养殖军曹鱼稚鱼骨骼畸形研究*
2022-01-14毛非凡周启苓黄建盛邝杰华
毛非凡 陈 刚 马 骞 周启苓 施 钢 黄建盛 邝杰华
(广东海洋大学水产学院 广东 湛江 524025)
骨骼是鱼类支持身体、保护柔软器官的重要结构,在海水鱼类从初孵仔鱼发育至成鱼的过程中,骨骼发育起到至关重要的作用,遗传因素、环境因素以及营养因素等均可影响骨骼发育导致骨骼畸形(Cahu et al, 2003)。骨骼畸形会导致仔、稚鱼摄食能力低,运动能力弱,生长缓慢,成活率低等问题(Gapasin et al,2001)。在人工繁育过程中,畸形鱼苗成长至商品鱼规格后,也会因为品相不佳无法销售带来直接经济损失(Cobcroft et al, 2012)。目前,对鱼类骨骼畸形的研究多集中在鲈形目(Perciformes) (Ma et al, 2016; Cobcroft et al, 2004; Wang et al, 2000; 温久福等, 2020; Fraser et al, 2005; Nagano et al, 2007; 李远友等, 2019)、鲽形目(Pleuronectiformes) (Hosoya et al, 1998; Martinez et al, 2003; Lewis et al, 2004; Gavaia et al, 2002; 马慧等, 2011)、鲤形目(Cypriniformes) (Ferreri et al, 2000)和鲑形目(Salmoniformes) (Witten et al, 2016)中的部分经济鱼类,研究内容主要包括骨骼畸形类型的鉴定和发生过程(Ferreri et al, 2000),以及遗传、环境和营养等因素对鱼类畸形率及死亡率的影响(Alix et al,2017; Fraser et al, 2019; Kuroki et al, 2016),其中,阐明骨骼畸形的类型及发生过程是研究基础和关键内容。
军曹鱼(Rachycentron canadum)隶属于鲈形目、鲈亚目(Percoidei)、军曹鱼科(Rachycentridae)、军曹鱼属(Rachycentron),为外海暖水性经济鱼类,主要分布于大西洋、印度洋、太平洋(东太平洋除外)等热带水域,在中国南海部分海域亦有少量分布。军曹鱼生长速度快,环境适应能力强,养殖周期短,经济价值高,近年在南方沿海的人工养殖规模不断扩大(陈刚等, 2004; 陈浩如等, 2006)。本研究团队在开展军曹鱼人工繁育研究中,观察到15%~20%的鱼苗存在畸形。军曹鱼作为极具潜力的养殖品种,控制畸形率是保障养殖利润的前提之一,但有关军曹鱼的骨骼发育的研究尚未见报道。
硬骨鱼类中,严重骨骼畸形的个体通常不能正常发育乃至存活,稚幼鱼在骨骼数量和形态等方面均已接近成鱼,骨骼已趋于完全骨化状态,这一发育阶段的骨骼畸形类型可能会在成鱼中延续。故本研究选取同一批次孵化的 25日龄(days post hatching, dph)军曹鱼稚鱼,通过采取软骨—硬骨双染色方法进行全骨骼染色,对其骨骼畸形发生部位、畸形种类和畸形率进行研究,为研究军曹鱼骨骼畸形的成因,降低畸形率及军曹鱼人工养殖的推广提供参考资料。
