大坎波斯盆地盐下湖相介形虫特征及其古环境指示意义*
2022-01-07贾建忠康洪全蔡文杰贾怀存刘美羽
贾建忠 康洪全 蔡文杰 贾怀存 李 丹 刘美羽
(1. 中国海洋石油国际有限公司 北京 100028; 2. 中海油研究总院有限责任公司 北京 100028)
大坎波斯(Great Campos)盆地由桑托斯(Santos)、坎波斯(Campos)和圣埃思皮里图(Espirito Santos) 3个盆地组成。该盆地盐下湖相碳酸盐岩储层分布广,厚度大,油气资源潜力十分巨大。2007年以来在该盆地陆续发现了一批超大型油气田,如Lula油田、Franco油田和Libra油田等[1-3],均以盐下湖相碳酸盐岩为主要的储层,使得巴西成为仅次于委内瑞拉的南美洲第二大油气资源国。但是,近几年该盆地油气勘探进入瓶颈期。由于碳酸盐岩储层具有较强的非均质性,加上盐下地震成像相对较差,优质储层的分布预测一直是勘探面临的难题,而发育环境和控制因素是储层预测的关键。该套碳酸盐岩储层的成因目前尚无定论,争议的焦点在于湖泊环境能否形成如此广泛而稳定的碳酸盐沉积,以及控制其发育和消亡的关键环境因素[4-6]。
大坎波斯盆地盐下裂谷期的主要储层类型为生物碎屑灰岩和微生物灰岩。生物碎屑灰岩主要包含介壳灰岩和颗粒灰岩两种类型,生物碎屑以双壳类为主,介形类和腹足类为辅,并含有其他碳酸盐岩组分和部分硅质碎屑[7];微生物灰岩主要为藻类和叠层石相互粘结而形成的礁体或席状沉积[6],含少量介形类等微体化石。介形虫是具有双半壳的节肢动物,主要营底栖或游泳生活,对湖相生态环境,包括湖水的盐度、深度、水化学条件等反应敏感。本文通过部分盐下探井的介形虫生物地层及古生态学分析,重建了研究区裂谷期古湖泊水深和盐度等环境条件,进而对盐下湖相碳酸盐岩储层发育的控制因素进行了初步探讨。
1 区域地质背景
大坎波斯(Great Campos)盆地位于巴西东南部被动大陆边缘,由桑托斯(Santos)、坎波斯(Campos)和圣埃思皮里图(Espirito Santos) 3个盆地组成,三者中间被卡布弗里奥(Cabo Frio)高地和维多利亚(Vitoria)高地所分隔,总面积约58.68×104km2(图 1)[3,8]。受晚三叠世以来大西洋持续裂解作用的控制,盆地形成了近东西向“隆坳相间”的构造格局,为碳酸盐岩的广泛发育提供了构造条件。
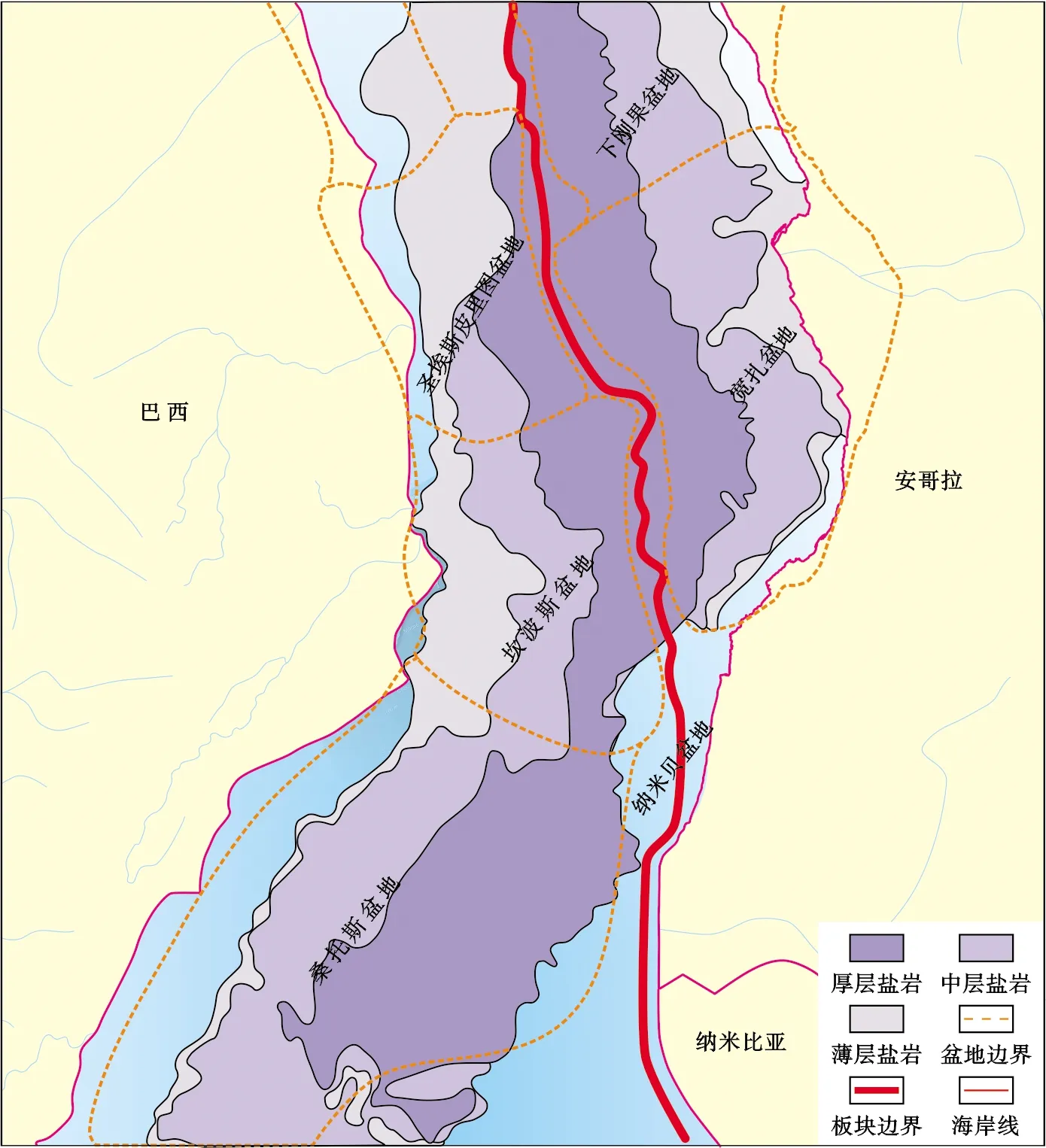
图1 大坎波斯盆地盐下裂谷期构造位置图Fig .1 Syn-rift tectonical location of the Great Campos basin
大坎波斯(Great Campos)盆地主要经历了裂谷期、过渡期和漂移期三大构造演化阶段。裂谷期又可划分为断陷早期、断陷晚期、反转期和拗陷期,分别对应区域超层序KS4—KS7(图 2)。断陷早期(KS4)为早白垩世Neocomian-Barremian期,该阶段地壳减薄,冈瓦纳大陆解体,非洲大陆和南美大陆间发生陆内裂谷作用,形成平行海岸线的地堑和地垒,发育了一系列NW—SE向克拉通裂谷盆地;裂陷早期下部以火山岩充填为主,晚期广泛沉积了富含有机质的深水湖相泥页岩,是研究区重要的烃源岩。