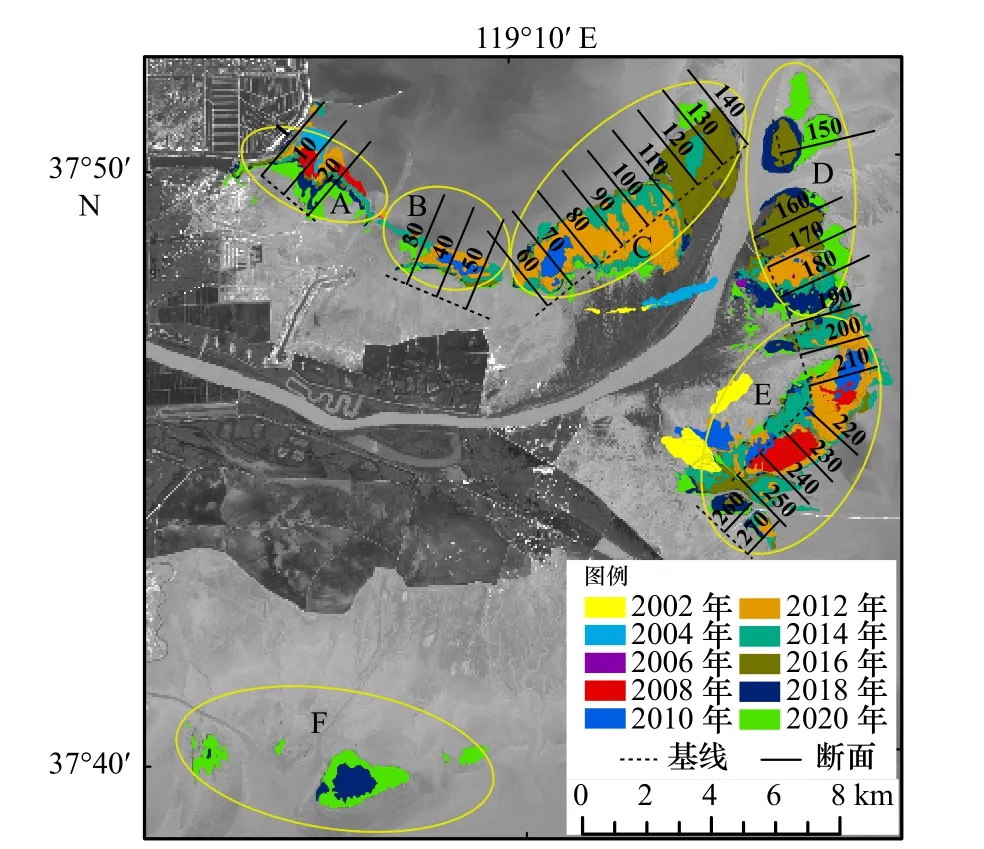
图4 2002-2020年不同入侵年份互花米草分布格局及DSAS 计算2014-2020年互花米草扩张速率所用基线和断面分布Fig. 4 Distribution pattern of Spartina alterniflora in different invasive years from 2002 to 2020 and the distributions of baseline and transects used by DSAS to calculate the expansion rate of S. alterniflora from 2014 to 2020
统计各个分区互花米草的面积(图5),可以发现,2000-2010年区域A互花米草面积呈波动变化状态,这一期间互花米草的剧烈变化与该区域岸线不断向内陆蚀退和频发的极端天气条件(如飓风、风暴潮等)有关。2000-2008年区域A海岸线向后蚀退了1.36 km左右,2008年之后海岸侵蚀速率减缓,岸线趋于稳定,互花米草面积逐渐增加;区域B于2010年出现互花米草分布且面积较小,2014年面积达到最大值,为190.98 hm2;近年来由于黄河输沙稳定,黄河入海口门处陆地面积不断增加,给予了互花米草足够的生存空间,这使得区域C和区域D的互花米草面积不断增加,其中区域C互花米草面积从2010的73.44 hm2增至2016年的1 526.49 hm2,增幅近20倍;区域E于2002年首次出现互花米草,面积为204.21 hm2,2004年时面积下降到15.32 hm2,这是由于受2003年黄河口特大风暴潮的影响,互花米草经过一段时间的恢复期,于2008年开始大片生长;区域F于2018年首次出现互花米草,2020年时互花米草已增加至426.87 hm2,并且有越过潮沟向两侧扩张的趋势。
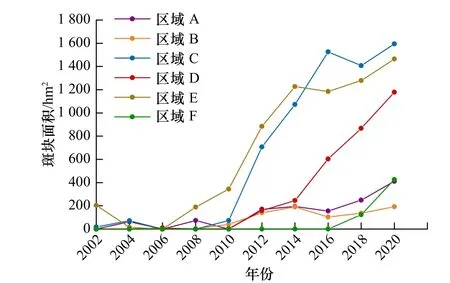
图5 不同区域互花米草斑块面积变化Fig. 5 Variation of Spartina alterniflora plaque area in different regions
