福建天台山半枫荷天然群落的物种组成、生态位和种间关系
2021-12-28叶兴状王妙青张明珠范辉华张国防赖日文
叶兴状, 王妙青, 程 诺, 张明珠, 范辉华, 张国防, 赖日文, 刘 宝,①
(1. 福建农林大学林学院, 福建 福州 350002; 2. 福建省林业科学研究院特色资源研究所, 福建 福州 350012)
研究濒危植物所处群落的生态位特征和种间关系,是揭示濒危植物生存现状和群落演替的有效方法[1]。研究生态位和种间关系不仅有助于加强对研究对象濒危机制的认识,分析生境差异对其分布格局的影响,还可为其保护措施的制定提供科学依据[2]。许多濒危植物受自身生物学和生态学特性的制约,野外种群数量相对较少[3]。濒危植物数量减少或灭绝直接影响生物多样性的稳定,进而引发生态系统结构和功能的剧烈变化[4]。藤枣〔Eleutharrhenamacrocarpa(Diels) Forman〕[5]、紫纹兜兰〔Paphiopedilumpurpuratum(Lindl.) Stein〕[6]和长序榆(UlmuselongataL. K. Fu et C. S. Ding)[7]等国家重点保护植物的种间联结性研究,揭示了这些植物对环境的适应性及其生境地植物种群动态,为各物种多样性保护及濒危机制探索提供了理论依据。
半枫荷(SemiliquidambarcathayensisChang)为中国特有种,属于深根、喜光偏阳性树种,残存于中国南方山区[8]。半枫荷对生境条件要求苛刻,分布范围狭窄,种群规模小,大多数单株呈散生分布状态,在《中国高等植物受威胁物种名录》中被列为易危(VU)[9-10]。半枫荷具有较大的开发价值,首先,其材质优良、树干通直,是理想的家具和工业原材料;其次,半枫荷是珍贵的药用植物,其根、叶、花和果均可入药,且叶中有效成分种类多、含量丰富,可治疗风湿跌打、瘀积肿痛和产后风瘫等疾病[11];再次,半枫荷具有异形叶,初春嫩叶为紫红色,深秋则有绿、黄、紫红至暗红等颜色,是中国南方潜在的园林绿化树种[12]。然而,半枫荷的生境破坏严重,繁育更新困难,野生种群数量急剧下降,资源几近枯竭,生存现状堪忧。目前,国内学者对半枫荷的研究主要集中在育苗造林[12]、遗传多样性[10,13]、药用价值[14]和优树选择[15]等方面,有关其物种保护方面的研究较缺乏,其濒危机制的外因尚不明确,严重制约半枫荷种质资源的保护和利用。
鉴于此,本研究基于福建沙县的天台山半枫荷天然群落设置调查样地,通过分析半枫荷群落的物种组成、生态位宽度、生态位重叠、种间联结性和群落稳定性等,探讨半枫荷的生态适应能力及其与群落伴生种的竞争资源压力强度,以期为半枫荷濒危机制、就地保护及迁地保护研究提供科学的理论依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于福建省三明市沙县区高桥镇桂岩村天台山,具体地理位置为东经117°47′58.12″~117°51′38.31″、北纬26°39′04.02″~26°42′56.48″,海拔650~1 105 m,地处武夷山东侧支脉区域,毗邻三明罗卜岩自然保护区。