硫、硅肥配施对铜污染下水稻幼苗生长及其生理特性的影响
2022-01-14袁源远卢志红吴建富魏雪娇
袁源远,颜 晓,2,陈 晨,卢志红,2*,吴建富,2,魏雪娇
(1.江西农业大学 国土资源与环境学院,江西 南昌 330045;2.江西省鄱阳湖流域农业资源与生态重点实验室,江西 南昌 330045;3.厦门海关技术中心,福建 厦门 361013)
【研究意义】铜作为植物所必需的微量营养元素之一,同时,也是污染环境的主要重金属元素之一[1]。最新发布的全国土壤污染状况调查公报[2]显示,铜的点位超标率为2.7%,在所有污染因子中排第四,作为耕地主要污染因子之一,严重影响了农田作物产量和品质。【前人研究进展】铜参与植物多种重要的生理代谢活动,如光合作用、呼吸作用以及氧化应激保护等[3-4]。研究表明土壤中适量铜有助于作物的生长发育,而过量铜元素导致的铜胁迫则会对植物体产生毒害作用。相关研究显示,铜胁迫对农作物的毒害作用主要是通过诱导植物细胞产生并积累大量活性氧,引起氧化胁迫,使得膜脂过氧化,从而破坏植物细胞结构,甚至导致细胞死亡[5]。同时,土壤中铜浓度过高还会破坏水稻光合系统稳定、降低氧化还原体系中的相关酶活性。前人研究发现,植物体内存在两类防御系统可有效清除活性氧:一类是非酶促防御系统,以抗坏血酸(AsA),谷胱甘肽(GSH)等含硫小分子物质的抗氧化物质为主;一类是酶促防御系统,以过氧化物酶(POD)、超氧化物歧化酶(SOD)等抗氧化酶类为主[6]。【本研究切入点】大量研究结果[7-10]表明,硫元素和硅元素对于稳固水稻细胞结构,维持光学系统的稳定以及提高氧化还原相关酶活性具有重要意义,施用硫肥和硅肥在很大程度上可以减缓重金属污染对农作物所造成的危害。硫元素作为植物生长所必需的中量营养元素之一[11],是半胱氨酸(Cyst)、谷胱甘肽(GSH)、植物螯合肽(PCs)及金属硫蛋白(MT)等含硫有机化合物的重要组成部分[12]。这些非蛋白巯基化合物和硫又构成了植物体内重要的防御系统,在调节植物生长发育和植物对逆境条件的反应中起着重要作用,在很大程度上可以缓解因重金属毒害引起的植物体内自由基所产生的氧化胁迫[7,13]。有研究表明[9,14],在缺硫状态下,植株体内Cyst、GSH、PCs 含量和蛋白质水平等显著下降,导致植物体生长迟缓,抵御非生物胁迫能力低下。施加硫肥可增加水稻体内半胱氨酸和谷胱甘肽等的含量,促进铜元素的迁移并降低铜的生物有效性,缓解铜胁迫对水稻植株的毒害[15-16]。硅元素虽然目前仍被认为是高等植物的非必需元素,但对于调节植物体的整个生理代谢过程以及提高对外界胁迫的抵抗能力意义重大。尤其在改善由过量金属诱导的植物毒性方面,硅元素起着极为重要的作用,此外,还能增加植物的生物量积累和作物产量[17-18]。故施用硅肥还是较为普遍的一种减轻重金属胁迫的方法。水稻作为喜硅作物,对硅元素的需求也远高于其他植物体。已有学者研究表明[10,19],硅元素对重金属的缓解除了限制根部对金属离子的吸收和固定外,主要就是通过调节次生代谢产物的合成、代谢和修饰,改变各种抗氧化剂的活性来影响植物的氧化状态,改善膜的稳定性,从而增强胁迫耐受性。施加硅肥后,还可通过提高超氧化物歧化酶(SOD)、过氧化物酶(POD)、抗坏血酸过氧化物酶(APX)和过氧化氢酶(CAT)等抗氧化防御系统相关酶活性,降低活性氧引起的氧化损伤,从而提高亚麻植株对铜毒害的抗性[20]。【拟解决的关键问题】因此,合理调节硫、硅营养元素的供应状况可能影响到植物对过量铜的耐性。本研究基于硫硅元素可有效调节植物体抗逆性,以水稻为材料,探讨外源硫、硅肥配施对稻田铜污染下水稻幼苗生长及其生理特性的影响,为阐明硫、硅肥缓解铜污染对水稻毒害机理提供理论依据,以期为防止水稻幼苗铜危害提供相应的解毒途径。
