基于野外观测检验动物廊道选址模型的准确性
——以白头叶猴为例
2022-01-16陈秋彤刘骏杰覃子浏
陈秋彤,刘骏杰,覃子浏,明 霜,姬 翔,杜 钦,*
1 桂林理工大学旅游与风景园林学院,桂林 541006 2 北京市企业家环保基金会,北京 100012
生物多样性的降低正在全球范围内加速,而生境破碎化是物种生存的重要威胁之一[1-2]。生境破碎化使野生动物迁移和建群受到限制,改变了种群迁移扩散的模式,限制不同种群间的遗传基因交流,增加近亲繁殖可能性,使后代存活能力下降,增加了种群灭绝的风险[3]。大量研究表明,构建廊道可以维持和提高生境景观连通性,是缓解生境破碎化负面影响的重要策略之一[1,4-6]。
廊道的构建第一步需要回答的问题是:在何处构建廊道?对此,已有许多廊道选址模型专门用于回答此问题。其中,应用较为广泛的是最小成本路径(Least Cost Path,LCP)和条件最小成本廊道(Conditional Minimum Transit Cost,CMTC)[2]。
LCP模型假定动物从源栖息地(模型中设为“源”)出发,到达需连通的目标栖息地(模型中设为“目标”),计算穿越不同景观单元所累积耗费的阻力代价,同时衡量物种迁移距离以及穿越不同景观难易程度[7]。该模型基于最短路径算法选择出累积耗费阻力最低的路径,作为动物廊道构建的最佳位置[8]。
由于LCP存在仅生成单一像元宽度路径的缺陷[6],CMTC模型对其进行了改进,优化了LCP单一路径的固有缺陷,考虑了指定源汇间物种迁移的最佳路径和次优路径的重要性,提高了动物迁移廊道模拟的准确性[8]。
过去的几十年中,这两种方法常常被用来确定廊道构建的具体位置,具体如薛亚东等运用LCP模型结合景观遗传学方法,绘制了云南境内滇金丝猴(Rhinopithecusbieti)种群间的潜在扩散廊道[9];Alexander等和曲艺等分别利用LCP模型构建了蒙大拿州雨狐(Vulpesvelox)和中国东北虎(Pantheratigrisaltaica)核心栖息地廊道[10-11],Loro等和Pinto等分别利用CMTC模型生成西班牙狍(Capreoluscapreolus)和巴西大西洋森林栖息地动物廊道等[12-13]。虽然过去这些研究从理论上为动物廊道的构建提供了具体的空间位置参考,但在实践过程中,管理者和动物学家更为关注的问题是:这些廊道理论模型获得的路径位置,是否就是动物迁移的实际发生位置,这些模型能准确预测动物迁移的实际发生位置吗?
事实上,国际上已有研究者对这一问题进行了关注。具体如LaPoint等在美国近奥尔巴尼(Albany)350 km2的渔貂(Martespennant)栖息地内,通过为渔貂佩戴GPS监测环圈记录到的迁移路径,对LCP模型进行检验[14];Bond等在30000 km2的坦桑尼亚塔兰吉尔生态系统(Tarangire Ecosystem),利用距离采样检测和GPS项圈定位所收集的非洲角马(Connochaetestaurinus)数据检验了LCP模型的性能[15]。尽管如此,目前的不足仍在于(1)之前的研究案例仅在大尺度上对模型进行了检验,缺乏在小尺度范围的验证案例;(2)廊道选址模型在国内虽已得到较多应用,却鲜有研究案例能回答上述问题。
白头叶猴作为我国特有、广西独有的珍稀濒危物种和桂西南喀斯特关键带亚热带季雨林生态系统的旗舰种及伞护种,是国家一级保护野生动物以及全球25种濒危灵长类动物之一,被IUCN濒危物种红色名录列为极危(CR)物种,全球种群数量仅1200多只[16]。20世纪80年代初,由于农民持续在白头叶猴核心保护区开垦农田、种植经济作物,砍伐石山森林植被等,种群分布范围愈加狭窄,栖息地斑块间联系“通道”被掐断,“人进猴退”的矛盾加剧,猴群数量下降到仅剩300余只[17-18],白头叶猴及其栖息地保护迫在眉睫。前人虽然有过许多研究,但主要集中在其生活习性和行为特征等方面[19-20],少有研究关注白头叶猴的生态廊道[21]。本研究以崇左白头叶猴为例,通过实地观测记录白头叶猴的迁移轨迹,检验LCP和CMTC两种方法模拟结果的准确性。主要回答以下问题:(1)与实地观测所得的真实迁移廊道对比,LCP模型和CMTC模型是否能准确模拟出白头叶猴的迁移廊道?(2)作为LCP模型的优化版,CMTC模型其结果是否优于LCP模型?(3)若两种模型不能准确模拟,其原因可能是什么?