该区域气候类型属于中亚热带季风气候,干湿季分明,3月至6月多阴雨,秋冬多晴,年平均气温19.2 ℃,最冷月(1月)平均气温7.5 ℃,极端低温-8.0 ℃,最热月(7月)平均气温28.7 ℃,极端高温40.1 ℃,年活动积温4 978.2 ℃,无霜期303 d,年降水量1 662.4 mm,年蒸发量817.3 mm,空气相对湿度71%。该区域土壤以黄壤和红黄壤为主,土壤松散,偏沙壤,pH 4.3至pH 5.5,土壤表面的凋落物和腐殖质层较厚,厚度达4~8 cm。半枫荷集中分布在天台山的中上坡。
1.2 研究方法
于2018年7月,在对天台山全面踏查的基础上,采用典型抽样法,以半枫荷生长的位置为中心设置样地。考虑到野生半枫荷数量稀少且呈零星分布,结合半枫荷的分布位置和地形,设置4个面积20 m×20 m的样地(表1),调查面积合计1 600 m2。调查并记录样地中所有半枫荷的胸径(DBH)、枝下高、株高、冠幅和经纬度等信息。将每个样地划分为4个面积10 m×10 m的样方,调查并记录每个样方中乔木层所有植株(DBH≥5 cm)的种名、胸径、株高、冠幅、枝下高和经纬度等信息。在每个样地四角及中心设置5个面积5 m×5 m的小样方,调查并记录灌木层所有植株的株高、胸径(地径)和盖度。胸径使用围径尺(精度1 mm)测量,株高、冠幅和枝下高使用哈光林业测高器CGQ-1(精度0.5 m)测量,盖度采用方格法估测。
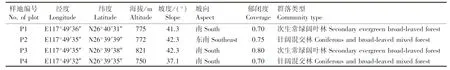
表1 福建天台山半枫荷调查样地的基本情况
1.3 数据处理
1.3.1 生态位和种间联结 分别对乔木层和灌木层的种类进行统计和整理,计算各种类的重要值、Levins生态位宽度[16]、Shannon-Weaver生态位宽度[17]和生态位重叠值[18]。

根据已建立的2×2联列表,用χ2统计量检测种间关联性。若χ2≥3.841,则种间呈显著性关联;若χ2<3.841,则种间呈不显著关联。然后,对关联不显著的种进一步利用结合点相关系数(PCC)、联结系数(AC)、关联度指数(OI)和共同出现百分率(PC)等综合分析,以测定联结程度即关联度是否紧密[20]。其中,PC≥0.6为紧密关联;PCC≥0.6或AC≥0.6表示紧密正关联;PCC≤-0.6或AC≤-0.6表示紧密负关联;PC=0.0或AC=0.0表示种间完全独立。
1.3.2 群落Godron稳定性指数 采用郑元润[21]改进的Godron稳定性指数法测定群落稳定性,对群落中乔木层所有种类按照相对频度由大到小排序,将乔木层种总数倒数累计百分比与相对频度累计百分比一一对应,采用散点图平滑曲线模拟一元二次方程与线性方程y=100-x的交点坐标,判断群落的稳定性,交点越接近群落的稳定点(20,80),群落越稳定。