早白垩世Barremian晚期进入断陷晚期(KS5),期间构造稳定,早期发生显著湖侵,湖盆面积迅速扩张;中期开始中央隆起带缓慢抬升,湖平面稳定下降,在隆起带及周边沉积了大规模湖相贝壳灰岩和生物碎屑灰岩;末期湖平面快速上升,碳酸盐台地大多被淹没,沉积一套薄层泥质岩。早Aptian期的反转期也可称为断拗转换期(KS6),以区域性的构造抬升和破裂不整合的发育为主要特征。从中晚Aptian期开始,盆地进入拗陷期(KS7),由断陷向热沉降过渡,形成了宽缓的湖盆,海水开始周期性进入盆地,在近岸地带发育浅湖相和三角洲相沉积,在中央隆起带及其周边广泛分布微生物席及藻叠层石建造。Aptian末进入过渡期,受Walvis Ridge打开的影响,南侧的海水大规模涌入盆地,沉积了Loeme组蒸发盐岩,盆地中心厚度可达3 000 m,几乎将裂谷期地层全部覆盖[2,8]。

图2 大坎波斯盆地盐下综合地层格架Fig .2 Syn-rift stratigraphic framework of the Great Campos Basin
2 介形虫生物地层划分及标志
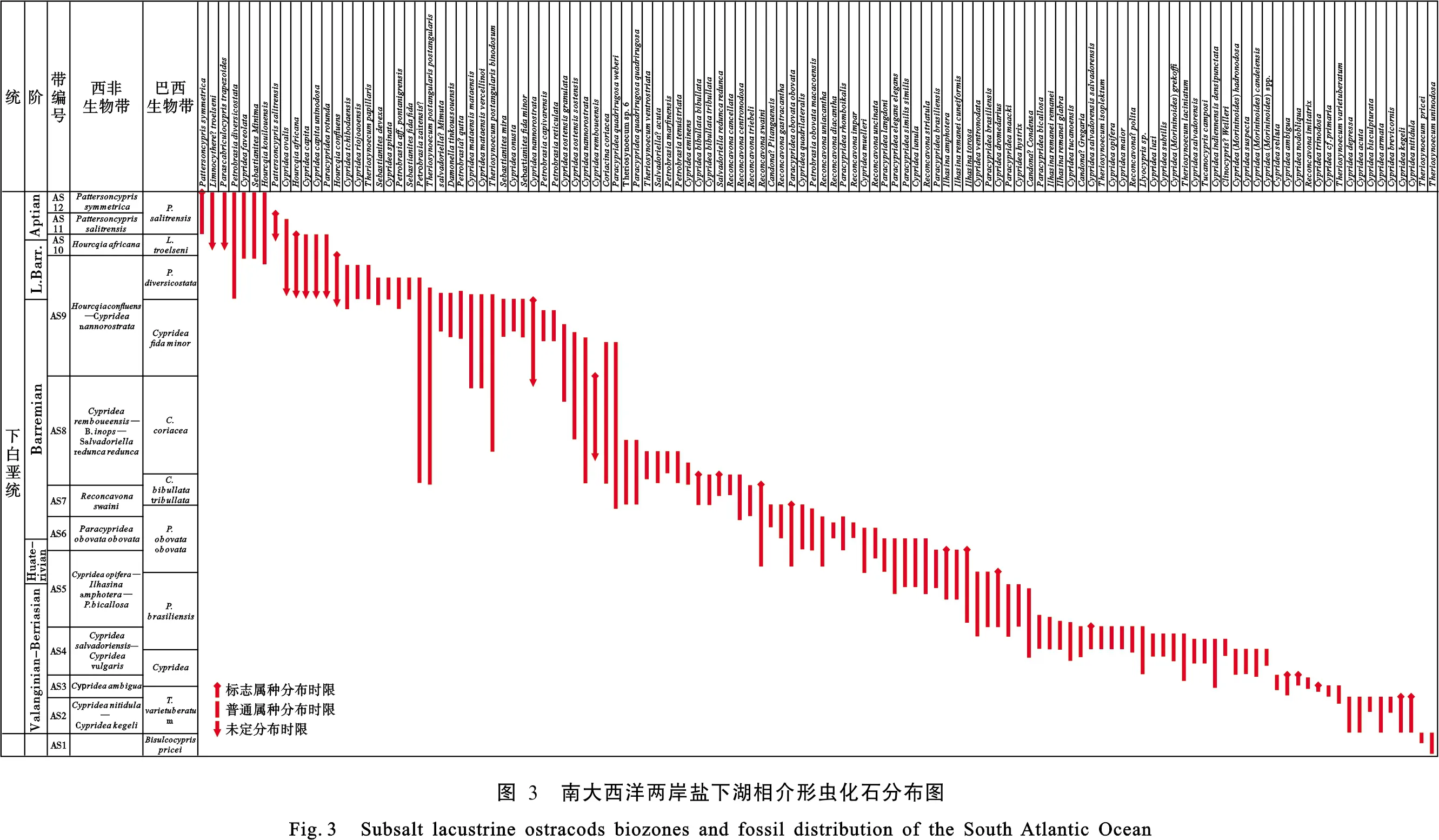
南大西洋两岸盆地盐下裂谷期介形虫分布广泛且种类丰富,是湖相地层划分和对比的重要依据。20世纪90年代,伴随着石油勘探热潮的兴期,道达尔等石油公司在巴西和西非海岸开展了较为系统的介形虫生物地层研究。Grosdidier等(1996)[9]建立了西非盐下裂谷盆地中的非海相介形虫化石带,并一直沿用至在;Bate(1999)[10]进一步对上述盆地介形虫组合及其在层序地层研究中的作用进行了系统论述,认为介形虫组合的演化特征与沉积演化旋回之间存在良好对应关系。Poropat和Colin(2012)[11]对巴西和西非海岸的早白垩世介形虫生物地层进行了综述性研究,为开展地层对比和古生态、古环境研究奠定了良好基础。本文在参考上述有关介形虫生物地层方面研究认识基础上,对研究区内已有文献资料和若干钻井的古生物报告资料进行了总结和对比,明确了盐下介形虫标准化石120种,其中西非特有14种,巴西特有41种,两岸共有种65种,建立了南大西洋两岸统一的盐下介形虫标准化石带(图 3),划分出介形虫带AS1—AS12,共计12个。
由下到上,各介形虫带的主要标志化石及其组合特征如下:
AS1带(Tithonian期):位于上侏罗统顶部,以Bisulcocyprispricei为带化石,见少量浅水标志种Darwinulacf.D.oblonga等。
AS2带(Barriasian期):位于下白垩统底部,带化石为Cyprideakegeli,并以Cyprideanitidula出现为标志。
AS3带(早Valangian期):以Cyprideaambigua为标志,见Cyprideasellata、Cyprideatrinodosa等。