1 研究区域和研究方法
1.1 白头叶猴种群特征和研究区域
白头叶猴仅分布于广西左江和明江之间不足200 km2的一片狭长地带内,栖息于喀斯特岩溶石山地区,现存种群栖息地空间分布呈现小而分散的特征[18]。白头叶猴善攀爬跳跃,群居于喀斯特岩溶石山崖壁洞穴或石壁平台,以植物嫩叶、枝条和野花野果为食,采食的植物种类主要有构树(Broussonetiapapyrifera)、斜叶榕(Ficusgibbesa)、朴树(Celtissinensis)、木棉(Gossampinusmalabarica)等[22],因觅食需求,白头叶猴会增加日漫游距离和时间迁移至其他石山植被处搜寻食物资源,白头叶猴的日漫游距离一般为400—600 m[19,23]。白头叶猴种群结构主要有一雄多雌的两性群和全雄群两种类型,每个猴群一般有4—5处夜宿点,猴群定期在这些夜宿点间更换,以适应不同的季节气候、躲避危险以及合理利用领域内的食物资源[24]。综上,栖息地斑块间的“通道”对白头叶猴日常迁移扩散以满足生存需求极其重要,维持栖息地连通性是保护白头叶猴的重要举措。
研究区域(22°25′30″—22°28′15″N,107°51′40″—107°56′30″E)即渠楠自然保护小区隶属于广西崇左市扶绥县山圩镇昆仑村渠楠屯(图1),面积约31.68 km2,紧邻崇左白头叶猴国家级保护区岜盆片区九重山东南部。渠楠自然保护小区所处的桂西南喀斯特山区,位于中缅国际生物多样性热点区内,是中越交界的喀斯特廊道优先区,同时是《中国生物多样性保护战略和行动计划(2011—2030)》确定的中国生物多样性保护优先区域之一[16]。尽管研究区域只位于崇左白头叶猴国家级自然保护区边缘,但其白头叶猴种群数量占全球总量的比例高达20%,是极其重要的自然栖息地[25]。研究区以峰丛喀斯特地形地貌为主,岩溶石山林立。虽然在保护区和渠楠自然保护小区相继建立后,非法盗猎制作“乌猿酒”的不良风俗得以杜绝,但毁林开荒等人为活动影响,峰丛洼地和谷地被不断占用和侵蚀,白头叶猴在石山间联系的通道被阻断。根据当地民间组织及社区巡护队联合监测,共记录到31群249只白头叶猴,分布于渠楠屯及周边6片岩溶石山斑块中的八十余处夜宿地内。

图1 研究区域位置及用地类型图Fig.1 Location of the study area and the type of land useP1—P11表示栖息地源斑块
1.2 观测的白头叶猴迁移廊道
以野外实地观测所得的目标种迁移扩散数据作为检验模型性能的依据是最直接且直观的方法,观测数据应满足以下要求:(1)通过扩散轨迹能识别出动物迁移过程的源和目标区域;(2)能识别出除源斑块外影响动物迁移的重要节点斑块;(3)数据收集时间应足够长,以反映同一范围白头叶猴迁移频率[8]。