1.3.3 数据处理和分析 采用EXCEL 2010和SPSS 23.0软件对数据进行处理和分析。因灌木层基本没有半枫荷幼苗和幼树,所以种间联结性和群落稳定性仅统计乔木层。
2 结果和分析
2.1 群落物种组成、重要值和生态位宽度分析
2.1.1 群落物种组成 调查结果(表2和表3)显示:半枫荷群落乔木层和灌木层植物共有27科39属70种645株,其中,乔木层有47种424株,隶属于23科36属,灌木层有52种221株,隶属于26科38属。在乔木层和灌木层中,株数超过10株的种类分别占乔木层和灌木层种数的19.1%和11.5%,其株数分别占乔木层和灌木层株数的71.0%和37.6%;乔木层和灌木层仅有1株的植物分别有9和16种,分别占乔木层和灌木层种数的19.1%和30.8%。

半枫荷群落中栲(CastanopsisfargesiiFranch.)的株数最多,有165株,占总株数的25.6%;半枫荷有28株,占总株数的4.3%;赤杨叶〔Alniphyllumfortunei(Hemsl.) Makino〕、虎皮楠〔Daphniphyllumoldhami(Hemsl.) Rosenthal〕、大叶苦柯(LithocarpuspaihengiiChun et Tsiang)、油茶(CamelliaoleiferaAbel.)、杨桐〔Adinandramillettii(Hook. et Arn.) Benth. et Hook. f. ex Hance〕和百两金〔Ardisiacrispa(Thunb.) A. DC.〕的株数也较多,均在15株及以上。
2.1.2 重要值特征 调查结果显示:乔木层中,栲的重要值最大,为20.16%;半枫荷的重要值次之,为7.19%,是乔木层优势种之一;重要值大于5%的种类还有锥栗〔Castaneahenryi(Skan) Rehd. et Wils.〕、赤杨叶、大叶苦柯和马尾松(PinusmassonianaLamb.)。灌木层中,栲的重要值也最大,为7.15%,而半枫荷的重要值仅为0.94%,位居第31,表明半枫荷幼树(幼苗)数量稀少,天然更新弱;重要值大于5%的种类还有杨桐和油茶。栲在4个样地的分布较均匀,数量较多;与栲相比,半枫荷的分布不均匀,优势不明显,数量仅为栲的17.0%。
2.1.3 生态位宽度特征 调查结果(表2和表3)显示:乔木层中,半枫荷的重要值位居第2,Levins生态位宽度和Shannon-Weaver生态位宽度位居第4,Levins生态位宽度和Shannon-Weaver生态位宽度分别为3.340和1.279,说明半枫荷在群落中并不是处在优势地位,适应能力较弱。