AS4带(晚Valanginan期):以Cyprideasalvadoriensis、Cyprideavulgaris的出现为标志,同时产出Cyprideahadronodosa、Cyprideasellata、Cyprideagrekoffi、Cyprideasubtillis、Tucanocypris、Tucanocyprisapiculata等。
AS5带(Hauterivian期):以Cyprideaopifera、Cyprideadromedarius、Ilhasinaamphotera、Ilhasinatorosa等种出现为标志,亦可见Paracypridea属的Paracyprideamaacki、P.impara、P.bicallosa等。
AS6带(早Barremian期):以Paracyprideaobovataobovata、Paracyprideaspp.的出现及Reconcavona的增多为标志,出现Paracypridea属的P.langdoni、P.elegans、P.similisMetacyprisspp.等种。
AS7带(早—中Barremian期):以Cypridearemboueensis为标志,见Cyprideabapounou、Cypridealukungaensis、Reconcavonatriebeli等种。部分地区缺失。
AS8带(中—晚Barremian期):顶面为Cypridearemboueensis末现面,见Salvadoriella?dissimilis、Paracyprideaquadrirugosaweberi、Cyprideafalcaoensis、Cyprideabibullatatribullata、Reconcavonacentronodosa、Reconcavonacancellata等种。
AS9带(晚Barremian期):顶界面以Hourcqiaconfluens的末现为标志,伴有Cyprideatchibodaensis、Cyprideaminima、Cyprideafida、Cyprideacapita、Cyprideamucuaensis、Cyprideaovalis、Hourcqiaafricana、Petrobrasiatenuistriata、Petrobrasiacompacta、Cyprideasostensis等,西非地区可见Cyprideahollensis、Cyprideasylvesterbradleyi等。
AS10带(晚Barremian—早Aptian期):以Hourcqiaafricana末现面为顶,部分地区现Cyprideacapita、Cyprideacapitauninodosa、Paracypridearotunda、Damonella?tinkoussouensis、Reconcavona?ultima.、Cypridealoango等。