本研究运用实地观测法收集白头叶猴实际迁移廊道。观测期间自2017年6月—2019年12月,观测信息有两个来源:(1)由观测小组实地观测记录,在观测期间内每月连续观测7—10 d,具体观测时段为清晨(6:00—9:00)和黄昏(17:00—19:30)[26],该时段是白头叶猴觅食及短距离迁移活动的高频阶段;(2)由渠楠自然保护小区巡护队员及村民日常目击记录,目击时段为村民田间耕作期间(6:00—11:00,14:00—17:00),该时段为白头叶猴觅食后和午休后的长距离迁移高峰期[26]。对观测结果实行“重复验证”的方式,重复观测村民提供的目击位置,同时委托村民加强关注小组实地观测记录的白头叶猴迁移轨迹位置。野外观测记录的主要内容有:迁移源斑块与目标斑块、每次目击的迁移轨迹(通过识别白头叶猴在土壤或植被所留活动痕迹判断)、每次目击迁移轨迹长度。基于野外观测的记录,进一步汇总统计分析的内容有:同一目击点单位时间内的目击频率(按月进行统计)、同一目击点多次记录迁移轨迹后形成的迁移轨迹范围、各迁移轨迹范围的宽度,汇总统计后获得的同一目击点的迁移轨迹范围即为本研究中白头叶猴迁移的实际廊道。
1.3 最小成本路径
最小成本路径分析方法源于成本距离模型,以栅格化的景观阻力表面为依据,生成从运动起始点(源)到目标之间的累积最小成本距离的路径[27]。利用ArcGIS 10.5成本距离工具计算累积的成本距离需要输入“源”图层、“目标”图层以及阻力面图层,输出的每个栅格单元值即表示从此栅格到最近“源”的最小累积成本距离,该运算是基于图论原理和8个领域计算原则,利用节点-链接表示相邻单元间的成本距离[28],源单元到直接相邻单元成本距离计算公式为:
Ni+1=Ni+(ri+ri+1)/2
源单元到对角相邻单元成本距离计算公式为:
式中,i为“源”单元,i+1为相邻的目标单元,r为单元阻力系数,Ni为源单元的阻力成本,Ni+1为源单元到目标单元的累积成本距离[29]。本研究以观测廊道连接的11个岩溶石山斑块,作为白头叶猴迁移的栖息地“源”斑块进行模拟。
1.3.1航拍影像解译
研究区域的地理信息来源于2019年7月29日无人机拍摄的正射影像,空间分辨率4 m,影像精度较高,土地利用类型可辨识性好,满足研究区域景观分析的精度要求。运用ArcGIS 10.5,将正射影像图进行地理配准和定义投影,进行人工目视解译,经过实地勘察测绘以及专家检验对解译结果进行反复比对和校正。其中考虑到白头叶猴生态习性及其对各景观用地类型的敏感程度,将研究区域用地划分为岩溶石山、农田、田埂路、桉树林、剑麻种植地、柑桔种植地、村道、乡道、矿坑、村庄、水域,其中由于柑桔生长年限不同,其植株冠幅和密度不同,因此将柑桔地分为生长期1—3年和4—6年两种,共12种用地类型,最终生成研究区域的土地利用类型现状图(图1)。
1.3.2阻力面构建