1 材料与方法
1.1 试验材料
本试验于2013年在江西农业大学科技园网室内进行。供试土壤为东乡铜矿孝岗镇张坊乡被污染农田的耕层土,肥力中上,质地为壤土,有机质22.39 g/kg,碱解氮153.47 mg/kg,速效磷28.63 mg/kg,速效钾97.09 mg/kg,全量铜213.82 mg/kg,有效铜89.77 mg/kg,pH值4.31,有效硫40.89 mg/kg,有效硅196.45 mg/kg。
供试品种为常规稻中早25;硫肥采用硫磺,其中含S 99.99%;硅肥用分析纯硅酸钠(Na2SiO3·9H2O),其中含SiO221%。
1.2 试验设计
共设9个处理(表1),硫磺设3水平,分别含S为0,0.013,0.026 g/kg,分别表示为S0、S1、S2;硅酸钠设3水平,分别含Si为0,0.05,0.1 g/kg,分别表示为Si0、Si1、Si2。此试验为完全随机设计,重复4次,共计36盆。
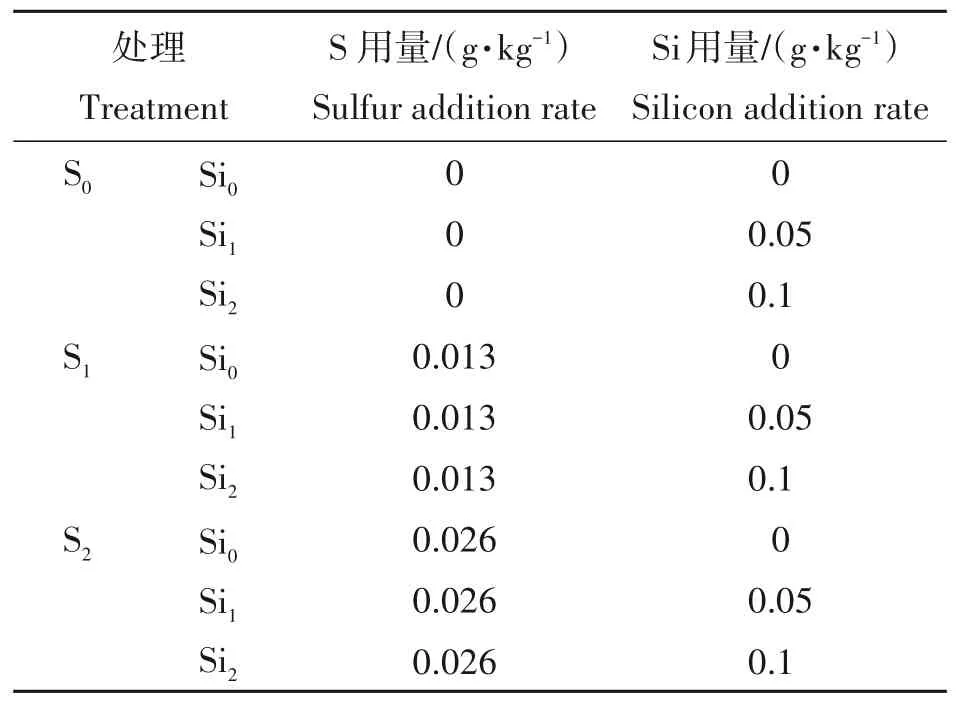
表1 试验设计Tab.1 Design of experiment
试验盆钵为白色塑料盆,盆长30 cm,宽20 cm,高10 cm,每盆装风干土7.5 kg。播种前1个星期加入化学纯氯化铜,使土壤含铜量(以纯Cu 计算)达到400 mg/kg(国家农田土壤环境质量三级标准[21]),施入后与盆中土搅拌均匀,盆钵土面上保持水层,使土壤与外源铜平衡。各处理土壤供肥(N、P2O5、K2O)水平相同,播种前1 d 每盆基肥施纯N 0.24 g、P2O50.19 g、K2O 0.33 g 及相对应的硅肥于盆中与土搅拌均匀,硫磺施入与表层土混合均匀。第2 天每盆摆播已露白种子150 颗并塌谷。发芽前保持湿润,出芽保持浅水层。一叶一心时每盆追施纯N 0.1 g。均以不施硫磺和硅酸钠处理为对照,随机排列,所用肥料分别为尿素、钙镁磷肥和氯化钾,其他管理措施一致,灌溉水用自来水。
1.3 测定指标与方法
(1)水稻幼苗50 d后取样素质考查及生物量的测定[22]。(2)水稻幼苗生长50 d后取样用自来水冲洗泥土,再用蒸馏水冲洗2次,分根、茎叶两部分,先在105 ℃恒温箱中杀青0.5 h,再在70 ℃条件下烘干称量。(3)叶片丙二醛(MDA)、过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)测定[23-24]:每处理取混合样100 株左右,用湿纱布包裹放入冰水中,立即送实验室处理——用蒸馏水稍微冲洗,再用干纱布擦干,取相对大、宽的叶片,一般去除主脉(若苗太小,则可不除),剪除靠茎端,再把剩余叶片剪成1~2 cm 小段于小烧杯中,立刻用锡纸、捏子称样,每份1 g或0.5 g,5~6份备用。用记号笔标记,立刻用液氮速冻,再用封口袋装好,写好标签,放入超低温(-70 ℃)冰箱保存,用于丙二醛(MDA)、过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)以及可溶性蛋白质(Pr)、可溶性糖含量的测定。具体测定方法:MDA 含量用TAB 法测定并计算出可溶性糖含量、POD 活性用愈创木酚法测定、CAT 活性测定用消光系数法、SOD 活性测定用氮蓝四唑光化还原法、Pr含量采用考马斯亮蓝法测定。