而栲的重要值和生态位宽度均最大,Levins生态位宽度和Shannon-Weaver生态位宽度分别为3.980和1.384,生态适应能力较强,且栲在各样方分布较均匀,竞争能力较强。虽然锥栗、马尾松和木油桐(VerniciamontanaLour.)等种类的重要值排名靠前,但这些种类只分布在个别样方中,生态位宽度排名相对靠后。与之相反,山槐〔Albiziakalkora(Roxb.) Prain〕、黄牛奶树〔Symplocoscochinchinensisvar.laurina(Retzius) Nooteboom〕、钟花樱桃〔Prunuscampanulata(Maxim.) Yü et Li〕和野柿(Diospyroskakivar.silvestrisMakino)等种类的重要值排名相对靠后,但这些种类分布的样方和频度均较多,生态位宽度排名相对靠前。毛锥(CastanopsisfordiiHance)、南酸枣〔Choerospondiasaxillaris(Roxb.) B. L. Burtt et A. W. Hill〕和木荷(SchimasuperbaGardn. et Champ.)虽然分布的样方较少,频度较小,但胸高断面积较大,三者的生态位宽度排名也相对靠前。
灌木层,半枫荷仅在P1样地的1个样方中分布2株,其重要值和生态位宽度排名均靠后。与之相反,虽然百两金重要值排名第7,但其生态位宽度最大,Levins生态位宽度和Shannon-Weaver生态位宽度分别为3.383和1.305。此外,短尾越橘(VacciniumcarlesiiDunn)、杨桐、栲、南烛(VacciniumbracteatumThunb.)、朱砂根(ArdisiacrenataSims)、虎皮楠和油茶的生态位宽度也较大,在各样方基本都有分布。而米槠〔Castanopsiscarlesii(Hemsl.) Hayata〕、浙江润楠(MachiluschekiangensisS. Lee)、马银花(Rhododendronovatum(Lindl.) Planch.)、米碎花(EuryachinensisR. Br.)、杨梅(MyricarubraSieb. et Zucc.)和青冈〔Cyclobalanopsisglauca(Thunb.) Oerst.〕等种类的生态位宽度较小,仅在1个样地有分布,与半枫荷相似。
2.2 群落种类生态位重叠和种间联结
2.2.1 生态位重叠特征 半枫荷天然群落乔木层和灌木层主要种类(重要值排名前31位)的生态位重叠值(NOV)见图1。由图1可见:乔木层主要种类生态位重叠值的平均值为0.571,其中,半枫荷与赤杨叶、米槠和红楠(MachilusthunbergiiSieb. et Zucc.)等7个种类的生态位重叠值大于0.9,与乔木层其他主要种类的生态位重叠值的平均值为0.720;0.9≤NOV≤1.0的种对有43对,占总种对数的9.2%,其中,生态位重叠值为1.0的种对有4对,分别是油茶和杉木〔Cunninghamialanceolata(Lamb.) Hook.