确定物种在异质景观中迁移扩散所要克服阻力的关键步骤是对不同景观单元赋予相应的阻力系数,以量化表现不同用地类型和环境因子对物种迁移的促进或阻碍程度[30-31]。阻力因子的选择和阻力赋值应基于实地观测、调查,考虑目标种的生态习性以及扩散行为特征[32]。本文结合实地观察和文献资料,并咨询相关专家意见,选择土地利用类型、水源地距离(大量目击记录显示白头叶猴下地迁移以寻找饮水点)、夜宿地距离(猴群更换夜宿地的迁移行为一般成群行动,迁移路线短,以安全性为优先考虑因素)、桉树林斑块距离(大量观测记录显示,白头叶猴常以人工桉树林作为眺望点甚至迁移通道,以保证自身迁移的安全)作为阻力因子。
首先,对12种用地类型进行阻力值设定,取值范围为1—1000,其中11个栖息地斑块的阻力值设为最小值1,阻力值越高,表示物种穿越该用地类型难度越大,一般阻力数值大小顺序不变,对廊道生成结果影响较小[27];其次,通过欧氏距离分析工具,划分水源地距离、夜宿地距离、桉树林斑块距离的阻力系数,距离越近则阻力值越小,其中,桉树林斑块通过提取猴群和夜宿地密度较高区域周边的桉树林进行识别;最后,通过yaahp软件层次分析法(Analytic Hierarchy Process,AHP)确定四个阻力因子的权重[33],根据专家、社区巡护队及村民的意见,采用1—9标度法对四个阻力因子的相对重要性进行两两比较评价,最终确定的阻力因子权重分别为土地利用类型(0.568)、水源地距离(0.120)、夜宿地距离(0.059)、桉树林斑块距离(0.252),利用ArcGIS 10.5加权叠加分析生成像元尺度为3 m的栅格阻力面(表1)。
1.3.3最小成本路径生成
本文通过ArcGIS 10.5平台,以观测所得廊道连接的栖息地斑块互为源和目标,利用要素转点工具提取斑块重心作为“源点”,以栅格阻力面为计算成本,利用距离工具中的CostDistance工具计算各像元到指定源的累计成本距离,利用CostBackLink工具定义任一像元延最小成本路径到指定源的方向,最后利用CostPath工具生成累计成本距离最小的路径,即模拟的最佳迁移路径(图2)。
1.4 条件最小成本廊道
CMTC廊道基于最小成本路径模型,通过叠加“源”和“目标”的成本距离栅格表面,提取大于最小成本距离一定阈值范围内的成本距离栅格作为潜在廊道。当提取范围为最小成本距离(Min CMTC+0% CMTC)时该廊道结果与同源的LCP一致,阈值越大,廊道的宽度和冗余性增大[34]。计算公式如下:
CMTC(S,V,D)=Cumulative cost(V,S)+Cumulative cost(V,D)
CMTC为条件最小成本距离,Cumulative cost为累积成本距离,S为“源”斑块,D为“目标”斑块,V为S到D经过最小成本路径中的任意一点。