(4)非蛋白质巯基总量(NPT)、谷胱甘肽(GSH)、植物螯合肽(PCs)和硫蛋白(MT)含量的测定:非蛋白质巯基总量(NPT)的测定参照AshRaft 等[25]的方法,称取1 g 鲜样,加入预冷的5%(V/V)SSA(含6.3 mmol/L DTPA,pH<1)3 mL和少量石英砂冰浴上充分研磨,4 ℃低温离心(10 000 r/min)20 min,取上清液0.25 mL加0.5 mL 0.01 mol/L磷酸缓冲液(pH7.5)和3 mL 10 mmol/L 的DTNB,室温下放置20 min,在412 nm 波长下用分光光度计比色测定;GSH的测定则参照赵旭东等[26]的方法,称取1 g鲜样,需加入5 mL 5%的TCA于冰浴中研磨,离心(16 000 r/min)20 min,所得上清液加入1.5 mL 0.1 mmol/L NaOH 调节pH值为6~8后加入反应液。反应液中含0.5 mL水,2.0 mL提取液,0.5 mL 0.2 mol/L磷酸钾缓冲液(pH 7.0),0.1 mL DTNB(75.3 mg溶于30 mL 0.1 mol/L 缓冲液)以及3%甲醛(pH 8.0)1 mL,以缓冲液代替DTNB 作空白,于波长412 nm 下比色测定。根据同样程序制作标准曲线,植物螯合肽(PCs)的测定采用差减法,即PCs=非蛋白质态巯基总量(NPT)—谷胱甘肽含量(GSH)。
金属硫蛋白(MT)的测定取1 g鲜样于研钵中,加入0.01 mol/L,pH 为8.6的Tris-HCl 缓冲液10 mL 充分研磨,后于4 ℃冰箱过夜抽取。抽提液离心(4 ℃,10 000 r/min)30 min,收集上清液并于100 ℃水浴加热2~3 min,重复离心20 min,收集上清液。加入3 倍体积-20 ℃预冷的无水乙醇,于-20 ℃过夜沉淀后,离心(4 ℃,12 000 r/min)20 min。取沉淀加入0.01 mol/L Tris-HCl缓冲液5 mL,溶解数小时后,离心(4 ℃,15 000 r/min)20 min,收集上清即为MT提取液。取提取液1 mL用Cd-Hb饱和法测定MT含量[27-28]。
1.4 数据处理与分析
应用Excel软件计算分析,应用Origin Pro 7.0软件作图。应用DPS软件进行数据方差分析(Duncan’s法)。
2 结果与分析
2.1 硫硅配施对铜污染下水稻幼苗生长的影响
2.1.1 硫硅配施对铜污染下水稻幼苗苗高的影响 由图1 可看出,硫肥施用能显著促进苗的生长,但与硫肥用量密切有关,其中S1苗高平均值小于对照,但与对照差异不显著(P>0.01),S2苗高平均值显著高于对照和S1(P<0.05),增幅分别为4.86%和6.80%。施用硅肥与对照相比苗高增加极显著(P<0.01),且苗高随着硅肥用量的增加而增加,增幅为21.97%~26.42%,但Si1和Si2间苗高差异不显著。因硫硅肥互作对苗高影响不明显,故硫、硅肥配施以S2Si2效果最佳,比对照苗高增加了37.03%。相关分析表明,苗高与单株根、茎叶干质量呈极显著正相关,相关系数分别为r=0.830**、r=0.894**、与根、茎叶铜含量呈显著、极显著负相关,相关系数分别为r=-0.752*和r=-0.851**。
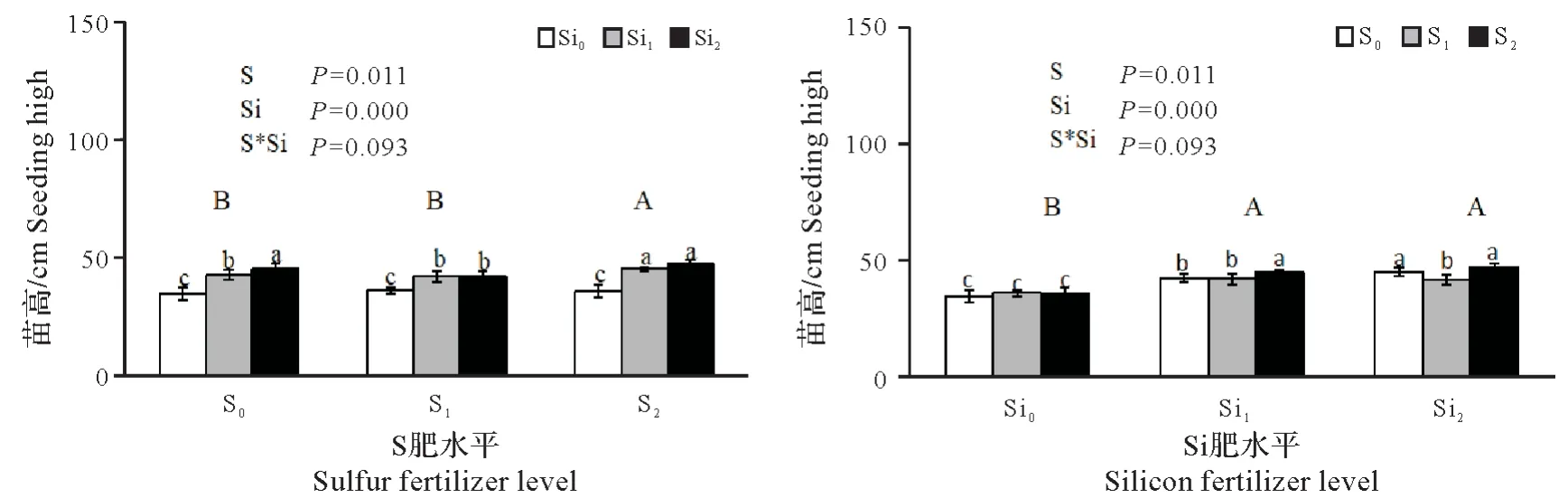
图1 硫硅配施对铜污染下水稻幼苗苗高的影响Fig.1 Effects of combined application of sulfur and silicon on rice seedling height under copper contamination