AS11带(中Aptian期):以化石Pattersoncyprissalitrensis为标准带化石,可见Pattersoncyprissymmetrica、少量Pattersoncypriselongata等同属相关种,部分地区现Pattersoncypriselongata、Parascabriculocypristrapezoidesnodosa、Limnocytherebituberculata、Cyprideatribulbosa、Cyprideaposteronodosa、Parascabriculocyprisnodosa、Theriosynoecumkwanzaensis、Cyprissp.A、Strenestroemiasp.、Cyprideasetinaangolensis、Cyprideaaptiansa、Cyprideaquadribulbosa、Paraklieanaalata、Cypridealoango、Cyprideacapita等。
AS12带(晚Aptian):顶面与Pattersoncyprissymmetrica末现重合,带内现丰富的平滑壳介形虫,静水种十分常见,同时可见Limnocythere?troelseni、Parascabriculocypristrapezoides及少量Discocyprisplana。
3 介形虫古生态环境分析
在湖泊体系中介形虫对盐度、温度和水深等环境因素的变化敏感。因此,介形虫属种组合特征可作为还原古盐度、古温度、古水深等古环境参数的重要参考[12-14]。
随着盐度升高,仅能在淡水和低盐度水体中生存的淡水种—微咸水种会快速消失,取而代之以半咸水种和咸水种[15]。同时,介形虫的一些生态指标和特定种属的出现可以反映古水深变化。李守军 等[16](2005)利用介形虫优势分异度实现了对东营凹陷古近系沙三段古湖泊水深的半定量恢复。李顺 等[17](2011)利用松辽盆地松科-1井介形虫生物群的简单分异度和复合分异度对盆地晚白垩世湖平面变化进行了分析。
通过研究区钻井介形虫古环境分析,结合国内外相关对比研究[14-17],明确了大坎波斯盆地盐下裂谷期湖相介形虫的盐度和古水深标志种(表1、2)。盐度标志种分为淡水种、微咸水种、半咸水种和咸水种4类;古水深标志种分为浅水种、半深水—深水种和海陆过渡种3类。

表1 大坎波斯盆地裂谷期介形虫古盐度标志种Table 1 Ostracods marker species of syn-rift paleo-salinity in the Great Campos basin

表2 大坎波斯盆地裂谷期介形虫古水深标志种Table 2 Ostracods marker species of syn-rift paleo-water depth in the Great Campos basin
研究区南部的桑托斯盆地盐下湖相介形虫化石丰富,满足古生态学精细研究的需要。通过对介形虫盐度标志种的统计,发现层序KS5(Itapema组)贝壳灰岩沉积期介形虫以淡水种—微咸水种占绝对优势;而层序KS7(Barra Vella组)微生物灰岩沉积期以半咸水种占明显优势(图4),指示贝壳灰岩沉积期为微咸水环境,微生物灰岩沉积期为半咸水环境。

图4 桑托斯盆地S-1井盐下湖相介形虫古盐度标志种分布图Fig .4 Ostracods marker species distribution of syn-rift lacustrine paleo-salinity in Well S-1 in the Santos basin