1 材料方法
1.1 仔稚鱼的培育
2020年4月,在广东海洋大学生物研究基地(东海岛)进行了军曹鱼人工繁育。受精卵收集分离筛选后置于500 L圆形孵化桶中充气孵化。仔鱼孵出后,将其从孵化桶转移至室内水泥池(5 m×3.5 m×1.5 m)进行培育。种苗培育过程投喂饵料和饲料:3~10 dph投喂轮虫(Rotifer),投喂量保持在 1~4 ind./mL;8~19 dph投喂桡足类(Copepods),投喂量保持在1~3 ind./mL;10~23 dph投喂卤虫(Artemia saline)幼体,投喂量保持在 1~3 ind./mL;17~30 dph 投喂越群小苗 3#~6#料(成分组成:蛋白质55.2%、粗脂肪12.7%、粗纤维1.0%、粗灰分12.2%、水分6.1%、赖氨酸3.5%),投喂量保持在体重的8%~10%,3~17 dph投喂 3~4次/d,17~30 dph 投喂 2~3次/d。
1.2 实验样品采集
选取同一孵化批次、25 dph军曹鱼稚鱼为研究对象,取样前一天停止投喂。稚鱼经丁香酚麻醉后,再置于 4%多聚甲醛固定液中于4℃充分固定,之后转至70%酒精溶液中(-20℃)保存。
1.3 骨骼染色
通过对Guido等(1977)的软骨—硬骨双染色方法并加以改进,采用1% KOH溶液对样品进行漂白脱色,对军曹鱼稚鱼全骨骼进行染色,骨骼染色具体步骤如下:
清洗:将经4%多聚甲醛固定的稚鱼放入ddH2O中浸泡 1~2 d。
软骨染色:将清洗过的标本置于0.02%阿尔新兰染液中12 h,直至观察到鳍基部有明显蓝色。
梯度脱水:将标本依次放入95%、75%、40%、15%的酒精溶液和ddH2O中进行梯度脱水,每个梯度2 h。
透明化:将标本放入 1% KOH溶液中进行透明化,直至标本完全透明。
硬骨染色:将标本放入0.1%茜素红染液中约45 min,直至硬骨染上红色。
保存:将标本依次移入0.5% KOH和甘油体积比为(3∶1、1∶1和1∶3)的混合液中,脱去多余颜色,最终移入甘油中,并加入3~4粒廖香草酚保存。
1.4 实验标本的观察
采用游标卡尺测量透明样本全长,用Leica M205 FCA体视显微镜对透明标本进行观察拍照,记录骨骼畸形的发生部位和形态特征,最后进行资料整理归类。骨骼命名参照Harder (1975),骨骼畸形的归类参照Favaloro等(2006)。
1.5 数据处理
对骨骼正常组与骨骼畸形组鱼体全长(TL)数据进行统计分析,所得数据以平均值±标准差(Mean±SD)表示。利用统计学软件SPSS 20.0进行单因素方差分析(one-way ANOVA),分析骨骼正常组与骨骼畸形组之间鱼体全长的差异水平;利用单因素方差分析和Duncan’s多重比较,分析骨骼正常组、各不同畸形部位组之间鱼体全长的差异水平。畸形率计算公式:
畸形率=(畸形数/总数)×100%
2 结果
180个军曹鱼稚鱼骨骼样本的总畸形率为40.00%,共存在22种骨骼畸形类型。骨骼畸形发生部位主要为颅骨(cranium)、脊柱(vertebral)、背鳍(dorsal fin)、臀鳍(anal fin)、尾鳍(caudal fin)。具体骨骼畸形的发生部位、畸形类型和畸形率见表1。其中,62个样本存在1种骨骼畸形,10个样本存在1种以上的骨骼畸形。