本文以11个源斑块面要素互为源和目标,基于栅格阻力面,利用ArcGIS 10.5空间分析模块分别计算出11个源斑块的累积成本距离栅格表面,并将表面两两求和形成每对源斑块间的成本距离表面,提取该成本距离表面中指定阈值范围的栅格作为最终的模拟廊道结果(图2)。CMTC廊道的阈值必须考虑实际的生态过程来确定,本文选取Pinto等建议的最具使用潜力的10%范围(即 Min CMTC+10% CMTC,Min CMTC为最小成本距离)内的潜在廊道作为最终的模拟廊道[13]。
1.5 两种模拟廊道的准确性判定
通过比较模拟廊道位置与实际观测廊道的重合程度,以判断模拟结果准确性。若模拟廊道完全落入对应的实际观测廊道(即迁移轨迹范围),则表示该模拟结果能准确识别;若模拟廊道完全未落入对应观测廊道,则该模拟结果完全不准确;若模拟廊道部分落入,则为部分准确。

表1 阻力值及阻力因子权重设定
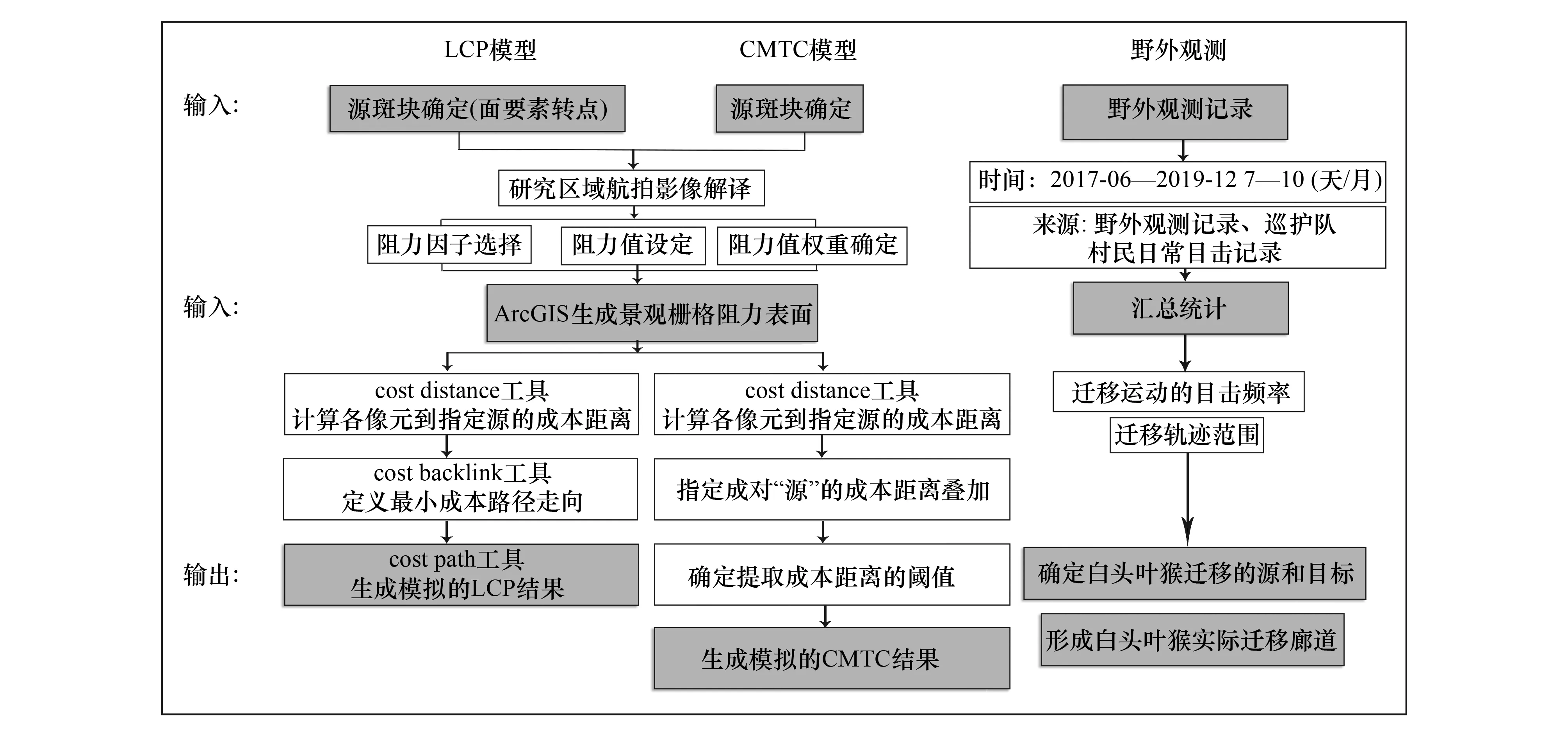
图2 两种模型及野外观测的操作步骤Fig.2 Two models and field observation stepsLCP:最小成本路径 Least cost path;CMTC:条件最小成本廊道 Conditional minimum transit cost
2 结果
2.1 观测廊道
观测记录所得的迁移廊道显示(表2),在11个栖息地源斑块间共有17条较高频率的迁移廊道(R1—R17),面积共约0.31 km2,廊道平均宽度为124 m,其中R4和R7以P1和P5为源,R13、R14以P3和P4为源,其余源斑块间仅观测到单一的迁移廊道(图3)。由于P1面积较大,白头叶猴数量多且夜宿地分布密度高,人类活动干扰较少,以P1为源的斑块间白头叶猴迁移频率较高。观测所得的白头叶猴迁移廊道多分布于源斑块间距离最短的区域,成对斑块间可能存在多条通道。
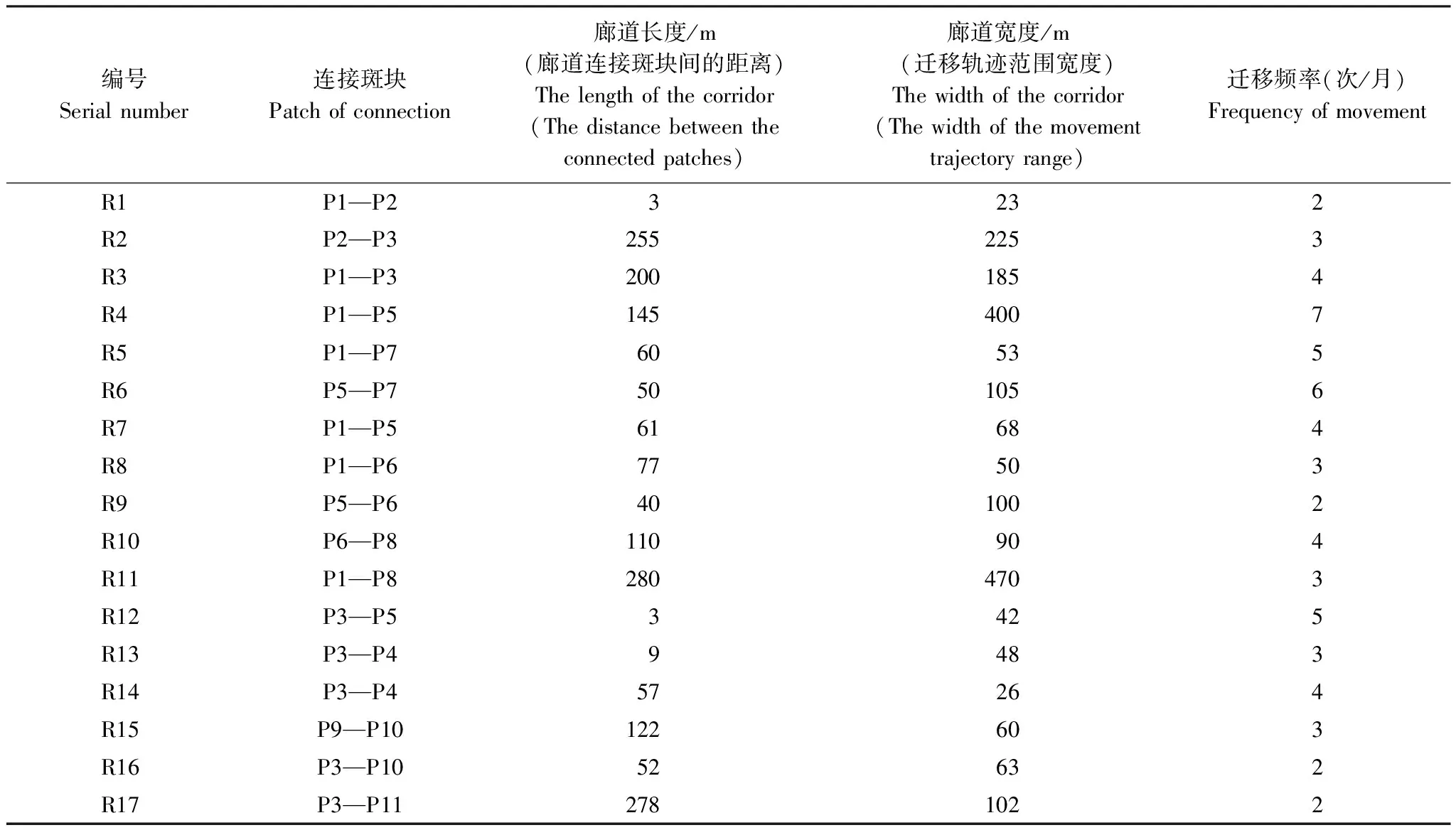
表2 观测的白头叶猴迁移廊道

图3 观测廊道及两种模拟廊道的位置Fig.3 Location of observation corridor and two simulation corridorsP1—P11表示栖息地源斑块;R1—R17为观测所得的白头叶猴迁移廊道;L1—L15为模拟的LCP结果;C1—C15为模拟的CMTC廊道
2.2 LCP模型模拟结果
如图3所示,以观测廊道连接的11个岩溶石山斑块质心为源,共生成了15种源汇组合形式的LCP模拟结果(L1—L15),虽然P1和P5间、P3和P4间都分别存在两条观测廊道,但结果显示,成对源间仅能生成单一路径。
2.3 CMTC模型模拟结果
如图3所示,根据15组源汇组合形式的叠加成本距离表面,提取10%最小成本范围栅格,共生成了15组模拟的CMTC廊道(C1—C15)。