〕、芬芳安息香(StyraxodoratissimusChamp. ex Benth.)和树参〔Dendropanaxdentiger(Harms) Merr.〕、山胡椒〔Linderaglauca(Sieb. et Zucc.) Bl.〕和山乌桕(TriadicacochinchinensisLour.)以及福建山矾(SymplocosfukienensisLing)和野漆〔Toxicodendronsuccedaneum(Linn.) O. Kuntze〕;生态位重叠值小于0.2的种对有62对,占总种对数的13.3%,其中生态位重叠值大于0.5的种对有312对,占总种对数的67.1%,表明多数种对的生态位重叠值较高。
灌木层主要种类生态位重叠值的平均值为0.500,其中,0.9≤NOV≤1.0的种对有97对,占总种对数的20.9%,生态位重叠值为1.0的种对有4对,分别为小叶石楠〔Photiniaparvifolia(Pritz.) Schneid.〕和苦槠〔Castanopsissclerophylla(Lindl. et Paxton) Schottky〕、米碎花和杨梅、半枫荷和米碎花以及半枫荷和杨梅;生态位重叠值大于0.5的种对有231对,占总种对数的49.7%;而生态位重叠值小于0.1的种对有91对,占总种对数的19.6%,生态位重叠值为0.0的种对有66对,占总种对数的14.2%。

: 0.9≤NOV≤1.0; : 0.8≤NOV<0.9; : 0.6≤NOV<0.8; : 0.4≤NOV<0.6; : 0.2≤NOV<0.4; : 0.0≤NOV<0.2.
从表4可知:半枫荷与乔木层主要种类的χ2小于3.841(P>0.05),说明其与乔木层主要种类联结不显著。从联结系数(AC)和结合点相关系数(PCC)看,与半枫荷正联结的种类有20种,占乔木层主要种类数(除去半枫荷)的66.67%,负联结的有9种,占乔木层主要种类数的30.00%。|AC|>0.6的仅有5种,其中,木油桐、毛锥、木荷和山胡椒为紧密正关联,甜槠〔Castanopsiseyrei(Champ. ex Benth.) Tutch.〕为紧密负关联。而|AC|<0.4的有17种,占乔木层主要物种数的56.67%;除了甜槠和木油桐,其余种类的|PCC|都小于0.5;除了栲、赤杨叶和木油桐外,其余种类的共同出现百分率(PC)均小于0.5,说明半枫荷与大部分乔木的关联性不强或倾向于正联结。
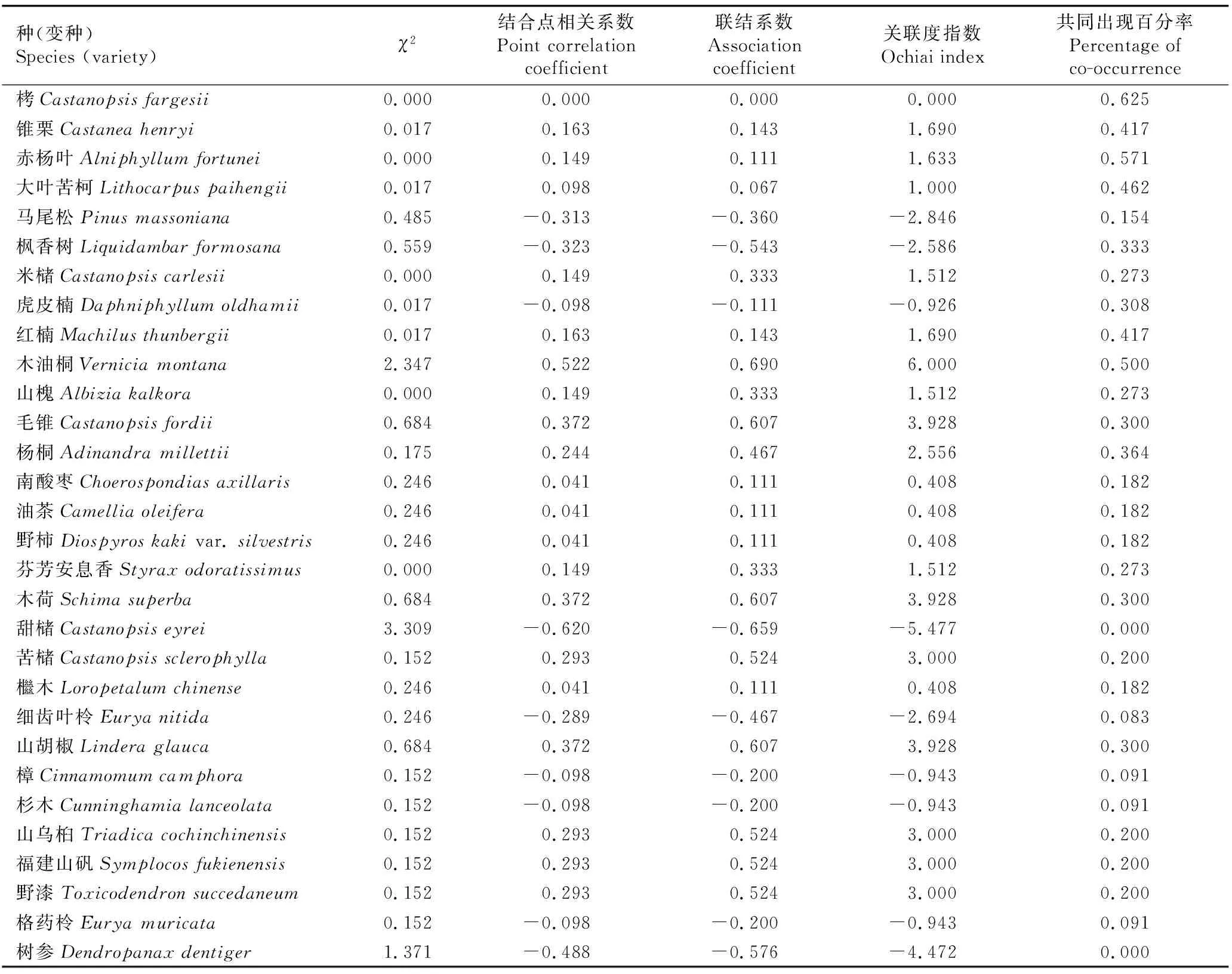
表4 福建天台山半枫荷天然群落中半枫荷与乔木层主要种类(重要值排名前31位)的联结性指标
2.3 群落稳定性分析