表1 军曹鱼稚鱼骨骼畸形种类与畸形率Tab.1 Type and incidence of deformities in juvenile R. canadum
军曹鱼稚鱼颅骨畸形个体 TL=(43.14±7.13) mm,畸形率为17.22%,共发现米克尔氏软骨畸形(MCD,图1A)、基舌骨畸形(BD, 图1B)、基舌骨异位(BA,图1C)和舌弓下沉(LHA, 图1D) 4种骨骼畸形类型,畸形率分别为12.22%、2.78%%、3.89%和1.11%。4种颅骨骨骼畸形均未表现出显著可见的外部形态变化。

图1 军曹鱼稚鱼颅骨骨骼畸形Fig.1 Cranium deformities in juvenile R. canadum
军曹鱼稚鱼脊柱畸形个体TL=(46.67±6.34) mm,畸形率为10.56%,共发现 6种骨骼畸形种类,分别为脊柱前凸(LO, 图 2A)、椎骨畸形(VD, 图 2B)、神经棘分叉(BNS, 图 2C)、脉棘分叉(BHS, 图 2D)、脉棘融合(HSF, 图 2E)、软骨冗余(CR, 图 2E、图 2F)。脉棘分叉和神经棘分叉的发生率最高,分别为4.44%和3.33%。在脊柱周围与脉棘之间发现有冗余的软骨存在。6种骨骼畸形均未表现出显著可见的外部形态变化。

图2 军曹鱼稚鱼脊柱骨骼畸形Fig.2 Vertebral column deformities in juvenile R. canadum
军曹鱼稚鱼尾鳍骨骼畸形个体TL=(43.36±8.27) mm,畸形率为15.56%,共发现尾上骨缺失(ED, 图3A)、尾上骨融合(EF, 图3B)、尾上骨畸形(EDM, 图3C)、尾下骨缺失(HYD, 图3D)、尾下骨与侧尾下骨愈合(HAHF, 图 3E)、尾下骨畸形(HYDE, 图 3F)等 6种骨骼畸形种类。尾上骨缺失和尾上骨融合的发生率最高,分别为 7.78%和3.89%。6种骨骼畸形均未表现出显著可见的外部形态变化。

图3 军曹鱼稚鱼尾鳍骨骼畸形Fig.3 Caudal fin deformities in juvenile R. canadum
军曹鱼稚鱼背鳍和臀鳍骨骼畸形个体 TL=(51.88± 7.12) mm,畸形率为5.56%,共发现支鳍骨分叉(BPT,图4A)、支鳍骨畸形(PTD, 图4A)、支鳍骨冗余(PTR,图4B)、支鳍骨异位(PTA, 图4C)、臀鳍条冗余(AFR,图4D)、背鳍棘缺失(DSD, 图2A)等6种骨骼畸形种类,骨骼畸形主要与支鳍骨相关。6种骨骼畸形均未表现出显著可见的外部形态变化。
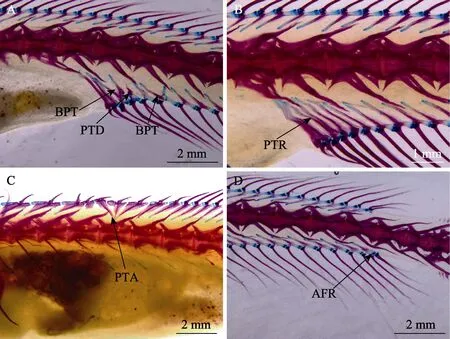
图4 军曹鱼稚鱼背鳍和臀鳍的骨骼畸形Fig.4 Dorsal and anal fin deformities in juvenile R. canadum