2.4 两种模拟廊道与观测廊道的比较
首先,LCP模拟出的7条廊道(L1、L6、L7、L11、L12、L13、L14)完全落入对应的观测廊道中,3条(L4、L8、L15)部分落入对应的观测廊道中,5条(L2、L3、L5、L9、L10)与观测廊道完全无交集(图3)。LCP模拟结果的完全准确率为46.7%,部分准确率为20%,完全不准确率为33%。
其次,CMTC廊道模拟中有4条廊道(C1、C6、C8、C12)完全准确落入对应的观测廊道中,其余11条(C2、C3、C4、C5、C7、C9、C10、C11、C13、C14、C15)均部分落入对应的观测廊道,没有出现完全不准确的情况。CMTC廊道的完全准确率为26.7%,部分准确率为73.3%,完全不准确率为0。由于CMTC廊道考虑了最小成本的次优廊道,其完全准确率较LCP模拟结果低20%,但消除了完全不准确的廊道,因而从廊道位置模拟的准确性上看,CMTC模型其结果优于LCP模型。
总体来说,与观测记录的真实廊道相对比,LCP模型和CMTC模型都能较准确地模拟出白头叶猴的迁移廊道位置,且LCP模型的完全准确率较高,CMTC模型的出错率较低(表3)。
3 讨论
3.1 模拟廊道与观测廊道位置偏差的原因
3.1.1输入“源”的要素类型
从两种模拟廊道的位置上看,CMTC与LCP的差异较大,如图3C、3E、3J所示,L3、L5、L10与观测廊道完全不一致,而相对应的CMTC廊道(C3、C5、C10)则与观测廊道有部分交集,且准确性都较高。该结果的原因可能是两种方法在模拟过程中,对“源”要素类型处理上的不同。LCP模型通过提取源斑块的重心作为输入的源,生成斑块间的最小成本路径,而CMTC廊道模拟以斑块面要素为源。提取斑块重心为源的做法忽略了出发点到斑块边缘的距离,若斑块面积较大,基于最短路算法生成的最小成本路径结果可能会低估目标种迁移的成本距离,同时也可能高估了白头叶猴本身对环境范围的感知能力,从而导致模拟廊道与实际观测廊道间的偏差。这一结果表明,输入“源”要素类型的差异对模拟结果产生较大影响。

表3 两种模拟廊道与观测廊道的对比结果
3.1.2阻力面栅格尺度
由图3—D可知,观测的R4和R7都以P1—P5为源,两种方法模拟结果(L4、C4)分别准确落入了观测廊道R4和R7中,但C4无法准确模拟观测的R4廊道位置,而R4廊道是白头叶猴平均迁移频率最高的区域(平均7次/月),说明该区域具有吸引白头叶猴迁移活动的因素,但在CMTC廊道模拟过程中无法被准确识别。根据实地调查发现,P1与P5斑块间距离约150—300 m,用地类型多为农田,田间散布1—3 m宽的微小石块,高30—50 cm,有目击记录显示白头叶猴常利用具有一定高度的石块作为瞭望点,以确保迁移过程的安全;其次,该区域灌溉水塘较多,是白头叶猴地表取水点。