2.1.2 硫硅配施对铜污染下水稻幼苗叶龄和绿叶数的影响 由图2可看出,单施硫肥对水稻出叶速率及绿叶数均有一定影响,且与硫肥用量密切有关。其中除S2叶龄平均值小于对照,但与对照差异不显著(P>0.05),S1叶龄平均值显著高于对照和S2(P<0.05),增幅分别为6.54%和7.24%。施用硅肥与对照相比出叶速率明显提高,差异极显著(P<0.01),增幅大致为7.59%~15.17%,Si1和Si2间叶龄差异显著。此外,硫肥的施用绿叶数平均值均小于对照,但绿叶数S1、S2与对照之间无显著差异(P>0.05)。施用硅肥与对照相比绿叶数明显增多,存在极显著差异(P<0.01),增幅为16.13%~19.35%。而硫硅肥互作对叶龄及绿叶数影响均极显著(P<0.01),对于提高出叶速率和增加绿叶数,硫、硅肥配施均以S2Si1效果最佳,相比于对照,出叶速率提高了约31.11%,绿叶数增加了30%。
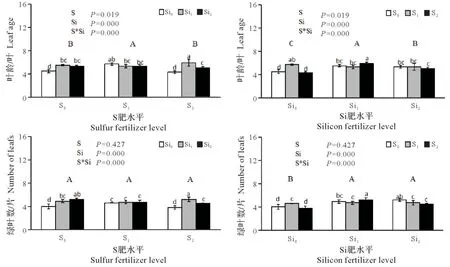
图2 硫硅配施对铜污染下水稻幼苗叶龄和绿叶数的影响Fig.2 Effects of combined application of sulfur and silicon on leaf age and green leaf number of rice seedlings under copper contamination
2.1.3 硫硅配施对铜污染下水稻幼苗茎基宽的影响 由图3 可看出,施用硫肥能明显促进茎部的生长,与对照相比,茎基宽增加极显著(P<0.01),施用S1,S2的茎基宽平均值均高于对照,增幅分别为7.54%和23.02%,S1和S2间的茎基宽也存在显著差异。施用硅肥与对照相比茎基宽增加极显著(P<0.01),增幅为28.38%~35.38%,但Si1和Si2间茎基宽差异不显著。硫硅肥互作对茎基宽影响差异极显著(P<0.01),其中硫、硅肥配施以S2Si2效果最佳,比对照茎基宽增加了116.36%。
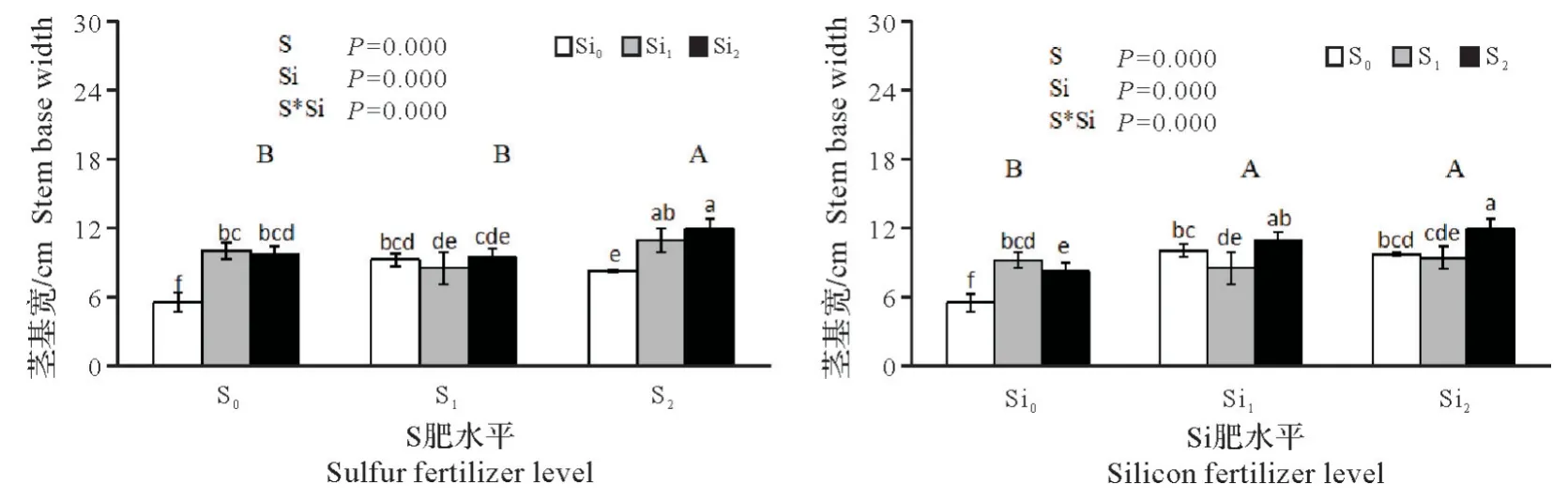
图3 硫硅配施对铜污染下水稻幼苗茎基宽的影响Fig.3 Effects of combined application of sulfur and silicon on stem base width of rice seedlings under copper contamination
2.1.4 硫硅配施对铜污染下水稻幼苗根系的影响 由图4可看出,施用硫肥对幼苗总根数有明显促进作用,且与硫肥用量密切相关,其中S2总根数平均值略高于对照,而S1总根数平均值则显著高于对照和S2,增幅分别为54.81%和18.21%。不同用量硫肥对幼苗最长根长的影响与对照相比,均存在显著差异,S1、S2的最长根长均值都小于对照。施用硅肥能显著影响幼苗总根数(P<0.01),且Si1和Si2不同用量的效果之间存在明显差异:相比于对照,Si1幼苗总根数有所增加,而Si2处理下总根数大大减少,显著低于对照与Si1处理。施用硅肥对水稻幼苗最长根长的影响极为显著,且根长随硅肥用量的增加而增加,增幅为28.38%~35.37%,且Si1和Si2间最长根长也存在明显差异。硫硅肥配施对总根数和最长根长均有显著影响,其中单施Si1对总根数的增加效果最佳,比对照增加了180.56%,对根长影响以S2Si2配施效果最为明显,比对照根长增加了177.46%。
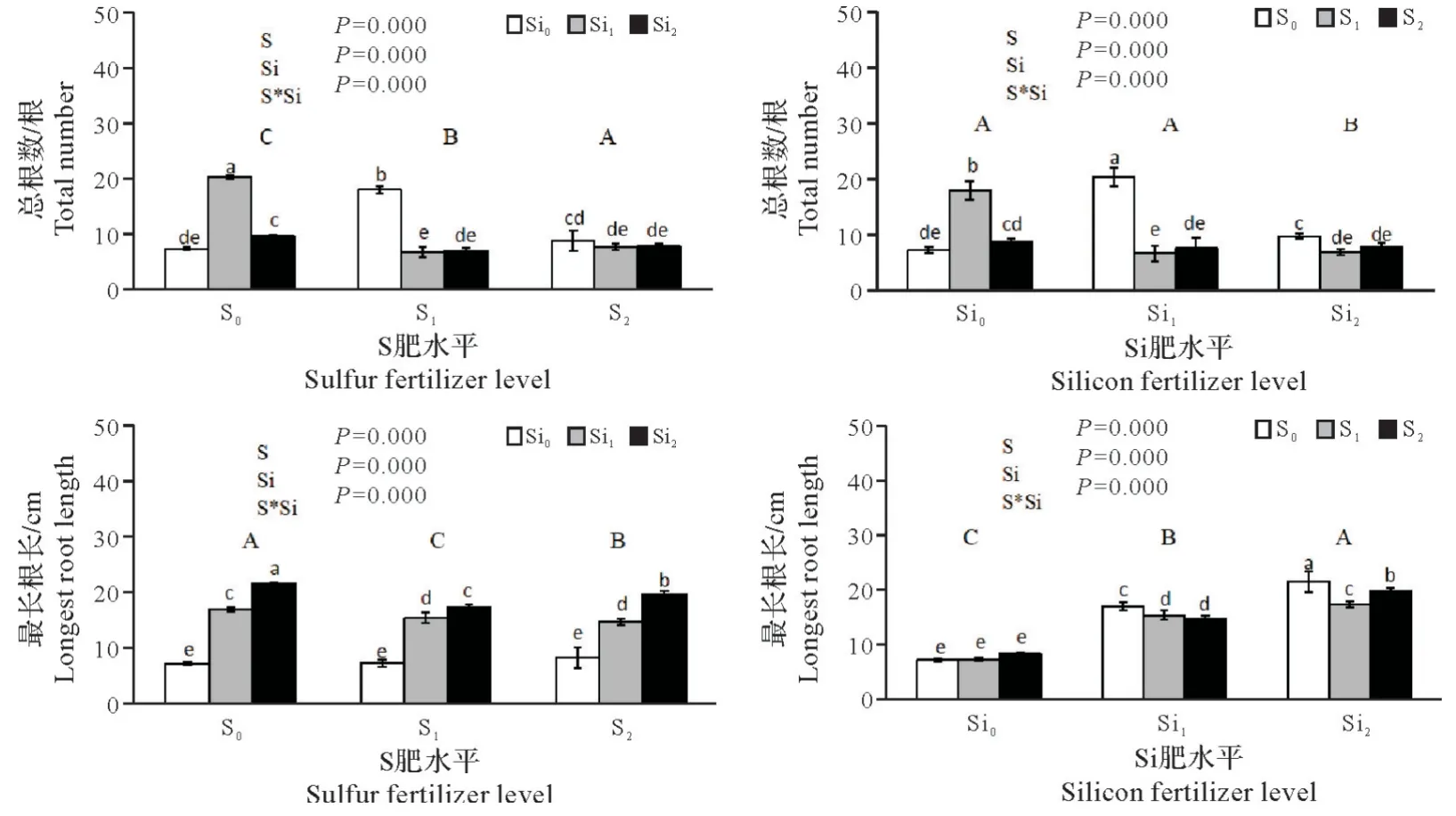