根据介形分异度及水深标志种,层序KS5—KS7纵向上可识别出5个湖平面变化旋回(图5),但总体为浅湖环境,且呈现水退趋势,只有在贝壳灰岩发育初期和末期发生两次较大规模的湖泛事件。另外,分析结果显示贝壳灰岩沉积期湖水明显较微生物灰岩沉积期深,这与之前的认识有所不同。在贝壳灰岩沉积早期和末期,介形虫分异度明显升高,且出现了代表水体相对较深的Hourcqiaafricanaafricana、Hourcqiaafricanaconfluens、Cyprideariojoanensis、Cyprideasebastianites等属种;在微生物灰岩沉积期,介形虫分异度较低,只有个别属种存活,如Pattersoncyprisspp.、Limnocyprideatroelseni等,代表了典型浅水环境。通过桑托斯盆地连井对比(图6),可以明显看出介形虫分异度逐渐下降,浅水种自下而上逐渐增多,反映了湖平面逐渐下降的趋势,尤其是微生物灰岩沉积之前,整个区域遭受抬升剥蚀,发育大型不整合面,导致整个AS11介形虫带缺失。

图5 桑托斯盆地S-1井盐下湖相介形虫属种分布及古盐度、古水深变化图Fig .5 Ostracods marker species distribution of syn-rift lacustrine paleo-salinity and paleo-water depth in well S-1 in the Santos basin
此外,本文首次在S-1和S-2井的多个层位发现了若干海陆过渡相介形虫属种(图4、5),包括Cytheridea? sp.308,Cytherideasp.,Pattersoncyprissymmetrica,Pattersoncyprismicropapillosa等,尤其Pattersoncyprismicropapillosa多生活在与海相连通的高盐度湖泊,或局限海湾环境中[18]。据此推测在KS5贝壳灰岩沉积早期、晚期,以及KS7微生物灰岩沉积中期发生了至少三期规模不等的海侵事件(图6)。

图6 桑托斯盆地盐下湖相介形虫与古水深、海侵事件关系图Fig .6 Ostracods palaeoecological index of syn-rift lacustrine water paleo-depth and marine incursion in the Santos basin
4 大坎波斯盆地盐下湖相碳酸盐岩形成环境分析
大坎波斯盆地盐下不同时期、不同类型湖相碳酸盐岩的发育,主要受控于构造抬升、古气候与水化学条件、海侵事件等关键因素,是各因素共同作用的结果。首先,隆坳相间的构造背景为湖相碳酸盐岩发育提供了有利的地貌条件。西部为近平行岸线NE走向的隆起带,中、东部为隆起欠发育的低隆区,中部坳陷带与东部坳陷带向北部合拢,内部发育多个低凸起区。构造隆起带有效阻隔了陆源碎屑的大量输入,形成了清澈而温暖的水体环境,有利于造礁生物的繁盛和化学沉积的发育。另一方面,研究区持续的抬升,在中央隆起带形成了广阔、清洁、透光性良好的浅水环境,有利于介形虫、双壳类、叠层石等造礁生物的繁盛,形成了大范围的礁滩相碳酸盐沉积[19-20]。