综上所述,互花米草在2008年之前处于不稳定发展状态,2008年之后出现暴发式增长。而由表7可以看出,2008-2014年互花米草的年扩展率整体大于2014-2020年期间的年扩展率。因此,为进一步揭示不同区域互花米草的时空动态变化特征及其对碱蓬群落的影响,本文基于数字岸线分析系统(DSAS)分别计算2008-2014年以及2014-2020年期间互花米草向海侵占光滩和向陆侵占碱蓬生境的扩张速率(图6)。经过对比分析,可以将现行黄河口保护区互花米草的扩张分为3个阶段:2008年以前为扩张初期,互花米草种子从5号桩随潮流漂散到黄河口潮间带区域,在合适的潮滩生长,这一阶段互花米草植株稀疏,且生存条件恶劣;2008-2014年为快速扩张阶段,互花米草已经呈大面积斑块分布,面积增速最高达到799.88 hm2/a,年平均扩展率达到54%,且这一阶段大部分区域均表现为向海扩张侵占光滩。其中区域C位于口门处,新生湿地不断增多,因此此处互花米草盐沼的向海扩张速率在所有区域中最高且一些断面处向海扩张速率可达600 m/a左右。此阶段由于互花米草的促淤能力,使得滩面不断增高,潮水很少能够到达碱蓬生长的地带,碱蓬的生存条件受到限制,互花米草向碱蓬边缘扩张,且迅速占领其生存空间,因此该阶段所有区域也表现为向陆扩张迅速侵占碱蓬生境;2014-2020年为缓慢增长阶段,年平均扩展率为9%。除了区域D,其他区域互花米草向海扩张趋势均显著减弱,这是由于滩面高程的限制。只有互花米草的前缘滩面继续淤长到满足其生长需要的潮侵频率以后,互花米草才会继续向海扩张。而大部分区域互花米草均表现为向陆扩张侵占碱蓬生境,但由于互花米草生态位的限制,其向陆扩张速率有所减小。总的来说,近几年互花米草进入了生长的稳定期,并且随着互花米草向陆扩张速率的减小,碱蓬的生存空间将得到一定的改善。
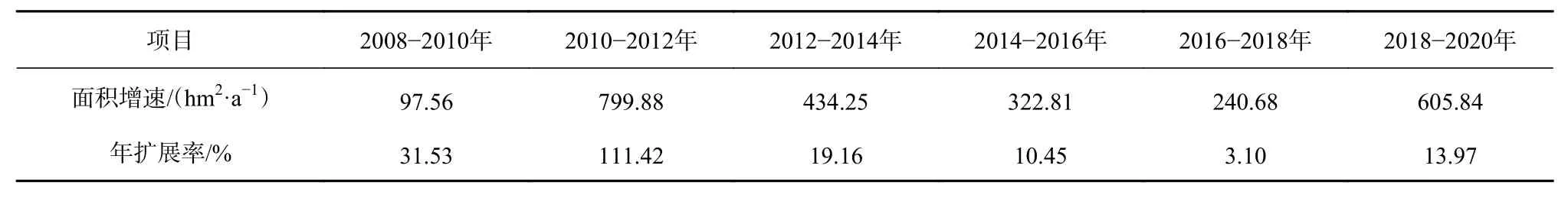
表7 研究区互花米草的新增面积增速及年扩展率Table 7 Area increasing rate and annual expansion rate of Spartina alterniflora in the study area