图4 硫硅配施对铜污染下水稻幼苗根系的影响Fig.4 Effects of combined application of sulfur and silicon on root system of rice seedlings under copper contamination
2.1.5 硫硅配施对铜污染下水稻生物量的影响 由表2可知,施用硫肥与对照相比可显著增加地下部分的质量,增幅在40.12%~65.27%,S1和S2平均值之间则无明显差异。此外,施用硫肥对地上部分的质量无明显影响,S1平均值与对照相比略有增加,S2平均值与对照相比略有减少,均与对照差异不显著(P>0.05),综合看来,施用硫肥对根冠比有显著影响,但与硫肥用量有关,S1根冠比平均值与对照差异不明显,S2平均值则显著高于对照和S1,增幅达64.29%。施用硅肥对单株干物质量和根冠比均有极显著影响,有明显促进作用。施用硅肥情况下,Si1和Si2的地下部分平均值均高于对照,增幅分别为168.26%和403.59%,Si1和Si2的均值间也存在显著差异(P>0.05)。施用硅肥地上部分的平均值也均高于对照,Si1和Si2的平均值相比对照,增幅分别为40.20%和64.30%,Si2的地上平均值略高于Si1,但无明显差异(P<0.01)。此外,施用硅肥还可显著提高根冠比,Si1和Si2的根冠比均值相比对照,增幅分别为85.71%和192.86,且Si1和Si2的根冠比均值之间也存在显著差异(P<0.01)。硫硅肥配施对单株整体干物质量有明显促进作用,以S2Si2效果最为显著,地下部分增幅可达330.54%,地上部分增幅可达90.60%,而S1Si2对根冠比的改善效果最佳,增幅为192.86%。
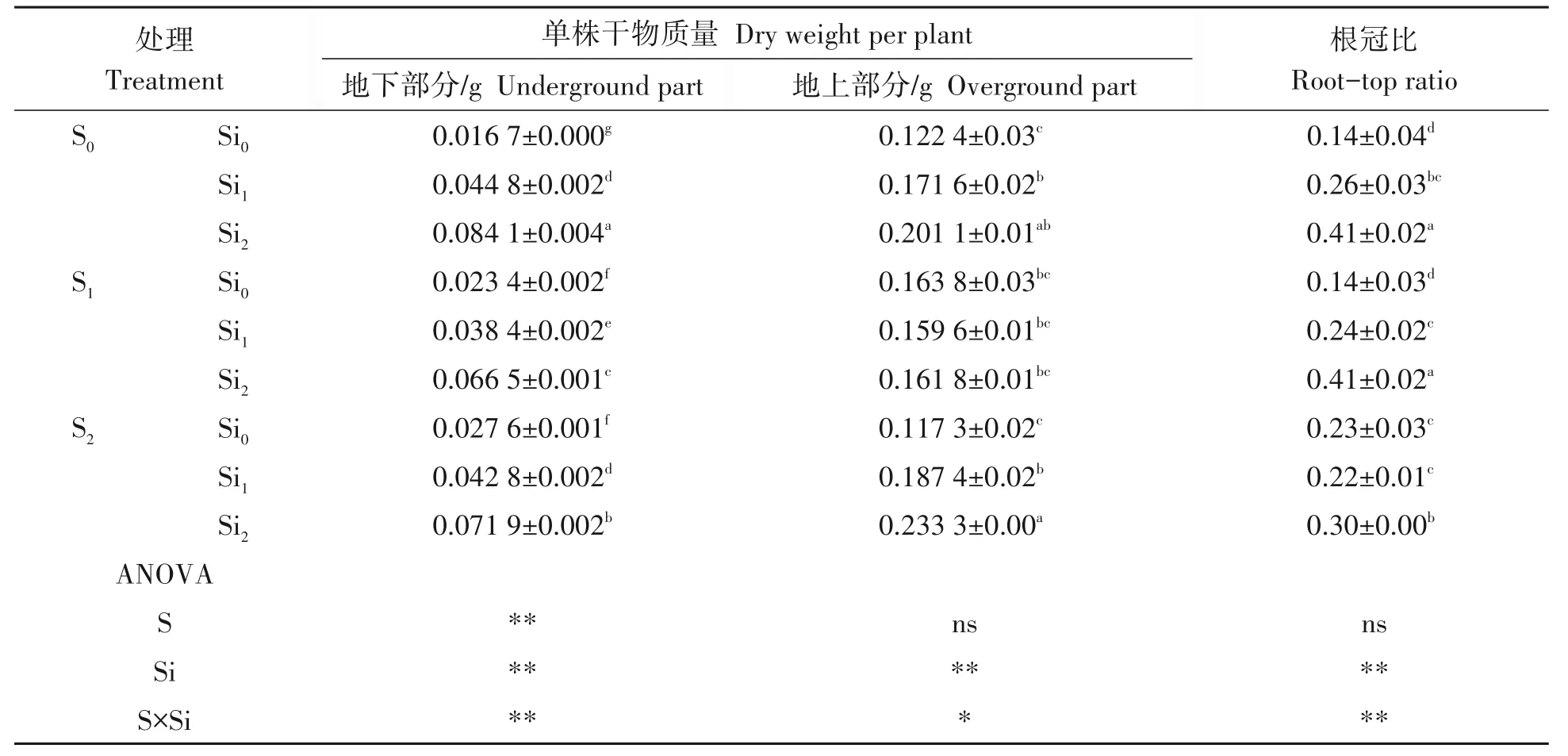
表2 硫硅配施对铜污染下水稻幼苗生物量的影响Tab.2 Effect of sulfur and silicon on rice seedling biomass under copper contamination
2.2 硫硅肥配施对铜污染下水稻幼苗叶片膜脂质过氧化作用的影响
由表3 可知,与对照相比,单施硅肥能一定程度减少膜脂质过氧化反应的产物丙二醛(MDA)含量,但与其用量密切相关,MDA 含量S2的均值略高于对照,但差异不明显(P>0.05),而S1含量均值显著低于对照和S2,减幅分别为15.53%和20.18%。施用硅肥则能显著减少MDA 含量(P<0.01),且不同用量的效果之间无显著差异,Si1和Si2的平均值相比对照降幅分别为30.42%和28.48%。硫硅肥配施同对照相比均能显著减少MDA 含量,但配施各处理间均无显著差异(P>0.05),降幅大致在20%左右,以S2Si2效果最佳,降幅达26.12%。
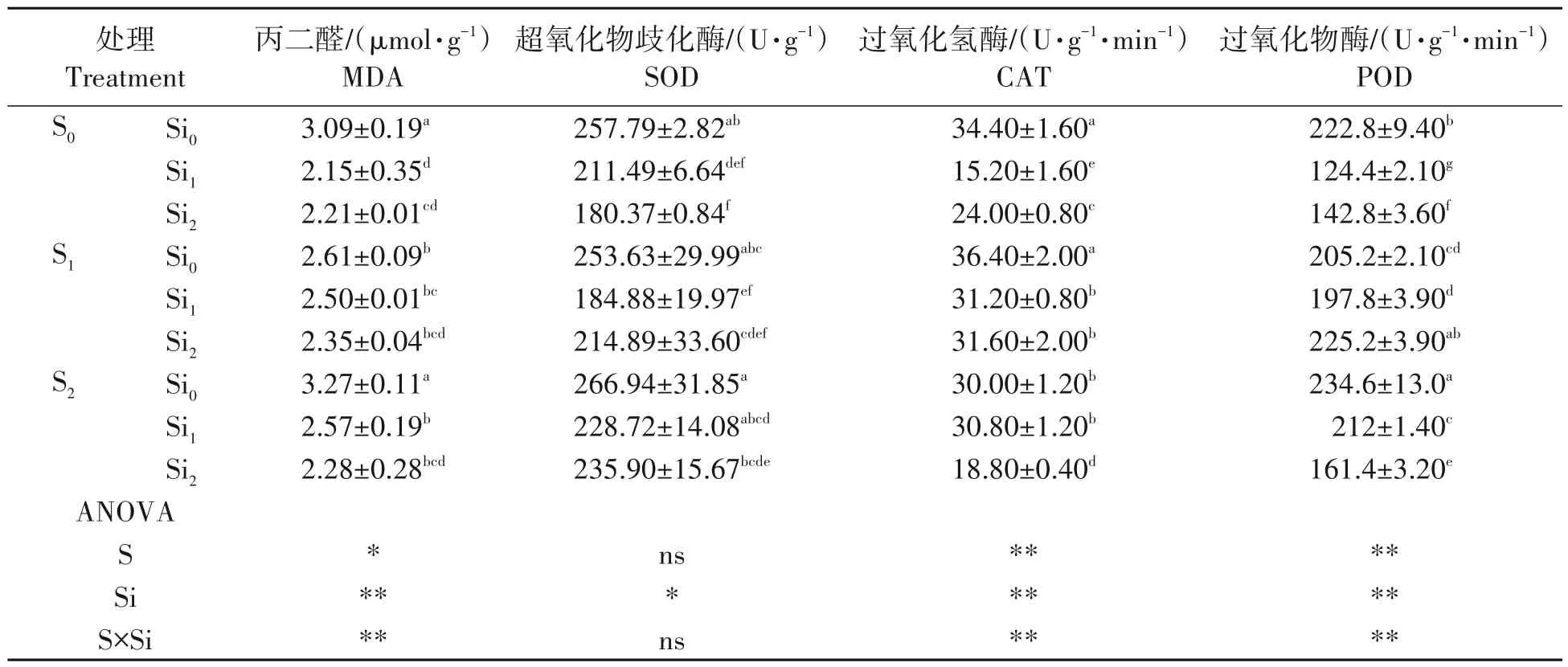
表3 硫、硅肥配施对铜污染下水稻幼苗MDA及抗氧化保护酶的影响Tab.3 Effect of sulfur and silicon on the MDA and antioxidant enzymes of rice seedling under copper contamination
2.3 硫、硅肥配施对铜污染下水稻幼苗抗氧化保护酶的影响
2.3.1 对水稻幼苗叶片超氧化物歧化酶(SOD)活性的影响 由表3可知,硫肥的施用对SOD活性无显著影响(P>0.05),而施用硅肥能显著降低SOD 活性(P<0.01),但与用量无密切联系,SOD 活性Si1和Si2与对照相比,分别降低了17.96%和30.03%。硫硅肥配施与对照相比SOD 活性降低不显著,降低活性效果最好的为S1Si1,与对照相比,下降了28.28%。