由于石块和部分灌溉水塘面积小于阻力面构建的栅格尺度,从而无法完全识别这些关键的景观元素,降低了阻力面构建的准确性。该结果表明了阻力面栅格尺度设定是影响两种模型模拟白头叶猴迁移廊道准确性的重要原因之一。该结论与陈春娣等对不同景观粒度影响最小成本路径模拟的析因测试研究结论相似,虽然该研究中景观结构和阻力赋值方式不同(利用无量纲相对值构建模拟景观的阻力面),但该研究结果显示,景观粒度差异对路径模拟有显著影响[31]。解译影像精度越高、栅格尺度越能准确反映实际的景观特征,模拟廊道的准确性越高[35]。阻力面栅格尺度应考虑目标物种对栖息地内微小景观元素的敏感程度来设定[36-37],一般栅格尺度不应大于区域内影响目标种迁移路径选择的最小景观元素或线性景观要素的最小宽度[27]。
3.1.3用地类型的转变
从图3I中L9和C9与对应的观测廊道(R10)的重合情况可以看出,L9与观测廊道完全无交集,C9仅有部分准确落入了观测廊道R10中,且重合程度较低,其原因是该区域在白头叶猴迁移数据观测记录期间(2—3年)用地类型发生了显著变化,即R10观测廊道区域内约2530 m2的农田转变为柑桔种植地,柑桔植株有刺,种植期达到2—3年的密植柑桔林对白头叶猴迁移产生了阻碍作用。这一结果表明栖息地用地类型变化对该区域景观连通性有较大影响,从而改变了白头叶猴对迁移路径的选择,但在廊道模拟过程中缺乏对该因素的考虑,导致模拟结果与观测的实际情况出现不一致。该结论与Burel等的研究结论一致,其通过测试六年内由玉米种植结构变化引起的区域连通性变化情况,表明作物多年轮耕、演替等造成的景观空间结构和用地类型变化会导致区域连通性变化,并强调了连通性分析应该是一个动态变化的过程[38];Mui等在布兰丁海龟(Emydoideablandingii)栖息地季节性连通性变化研究中也有相似的结论,该研究指出季节性变化引起的栖息地斑块重要性变化,也会对动物行为产生一定的影响[39]。
3.1.4动物迁移行为的复杂性
如图3B显示,以P2—P3为源的模拟廊道L2与对应的观测廊道R2完全无交集,该结果的原因可能是模拟的L2路径经过低阻力值的P1斑块,使该路径的累积阻力较低或与距离较短的R2廊道阻力相近,而实际白头叶猴迁移相较于使用耗费阻力更小的长距离路径,有可能更愿意选择短距离的R2廊道区域,以快速穿越,降低下地风险。这说明当源斑块间存在多条累积阻力相近的路线时,难以准确判定动物最终选择的迁移路线,该原因具有复杂性,且难以纳入到模型构建过程中,主要体现在两个方面:其一,模型理论上,LCP模型的算法本身已经假设了动物在迁移活动前对栖息地阻力信息、起始点和终点完全知晓并能始终选择最佳路径,这在实际的迁移过程中是难以实现的,该假设导致无法具体考虑动物个体的迁移意愿及其对环境范围的感知能力等,具有一定的局限