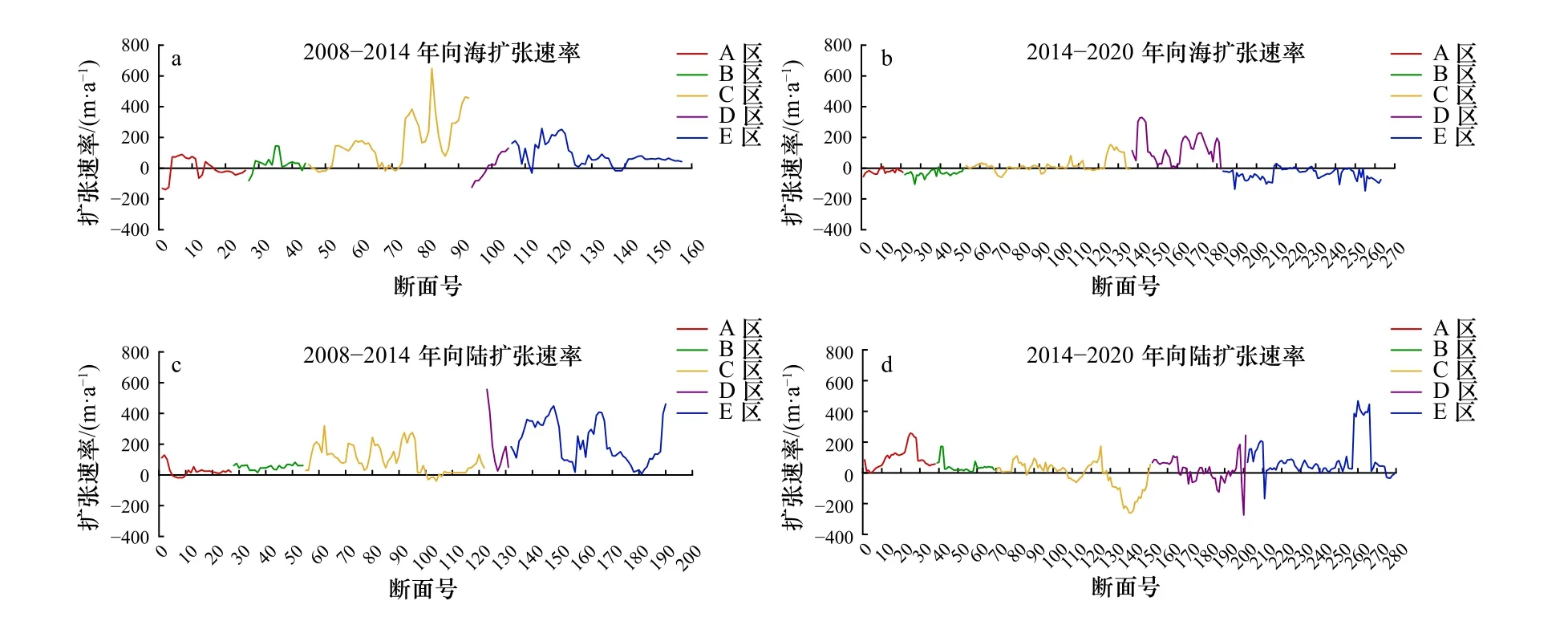
图6 区域A-E互花米草向海和向陆的扩张速率Fig. 6 Seaward and landward expansion rates of Spartina alterniflora in A-E areas
5 结论
本文基于多时相卫星影像数据和野外实地调查数据,结合面向对象和随机森林算法对现行黄河口保护区内3种典型湿地植被进行了精准提取,得到了2000-2020年保护区内芦苇、盐地碱蓬和互花米草的空间分布结果。利用GIS空间分析技术,揭示了近20年来3种植被的景观格局演变特征,并重点讨论了互花米草的动态变化过程。主要结论包括:
(1)基于面向对象和随机森林算法进行长时序湿地植被制图可以得到较高的分类精度,满足湿地植被时空动态分析的需求。
(2)现行黄河口保护区的湿地植被在空间上呈条带状交替分布。芦苇群落是保护区内的优势物种,面积总体呈平稳增长状态;碱蓬的最大斑块指数和聚合指数呈下降趋势,破碎化程度加剧,且斑块形状指数远高于其他两种植被,景观结构最为复杂;互花米草的斑块面积由2002年的221.85 hm2增加到2020年的5 267.79 hm2,其在保护区内的优势度不断提高。
(3)现行黄河口保护区互花米草的扩张过程分为3个阶段:2008年以前为扩张初期,互花米草生长不稳定;2008-2014年为快速扩张阶段,新增米草面积最高可达799.88 hm2/a,年平均扩展率为54%,且海向和陆向扩张都较为显著;2014-2020年为缓慢增长阶段,年平均扩展率为9%,互花米草开始进入生长的稳定期。
(4)本研究使用30 m分辨率的Landsat系列影像来提取植被信息,然而该影像不适合小尺度植被斑块的精细调查。在今后的研究中,可以基于图像超分辨率重建算法将时间序列数据与更高空间分辨率的数据(如Worldview-2/3、SPOT、GF-2等)融合来进行细小植被斑块的识别和监测;此外,由于缺少植被生物量调查数据和潮滩高程数据,本研究对互花米草入侵引起的生态环境效应的分析还不够深入,而互花米草盐沼的扩张与潮滩地貌之间的关系也需要进一步探讨。