2.3.2 对水稻幼苗叶片过氧化氢酶(CAT)的影响 由表3 可以看出,施用硫肥能显著降低过氧化氢酶(CAT)活性,但与硫肥用量相关,S1的活性均值略高于对照,但无显著差异(P>0.05),S2活性均值显著低于对照和S1(P<0.05),降幅分别为12.79%和17.58%。施用硅肥水稻幼苗叶片中CAT 活性也会显著降低(P<0.01),Si2活性均值与对照相比,下降了30.23%,Si1活性显著低于对照和Si2,降幅分别为55.81%和36.67%。硫硅肥配施均显著降低CAT活性,以S2Si2效果最佳,降幅达45.35%。
2.3.3 对水稻幼苗叶片过氧化物酶(POD)的影响 由表3 可知,不同用量硫肥对POD 活性的影响有所不同,但均与对照存在显著差异,S1活性均值低于对照,降幅在7.90%,差异显著,而S2活性均值高于对照,增幅在5.30%,存在显著差异。施用硅肥可显著降低POD 活性,Si2活性均值比对照低,降幅在35.91%,Si1活性显著低于对照和Si2,降幅分别为44.17%和12.89%。硫硅肥配施对POD 活性影响与对照相比,显著降低,活性降低效果最为显著的为S2Si2,降幅可达27.56%。
2.4 硫、硅肥配施对铜污染下水稻幼苗非蛋白质态巯基化合物含量的影响
由表4可知,施用硫肥或硅肥,或硫硅肥配施对非蛋白质态巯基总量(NPT)含量无显著影响,各施肥处理与对照相比,NPT 含量均无显著差异。单施硫肥或硅肥,与对照相比,谷胱甘肽(GSH)含量无显著差异,而硫硅肥配施下GSH 含量略有下降,但差异不明显,以S1Si1效果最为明显,降幅为30.45%。此外,施用硫肥可明显提高植物螯合肽(PCs)含量,S1和S2含量均值与对照相比存在极显著差异,增幅分别为760.96%和183.56%。单施硅肥则对PCs含量无显著影响,与对照相比,差异不明显。硫硅肥配施与对照相比极显著地提高了PCs含量,以S1Si1效果最佳,差异最为显著。
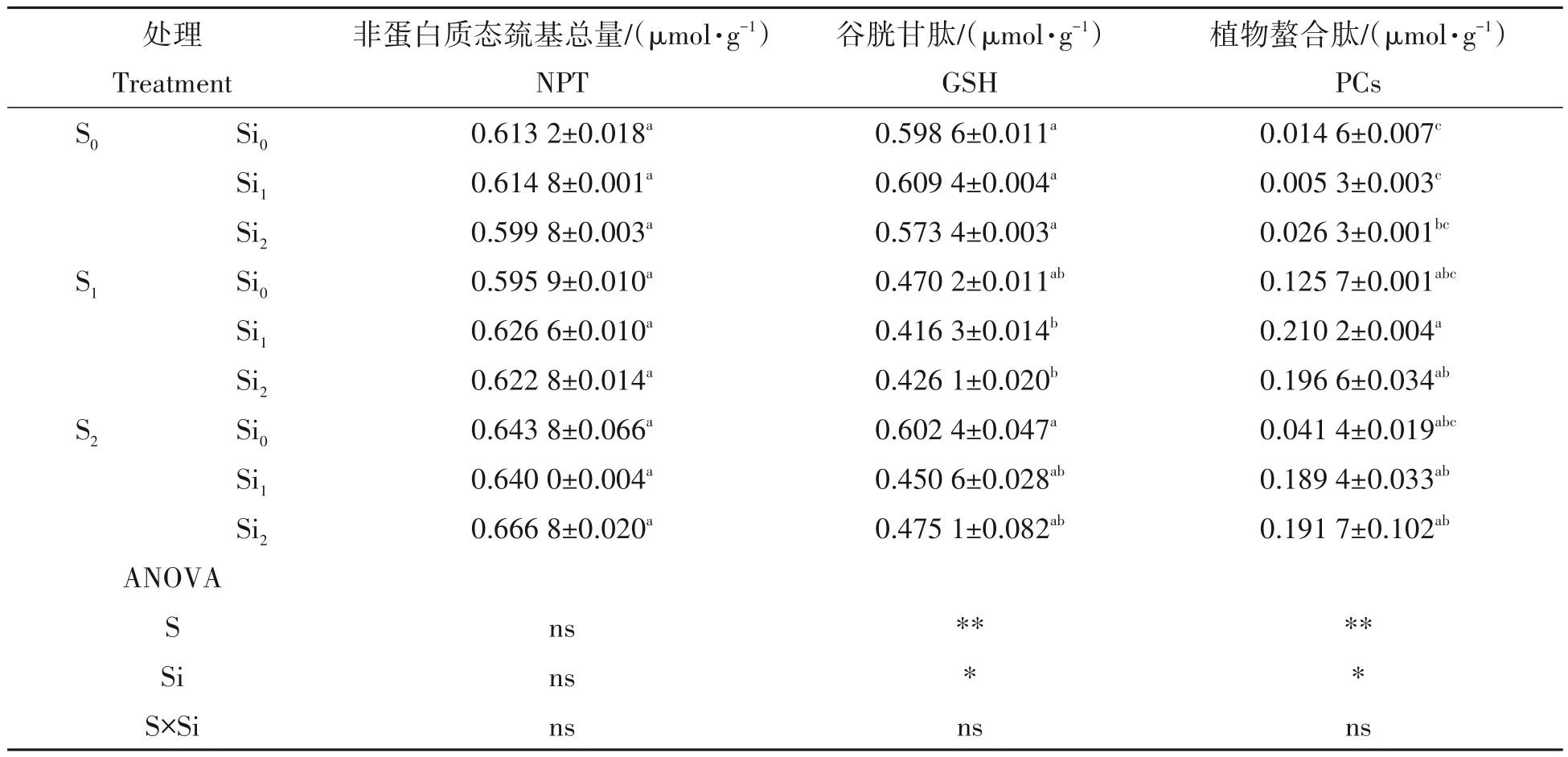
表4 硫、硅肥配施对铜污染下水稻幼苗非蛋白质态巯基化合物含量的影响Tab.4 Effects of combined application of sulfur and silicon fertilizers on the content of non protein sulfhydryl compounds in rice seedlings under copper contamination
2.5 硫、硅肥配施对铜污染下水稻幼苗金属硫蛋白含量的影响
由图5可知,施用硫肥对水稻类金属硫蛋白酶含量的有一定的提高,且与硫肥用量有一定关系,S1的平均值小于对照,但无显著差异。S2平均值极显著高于对照和S1(P<0.01),增幅分别为68.24%和116.09%。施用硅肥对类金属硫蛋白含量也有明显提高,Si2与对照和Si1相比均存在显著差异,且Si1的平均值显著高于对照和Si2,增幅分别为34.49%和24.11%。不同用量的硫硅肥配施对类金属硫蛋白的含量有不同影响,其中,S1Si2配施能有效减少类金属硫蛋白含量,比对照减少了19.46%。
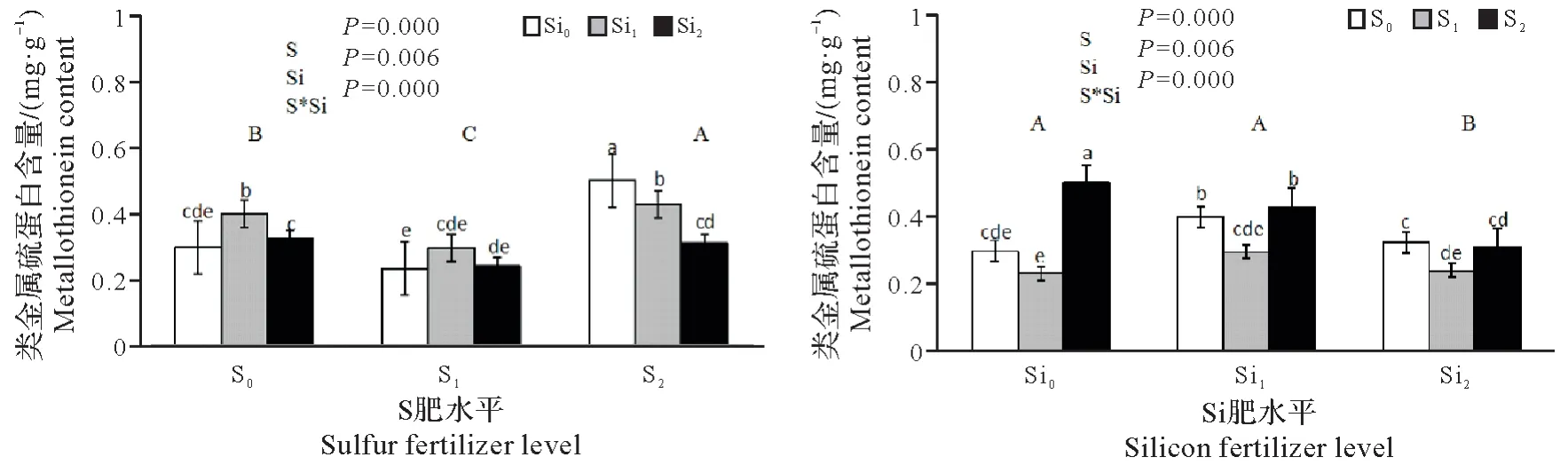
图5 硫、硅肥配施对铜污染下水稻幼苗金属硫蛋白含量的影响Fig.5 Effect of sulfur and silica fertilizer on metallothionein content of rice seedlings under copper contamination