结果(图2)显示:半枫荷群落乔木层种类Godron稳定性指数曲线模拟方程y=-0.009 9x2+1.821 9x+13.044 0(R2= 0.990 6)与线性方程y=100-x的交点坐标为(35.45,65.19),离群落稳定点(20,80)相对较远。
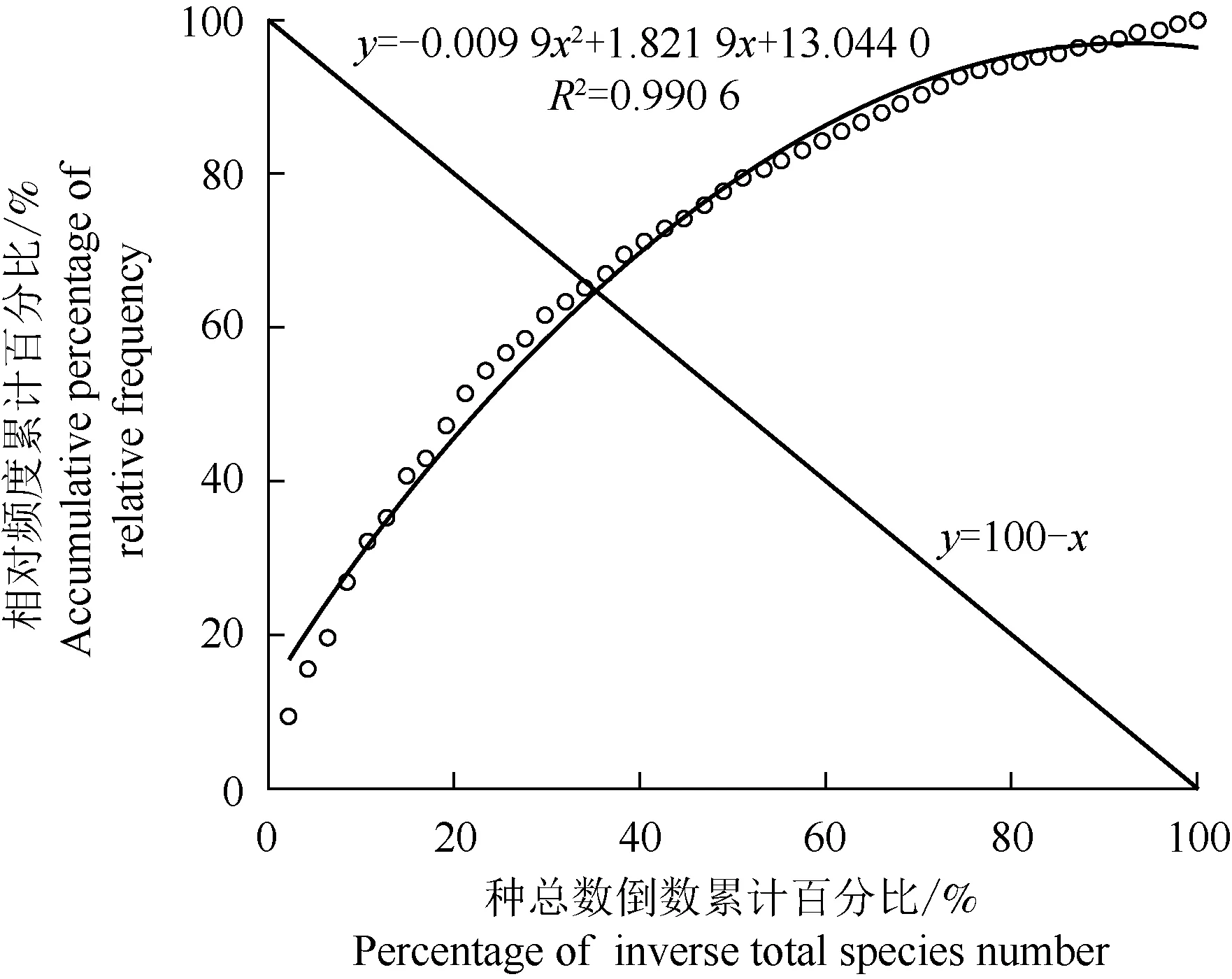
图2 福建天台山半枫荷天然群落乔木层种类的Godron稳定性
3 讨 论
本研究中,福建沙县天台山半枫荷群落乔木层和灌木层植物有27科39属70种645株,而半枫荷数量仅占总株数的4.3%。半枫荷群落类型为次生常绿阔叶林和针阔混交林,林分层次结构明显。其中,乔木层优势种有栲、半枫荷、锥栗、赤杨叶、大叶苦柯、马尾松和枫香树等,灌木层优势种有栲、杨桐、油茶、虎皮楠、草珊瑚〔Sarcandraglabra(Thunb.) Nakai〕、薄叶山矾(SymplocosanomalaBrand)和百两金等。调查中发现,半枫荷母树周围无更新苗,仅在1个样地有2株幼树,且均由周围母树根分蘖而来,说明半枫荷自然更新能力较弱。这可能是由于半枫荷为枫香树属(LiquidambarLinn.)和蕈树属(AltingiaNoronha)的天然杂交种[22],种子的败育率高、饱满程度低;此外,半枫荷种群规模小,大多数单株呈散生分布状态,迫使其采取自花授粉,不利于开花坐果,这也是导致其种子自然繁殖能力相对较弱的重要原因[10,23]。调查中还发现,现有的半枫荷绝大多数散生于枫香树、木荷和栲等高大乔木的中下层,难以获得充足的阳光和养分,导致其幼苗难以存活,这可能是造成半枫荷濒危的主要原因[10,15,24]。笔者对福建省内主要的半枫荷群落调查后发现,只有在3个郁闭度较低的竹林生境的半枫荷群落存在更新苗(未公布数据),而沙县天台山半枫荷群落平均郁闭度高达0.737,加之凋落物层较厚,不利于半枫荷种子萌发,间接印证了半枫荷天然更新能力弱。笔者利用SRAP和EST-SSR分子标记均得出半枫荷遗传多样性较高的结果,说明遗传多样性不是导致半枫荷濒危的主要原因[10,13]。结合本研究结果,半枫荷自身的繁育特性、群落郁闭度较高和较多负联结高大乔木挤占半枫荷生态位可能是导致其濒危的主要原因。
对半枫荷种群生态位研究发现,重要值与生态位宽度反映其在群落中的地位并非完全一致,说明植物种类的分布频度可能也是影响生态位宽度的重要因子。植物种类的重要值越大,生态位不一定越宽[25]。如在乔木层中,半枫荷虽然重要值较大(7.19%,第2位),而生态位宽度相对较小(Levins生态位宽度3.340,第4位),可能是由于栲、锥栗、山槐和赤杨叶等占据相同资源位的种类是大径级植株主要组成部分,且这些种类分布更均匀、重要值变异系数更小。锥栗的重要值和Levins生态位宽度分别为6.26%(第3位)和2.688(第14位),而山槐的重要值和Levins生态位宽度分别为1.92%(第12位)和3.941(第2位)。