性[40-41];其二,物种特性上,白头叶猴迁移行为与其本身复杂的生态习性密切相关,首先,不同组成结构的猴群迁移行为特征并不一致,两性猴群和全雄猴群迁移活动频率较低,迁移距离短,在一定家域范围内具有固定性,而两性猴群中的雄性亚成年个体性成熟后会离开原生猴群并争夺其他两性猴群的猴王位置,以获得交配机会和生活领地[42],这一期间迁出的雄性独猴具有较强的游荡性,迁移距离更远,但迁移方向难以预估,其次,白头叶猴作为叶食性灵长类动物,受季节性差异影响下的食物资源获取难度(夏易冬难)对其日漫游距离和家域范围调整具有明显影响[20],从而一定程度上影响白头叶猴迁移距离和频率,但此类因素的影响程度难以预估,无法量化并纳入到廊道模型阻力面构建中,导致廊道模拟结果与实际出现偏差。
3.2 廊道选址模型是否能准确预测动物迁移的实际发生位置
本研究运用野外实地观测的白头叶猴迁移数据,对照检验了LCP模型和CMTC模型两种廊道选址模型在小尺度白头叶猴栖息地进行廊道模拟的准确性,研究发现两种模型的准确率分别为66.7%和100%,表明两种模型都能较准确地预测白头叶猴迁移的实际发生位置,且CMTC模型准确性优于LCP模型。
本研究基于小尺度栖息地廊道进行模拟检验,而国外对大尺度栖息地动物迁移廊道的模拟验证也有类似的发现,但不同物种下模型准确性检验结果不尽相同。Epps等通过无线电遥测和轶事报道信息监测到美国加利福尼亚东南部约76780 km2范围内40对山脉间大角羊(Oviscanadensisnelsoni)的运动轨迹,以此检验了景观遗传学优化的LCP模型预测的大角羊扩散路径,其预测的准确率高达77.5%[43],较本研究中LCP模型模拟白头叶猴迁移路径的准确性高,表明了Epps等构建的拟合模型与目标物种大角羊迁徙运动规律的匹配度较高。Poor等通过GPS定位收集了北美蒙大拿州25200 km2栖息地内叉角羚(Antilocapraamericana)位置信息,比较了8组由不同阻力赋值方式、不同廊道模型和不同季节特征组合模拟的廊道,该研究通过计算模拟廊道覆盖叉角羚位置的百分比来评估模拟结果的准确性,与本研究的重合程度比较方法类似,在阻力面的10%阈值范围内,模拟的秋季最小成本走廊覆盖叉角羚GPS位置数量为63%[44],该结果与本研究中LCP模拟准确性结果相近。由此可见,同种廊道模型在不同物种或不同尺度下同样具有适用性,但其结果的准确性存在差异[45-46]。实际上,廊道模型的准确性取决于模型构建与目标物种对栖息地利用规律之间的关联程度[47],如何构建与物种迁移行为规律相匹配的廊道模型以及模型的不确定性会对动物迁移产生怎样的影响,仍需更多案例研究。
致谢:感谢广西崇左白头叶猴国家级自然保护区岜盆保护站站长、渠楠自然保护区、广西生物多样性研究和保护协会以及宋晴川等对本研究提供的帮助。