另外,古气候对盆地沉积充填同样具有重要控制作用。白垩纪中期正式进入温室气候期,大坎波斯盆地位于低纬度区,在Albian—Cenomanian期为最热时期[21-26],整体属典型干热气候,随降雨和淡水河流输入的减少及蒸发量的增加,导致湖泊盐度逐渐升高,形成了咸化的碱性湖水环境。温暖的气候和高矿化度的水体,非常有利于甲壳类生物的繁盛和碳酸盐岩的大量沉积。但是,当湖水盐度或pH值过高时,可能导致双壳类等大型底栖生物无法生存,取而代之的是菌藻类微生物,从而使岩相由早期的贝壳灰岩转变为晚期的藻叠层石灰岩。这主要表现在研究区中央隆起带,早期广泛沉积KS5贝壳灰岩和,转变为晚期广泛沉积了KS7席状微生物岩和藻叠层石灰岩。在KS7微生物灰中发现的介形虫Limnocyprideasubquadrata和Limnocyprideatroelseni等,也指示了偏碱性的水介质条件环境。
海侵事件则为大坎波斯盆地盐下湖相造礁生物的繁盛和碳酸盐的富集创造了有利的水介质条件。前已述及,桑托斯盆地在贝壳灰岩沉积期和微生物灰岩沉积期均发生数期海侵事件。在区域上也存在较多与之呼应的海侵证据。巴西东北部的阿拉里皮(Araripe)盆地发育早白垩世Aptian期Pattersoncyprissp.-Reconcavonasp.-Cyprideaararipensis半咸水介形虫组合,指示了与海连通的湖泊环境[18,21]。普提瓜尔(Potiguar)盆地Aptian期湖相介形虫和海相浮游生物(沟鞭藻、有孔虫等)共生,也指示了海侵作用的存在。另外,西非一侧的加蓬和下刚果盆地也在该时期分别发现了海相鱼化石、海相介形虫等,说明南大西洋中段的含盐盆地至少在Aptian期就已经有海水从北部灌入[20]。同时,南部坎波斯盆地出现了浮游有孔虫和海相低硫有机质、宽扎盆地发育的盐岩沉积以及在桑托斯盆地产出的海陆过渡相介形虫,足以说明在南大西洋中段含盐盆地南部也存在海侵通道[21]。在晚Barremian—早Aptian,受构造活动和海平面上升等影响,南大西洋南部海水数次越过Wives脊的阻挡,灌入裂谷期湖泊,形成了大范围的半咸水环境,为造礁生物繁盛和碳酸盐富集创造了有利的咸化水介质条件[22]。
5 结论
1) 建立了南大西洋两岸盐下裂谷期湖相介形虫化石带,划分出12个介形虫带(AS1—AS12),明确了120个具有年代意义的介形虫标志化石,其中西非特有14种,巴西特有41种,两岸共有种65种。
2) 明确了大坎波斯盆地盐下裂谷期湖相介形虫的盐度和古水深标志种,指出碳酸盐沉积期总体为浅湖环境,且由下到上呈现湖平面下降的趋势;同时,Itapema组贝壳灰岩沉积期为微咸水环境,Barra Vella组微生物灰岩沉积期为半咸水环境,并发现了Cytherideasp.,Pattersoncyprissymmetrica,Pattersoncyprismicropapillosa等半咸水海侵标志属种,指示了至少3期海侵事件。
3) 大坎波斯盆地大规模湖相贝壳灰岩和微生物灰岩的发育得益于构造抬升、温—热的气候条件和多期海侵事件等多种有利因素的良好匹配。