这是因为锥栗集中分布在P2和P3样地中,且其胸高断面积较大,具有较高的重要值;山槐虽然重要值相对较小,但其分布较锥栗更均匀,在每个样地均有分布,导致其生态位宽度较大。在灌木层中,与山槐类似,百两金、短尾越橘、南烛和朱砂根等均为生态位宽度排名较重要值更靠前,这是因为除重要值外,分布频度和重要值变异系数也是影响生态位宽度的主要因子。本研究中,半枫荷的生态位宽度较大,其在研究区内分布均匀性不高,如灌木层中仅在P1样地的1个样方紧邻分布2株半枫荷,表明其在天台山的生态适应能力一般。桫椤〔Alsophilaspinulosa(Wall. ex Hook.) R. M. Tryon〕[25]、红豆杉〔Taxuswallichianavar.chinensis(Pilg.) Florin〕[26]、四药门花〔Tetrathyriumsubcordatum(Benth.) Oliver〕[27]和海南五针松(PinusfenzelianaHand.-Mzt.)[28]的生态位宽度在群落中均较大,本研究也得出相似结果,说明生态位宽度与半枫荷濒危没有必然联系。
生态位重叠值能够反映2个种群对同一资源位的共同利用程度以及种群间占据资源位的交错程度。半枫荷群落乔木层和灌木层主要种类生态位重叠值大于0.5的种对分别占67.1%和49.7%,生态位重叠值的平均值分别为0.571和0.500,说明半枫荷与群落中主要种类的生态位重叠值较高,可能与赤杨叶和红楠等种类间存在较大的环境资源需求相似性、种间竞争较激烈以及群落稳定性较差有关。乔木层中半枫荷与赤杨叶、红楠、南酸枣、木荷、野柿、米槠、杨桐和山槐等种类不仅生态位重叠较高而且他们的生态位宽度也较大,这可能是由于这些种类在生境中资源利用谱较宽,对环境的适应能力强,对资源的利用能力也强,因此当他们出现在同一群落时,较易获得充足的资源,因而生态位容易出现较大重叠。张国强等[26]对红豆杉的研究也得到相似的结果。然而,半枫荷与甜槠、油茶和杉木等种类的生态位重叠值较小,这可能是由于这些种类对生态因子的需求与半枫荷的相似性较低、种间竞争较温和有关。与半枫荷生态位重叠值最高的红楠稍耐阴,为喜微酸性而多腐殖质土壤的速生树种,且在各样地中均与半枫荷伴生,生长速度快于半枫荷,资源竞争激烈;赤杨叶与半枫荷的生态位重叠值仅次于红楠,赤杨叶适应性较强,生长迅速,为强阳性树种,分布于常绿阔叶林中。实际调查中发现,半枫荷常分布于山谷、阔叶林及竹林中,南酸枣、野柿、米槠、格药柃(EuryamuricataDunn)、杨桐、山槐和黄牛奶树等都适生于湿润山地阳坡的阔叶林中,与半枫荷的生长环境较为相似。但生态位重叠值也只能反映出种对间资源的需求和利用的相似程度,不能完全反映其竞争关系[5]。