骨骼正常组[TL=(50.03±10.06) mm]与骨骼畸形组[TL=(44.56±7.79) mm]在全长上存在极显著差异(P<0.01);骨骼正常组分别与颅骨畸形组[TL=(43.14±7.13) mm] (P=0.003<0.01)及尾鳍畸形组[TL=(43.36±8.27) mm] (P=0.001<0.01)在全长上存在极显著差异,背、臀鳍畸形组[TL=(44.39±1.98) mm]分别与颅骨畸形组(P=0.033<0.05)和尾鳍畸形组(P=0.021<0.05)在全长上存在显著差异(图5)。
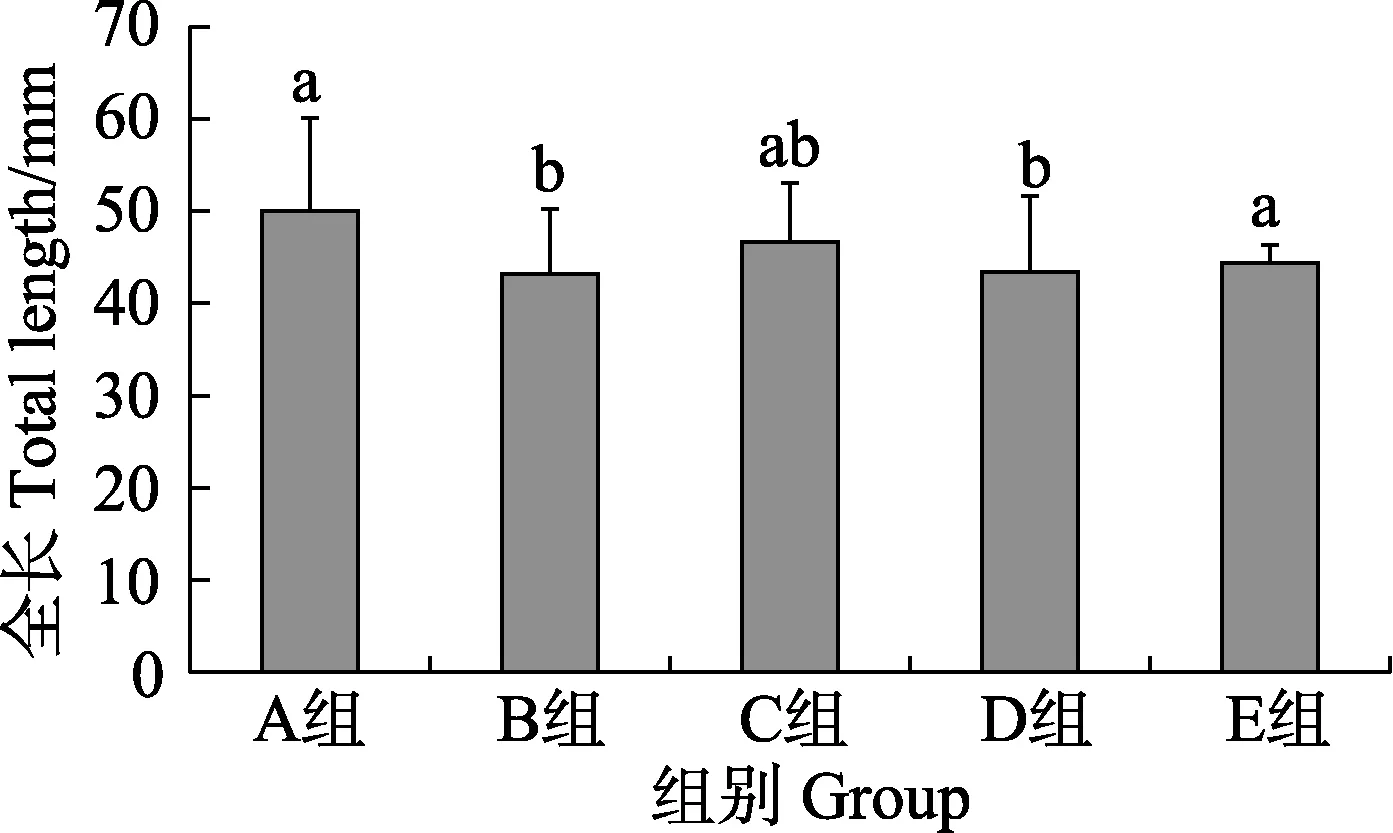
图5 骨骼正常组与各骨骼畸形组军曹鱼全长比较Fig.5 Total length of normal and skeletal deformed groups juvenile R. canadum
3 讨论
本研究首次报道了军曹鱼骨骼发育异常的形态特征。在 25 dph军曹鱼稚鱼的所有畸形种类中,颅骨畸形发生率最高,主要表现为米克尔氏软骨畸形、基舌骨畸形、基舌骨异位、舌弓下沉等,与郑攀龙(2015)研究发现的卵形鲳鲹(Trachinotus ovatus)仔、稚鱼的颅骨畸形类型一致,推测是由于二者发育阶段相似。米克尔氏软骨畸形是一种较为常见的鱼类骨骼畸形类型,黄尾(Seriola lalandi) (Cobcroft et al, 2004)、尖吻鲈(Lates calcarifer) (Fraser et al, 2005)、大西洋鲑(Salmo salar) (Witten et al, 2016)的仔稚鱼中均发现米克尔氏软骨畸形的存在。卵形鲳鲹中度和严重的米克尔氏软骨畸形可导致下颌变形(郑攀龙, 2015)。本研究中,军曹鱼米克尔氏软骨畸形属轻度畸形,鱼体未表现出明显的外部形态变化。此外,军曹鱼稚鱼中也未发现养殖2龄赤点石斑鱼(Epinephelus akaara)中所报道的鳃盖畸形或缺失现象(孔祥迪等, 2016),可能是由于发育日龄差异过大导致。军曹鱼颅骨畸形部位主要集中在与摄食相关的颌弓、舌弓,推测其原因可能与其摄食凶猛,摄食过程存在抢食现象有关(罗杰等, 2007)。此外,颅骨畸形个体全长指标极显著低于正常个体,可能这些畸形类型对摄食造成影响,并进一步对生长发育造成影响。
本研究中,军曹鱼尾鳍畸形率仅次于颅骨畸形率,尾鳍畸形个体全长指标极显著低于正常个体。种苗密度大会导致金头鲷(Sparusaurata)仔稚鱼尾鳍发生畸形(Koumoundouros et al, 1997)。本研究中,尾鳍畸形可能是由于饲养密度大,对鱼类生长速率产生影响,从而影响全长。此外,尾鳍骨骼畸形类型在不同鱼类间存在一定差异,军曹鱼的尾鳍畸形主要是尾上骨缺失和尾上骨融合;青石斑鱼(Epinephelus awoara)的尾鳍畸形主要是尾上骨的缺失(王秋荣等, 2012);大菱鲆(Scophthalmus maximus)中尾鳍畸形主要是鳍条畸形和尾杆骨畸形(吕雪娇, 2018);卵形鲳鲹主要是尾上骨和尾下骨的畸形(郑攀龙, 2015)。