2.6 硫硅肥配施对铜污染下水稻幼苗不同部位铜含量的影响
由表5可看出,硫肥与硅肥的施用都会显著影响水稻幼苗不同部分的铜含量。施用硫肥会显著减少水稻幼苗地上部分茎叶的铜含量,不同用量之间存在一定差异,S2含量比对照下降了8.91%,S1含量比对照下降了23.67%,比S2下降了16.21%,差异显著。施用硅肥的茎叶铜含量与对照相比大幅下降,差异极显著,且随硅肥用量的增加茎叶铜含量减少越明显,Si1茎叶铜含量比对照减少了25.42%,Si2茎叶铜含量比对照和Si1铜含量分别减少了69.82%和59.54%。硫硅肥配施极显著地减少了茎叶中铜含量,最为显著的是S1Si2,与对照相比减少了64.77%。

表5 硫硅配施对铜污染下水稻幼苗铜含量的影响Tab.5 Effect of combined application of sulfur and silicon fertilizers on the copper content of rice seedlings under copper contamination
同时,还可看出,施用硫肥可明显提高幼苗根部的铜含量,S1和S2根部铜含量与对照相比分别增加了30.31%和26.62%,差异显著。而施用硅肥与对照相比则是减少根部铜含量,Si1和Si2分别比对照减少了15.64和2.48%。硫硅肥配施也可明显降低根部铜含量,以S2Si1效果最为显著,比对照下降了47.11%。
2.7 水稻幼苗不同部位铜含量与抗氧化酶含量、非蛋白巯基化合物含量和金属硫蛋白含量的相关性
相关分析表明(表6),根质量与茎叶质量呈显著正相关,相关系数为r=0.769*,与茎叶铜含量呈极显著负相关,相关系数为r=-0.907**,茎叶质量与茎叶铜含量呈显著负相关,相关系数为r=-0.773*。MDA含量与根质量、茎叶质量呈显著负相关,相关系数分别为r=-0.745*、r=-0.797*,与茎叶铜含量呈显著正相关,相关系数为r=0.765*,SOD 也与茎叶铜含量呈显著正相关,相关系数为r=0.715*。除此之外抗氧化酶类SOD、CAT、POD 活性均与根质量和茎叶质量呈负相关,与铜含量和茎叶铜含量呈正相关。NPT 和MT均与根质量呈正相关、与茎叶质量及各部分铜含量呈负相关。GSH与各部位质量呈负相关,与各部位铜含量呈正相关,而PC刚好与之相反,与各部位质量呈正相关,与各部位铜含量呈负相关。
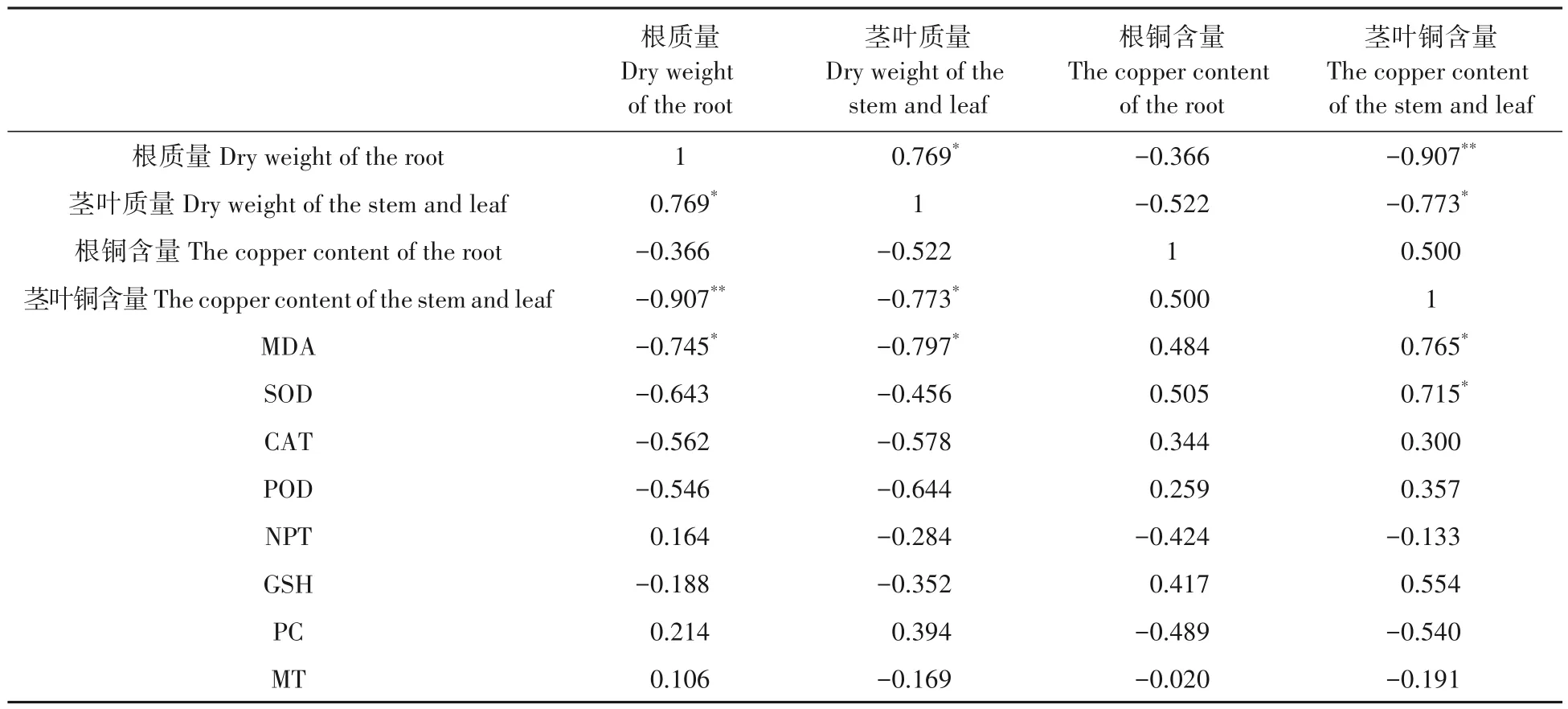
表6 水稻不同部位铜含量与抗氧化酶含量、相关非蛋白巯基化合物含量和金属硫蛋白含量的相关矩阵Tab.6 Correlation matrix of copper content in different parts of rice with antioxidant enzyme content,related non-protein sulfhydryl compounds content and metallothionein content
3 讨论与结论
水稻幼苗的基本素质直接反映了水稻幼苗的生长情况[29-30]。综合该试验结果显示,铜污染下单施硫肥:低硫处理的水稻幼苗基本素质相比高硫处理,壮苗效果相对较好,可能是硫肥的施用方式为蘸秧根。张延青等[31]研究表明,在水稻幼苗生长初期,施入的硫元素过量易在根部土壤氧化为SO42-,致使土壤pH值下降,造成一定的酸害,除此之外,短时间内还会影响土壤中铜的有效性,导致铜毒害加重。水稻本身就是喜硅作物,铜污染下单施硅肥的情况下,壮苗效果随硅肥用量的增加而更为明显。硅肥施入土壤根际pH 值升高,且施入土壤中所形成的SiO32-会与Cd、Pb 等重金属结合,通过与重金属离子结合形成更为稳定的硅酸化合物,从而降低重金属的有效性,缓解重金属毒害[10,19]。而不同用量的硫肥与硅肥配施所呈现的壮苗效果略有差异。