本研究中,与半枫荷呈正联结的种对数占总种对数的66.7%;半枫荷与乔木层主要种类的联结系数(AC)、关联度指数(OI)和结合点相关系数(PCC)检验结果基本一致。χ2检验结果表明:该群落中半枫荷与乔木层主要种类的种间关联不显著,说明半枫荷与这些种类间尚未达到稳定共存的格局。这与濒危植物长序榆[7]和黄梅秤锤树(SinojackiahuangmeiensisJ. W. Ge et X. H. Yao)[29]的种间联结性结果相似。半枫荷与木油桐、毛锥和木荷和山胡椒呈紧密正关联(AC≥0.6)。处于同一林层的种类呈正关联,如处于林冠上层的半枫荷与木油桐、毛锥、山乌桕和苦槠等,可能是由于他们在资源利用和生境需求方面的相似性,也可能是由于天台山半枫荷群落的资源有限;而处于不同林层的种类在生态位上出现重叠,种间关系表现为正关联,如半枫荷与次林层的山胡椒、木荷和福建山矾等,可能是因为他们对环境和资源的需求一致性较高。此外,虽然甜槠、树参和枫香树与半枫荷呈负关联(AC≤-0.5),但这3个种类与半枫荷生态位重叠值却较小,这可能是由于半枫荷与同林层的甜槠、树参和枫香树具有排斥性以及对环境和资源的需求存在较强的竞争关系。由于半枫荷多为单株散生分布,虽然天台山半枫荷种群是福建省内已发现的最大规模种群,但供试样本数量依然偏少,因此,本研究仅初步分析了福建天台山半枫荷群落现状和种间关系的外在生态特征,群落内种间复杂的内在生态过程还需进一步分析。
半枫荷群落稳定性交点坐标为(35.45,65.19),离稳定点(20,80)较远,其群落稳定性低于藤枣[5],但略高于受到择伐干扰的思茅松(PinuskesiyaRoyle ex Gordon)[30],同时与群落总体联结性不显著的结果相印证,也与实地调查结果相一致(该群落几无幼苗、龄级断层严重),说明天台山半枫荷群落稳定性相对较差。实地调查发现,半枫荷林下无幼苗,种群的径级结构呈纺锤型,缺少幼树及幼苗,为衰退种群,表明半枫荷幼树(幼苗)需经历高强度种间竞争才能长成大树,故随着群落的演替,半枫荷群落种间竞争将可能越来越激烈,在未来群落演替过程中,半枫荷可能会被偏阴性树种取代。栲、赤杨叶、米槠、苦槠、红楠、南酸枣、野柿、木荷、黄绒润楠(MachilusgrijsiiHance)、格药柃、豆腐柴、荚蒾、檵木、米碎花和杨梅等与半枫荷生态位重叠值较高的种类,有可能危及半枫荷的生存和繁衍,加之受自身生殖特性限制,半枫荷的种群规模呈逐渐缩小的趋势。濒危植物与群落中其他物种的正关联性越强,则种间互补性越强,可提高资源利用率,增强群落的稳定性[29,31]。为更有效地保护和利用半枫荷天然种群,应尽快建立保护小区,同时可适当疏伐上层高大乔木和开林窗以改善林下环境,适度清除与其竞争激烈的种类,如减少甜槠、树参和枫香树等显著负联结的种类,提高半枫荷种子发芽率及幼苗存活率,促进半枫荷天然种群的正常生长与更新,使半枫荷种群规模得以壮大。
致谢:野外调查得到福建农林大学林学院何宗明研究员和郑世群副教授以及顺昌县林业技术科技推广中心和沙县区林业局的大力支持,谨致谢忱!