军曹鱼神经棘及脉棘畸形发生的频率较高,这与大鳞副泥鳅(Paramisgurnus dabryanus) (张云龙, 2016)早期骨骼发育特征的结果相似,可能是由于二者体型相似。此外,军曹鱼软骨冗余主要发生在脉棘之间,但青石斑鱼中的软骨冗余发生部位为支鳍骨之间(王秋荣等, 2012),红鳍笛鲷(Lutjanus erythropterus)发育过程中相邻神经棘之间也存在增生软骨(程大川,2016),表明软骨冗余发生位置可能存在物种间差异。据报道,七带石斑鱼(Epinephelus septemfasciatus)(Nagano et al, 2007)、黄颡鱼(Pelteobagrus fulvidraco)(Huang et al, 2016)等在仔、稚鱼阶段即存在着较高的椎骨畸形率。卵形鲳鲹在11 dph即可观察到严重的椎骨畸形(郑攀龙等, 2014)。但在养殖军曹鱼稚鱼中仅发现极个别椎骨畸形个体。推测其原因,可能是由于脊柱畸形发生在发育早期,且这一畸形类型严重影响仔稚鱼的生存,因此,在存活的稚幼鱼个体中较少观察到这一畸形类型。
军曹鱼背鳍和臀鳍的畸形主要有鳍条冗余、支鳍骨分叉、支鳍骨畸形、支鳍骨弯曲、支鳍骨冗余和臀鳍条缺失等,类似的骨骼畸形在青石斑鱼和大菱鲆中均有报道(王秋荣等, 2012; 吕雪娇, 2018),但如大菱鲆和塞内加尔鳎(Soleasenegalensis)中鳍条的缺失、融合和分叉等畸形(吕雪娇, 2018; Gavaia et al, 2002)在军曹鱼中尚未发现。本研究中,军曹鱼样品背鳍和臀鳍的畸形主要集中在支鳍骨部位,推测其原因,可能是这一阶段稚鱼鳍条已经骨化完全,但支鳍骨仍处于骨化过程中,因此,鳍部骨骼畸形的发生可能与支鳍骨的骨化进程有关。
本研究结果显示,军曹鱼稚鱼骨骼畸形率与36 dph红鳍笛鲷的骨骼畸形率(39.50%)(程大川, 2016)相近,略高于相同发育阶段的卵形鲳鲹(22.00%~44.00%)(郑攀龙, 2015),但显著低于鞍带石斑鱼(Epinephelus lanceolatus)仔稚鱼(96.00%)(吕雪娇, 2018)、金头鲷稚鱼(43.20%~100%)(Boglione et al, 2001; Prestinicola et al,2013)、绯海鲷(Pagellus erythrinus)稚鱼(66.70%~80.00%)(Boglione et al, 2003)及舌齿鲈(Dicentrarchus labrax)稚鱼(>90.00%)(Barahona-Fernandes, 1982)的相应数值。本批次繁育的稚鱼畸形率低于其他鲈形目海水鱼,鱼体外表未见显著改变,可能是由于其培育环境优良。
常规养殖条件下,军曹鱼稚鱼骨骼畸形率最高部位为颅骨和尾鳍,这些骨骼畸形会直接影响鱼体摄食和游泳,进而影响其生长乃至存活(Koumoundouros et al, 1997)。加之骨骼畸形个体全长极显著低于正常个体,表明早期生长发育已受到骨骼畸形影响,由此可见,降低骨骼畸形率对军曹鱼养殖产业的意义重大。本研究通过鉴别常规养殖条件下军曹鱼稚鱼骨骼畸形的类型及其发生部位,为研究军曹鱼骨骼畸形的发生过程和原因、减少畸形率和优化苗种培育养殖条件提供理论基础。