随着硫肥、硅肥的施加,铜污染下水稻幼苗的生长情况也会得到极大的改善。水稻幼苗苗高、茎基宽、根系的生长和单株干质量等以浓度为0.026 g/kg硫肥与浓度为0.1 g/kg的硅肥配施改善效果最佳,而水稻幼苗的出叶速率及绿叶数和根冠比则以浓度为0.026 g/kg 硫肥与浓度为0.05 g/kg 的硅肥配施效果最佳,说明在一定浓度范围内,较高浓度硫肥与硅肥配施能更好地提高水稻干物质量。其原因可能是硫元素作为叶绿素合成的必需元素,影响着植物体的光合作用,施用较高浓度的硫肥可显著增强叶片的光合能力,从而增加叶片干物质量的积累,这与耿计彪等[32]、卢志红等[33]研究结果相一致。同时,前人[4,10]研究表明铜元素也参与了植物体的光合作用,单施较高浓度0.1 g/kg的硅肥的情况下,降低土壤中过高的有效铜含量,而土壤中适量的铜含量有助于加强植物光合作用等生理代谢活动,故试验结果显示0.026 g/kg硫肥和浓度为0.1 g/kg的硅肥配施与单施较高浓度0.1 g/kg硅肥的地上部干质量差异不显著。
丙二醛(MDA)是细胞膜脂过氧化的标志性产物,其含量高低反映细胞脂质过氧化程度和植物对逆境条件下反应的强弱[34-35]。本研究结果表明,随着硫硅肥的施加,MDA 含量均有着不同程度的减少,浓度为0.026 g/kg 硫肥与浓度为0.1 g/kg 的硅肥配施所降MDA 含量可达26%,这说明硫硅肥配施可减轻铜胁迫下水稻幼苗膜脂过氧化程度,使水稻幼苗细胞膜损伤减少。此外,为抵抗铜胁迫对植株所造成的危害,植株体内抗氧化酶类(SOD、CAT、POD 等)活性有所加强,而本研究结果显示,在硫硅肥配施条件下,此类抗氧化酶类活性均有不同程度的下降,说明施肥后水稻幼苗所遭受的铜胁迫有所减轻。这与蔡德龙等[36]研究结果随细胞膜损伤减轻,相对应的抗氧化酶类活性随之下降相一致。
谷胱甘肽(GSH)、植物螯合肽(PCs)、金属硫蛋白(MT)等非蛋白巯基化合物(NPT)在植物中的含量可以为重金属毒性耐受和解毒机制提供相关信息。GSH 作为一种重要的非酶类抗氧化剂,在重金属污染所导致的氧化胁迫方面起着非常重要的抵御作用,而植物络合肽(PCs)则是在植物螯合肽合成酶(PCS)的参与下,由GSH 转化合成而来,能与多种金属离子结合形成大分子络合物,使植物难以吸收,从而达到减轻重金属对植物的毒害作用[37-38]。金属硫蛋白(MT)是一类富含巯基的低分子量蛋白,在植物的重金属解毒及细胞氧化还原调控等方面也起着重要的作用。本研究结果显示,铜胁迫下,施加硫硅肥,NPT 总量基本没有很大改变,植物体内GSH 含量下降的同时PC 和MT 的含量有所增加。除此之外,该研究结果还表明,硫硅肥配施有利于减少植株整体的铜含量,很大原因可能是硫肥的施加加快了铜元素的转移易位[39],同时硅肥又提高了植株的代谢水平[40],故整体铜含量均有所下降。
本研究硫硅肥配施缓解铜对水稻幼苗毒害所造成的氧化胁迫,有效减少了MDA 含量,同时抗氧化酶类活性(SOD、CAT、POD 等)也有所下降。铜胁迫下,硫硅肥配施对非蛋白质态巯基总量(NPT)无明显影响,但在一定程度上会降低GSH含量,而PC和MT含量有所增加;施用硫、硅肥能明显减少水稻地上部分铜含量并提高水稻根冠比,其中以施用量为0.026 g/kg的S与0.1 g/kg的Si配施壮苗效果较为理想。
