小兴安岭典型阔叶红松林幼树的群落结构动态
2021-12-28杨晓惠林文树吴金卓
杨晓惠,林文树,刘 曦,吴金卓
(东北林业大学 a.工程技术学院;b.林学院,黑龙江 哈尔滨 150040)
群落结构是群落中各个物种种间、种内相互关系以及物种与环境相互作用、协同进化的结果[1]。植物群落结构的动态变化包括物种多样性的变化和空间分布格局的变化等[2]。其中,植物群落物种多样性是指在环境的影响下,经过长期自然选择后的群落的复杂程度,包括群落内物种的丰富程度和物种分布的均匀程度[3]。而植物群落的空间分布格局是指群落内某一种群在长期的自然选择下,经过干扰、竞争、生境差异性和生活史策略后在其生存空间内相对静止的分布模式,表现为随机分布、均匀分布和聚集分布3 种类型[4]。群落内不同树种之间和树木不同生长阶段之间存在一定的空间关联性,即表现为空间正相关、负相关或无相关,这是由邻木之间相互作用和生境异质性等因素造成的,树木不同生长阶段之间的空间关联性也可以用于判断群落未来的演替趋势[5]。因此,通过对森林群落的物种多样性和空间分布格局的动态分析,旨在揭示森林的群落构建机制和自然演替规律[6]。
近年来,进行长期定位监测逐渐成为森林植物群落结构和生物多样性研究的主要手段[7]。建立大型固定监测样地(Forest dynamic plot,FDP)为探讨物种多样性维持机制、揭示生态学基本原理及发展等诸多问题提供了有利的条件[8-11]。1980年,美国史密斯桑尼亚热带森林研究中心首次在巴拿马巴洛科罗拉多岛(Barro colorado island,BCI)建立了50 hm2热带雨林植物监测大样地[12],随后Condit[13]、Rosindell[14]、Ricklefs[15]和Legendre[12]等学者就空间分布格局、中性理论假说、生物多样性形成及维持机制等方面取得了重要的研究进展,在世界范围内形成了热带森林生物多样性监测网络[16-18]。2004年,中国建立了森林生物多样性监测网络(Chinese forest biodiversity monitoring network,CForBio)[19],目前,该网络覆盖的主要地带性森林类型有热带雨林、亚热带常绿阔叶林、暖温带落叶阔叶林、温带针阔混交林和寒温带针叶林。金毅、邹顺、赵桔超和李建等[2,20-22]学者通过监测古田山、鼎湖山、西双版纳和长白山等固定大样地的森林植被情况,研究了森林群落结构在较长时间下的动态变化过程,为森林保护和恢复生物多样性提供了理论依据。
阔叶红松林Pinus koraiensis主要分布在我国东北东部的长白山、小兴安岭、大兴安岭和完达山等地,群落物种繁多、结构复杂,稳定性强,作为顶级植被群落在东北针阔叶混交林中占有重要地位。但是,由于木材的大量需求,原始阔叶红松林遭到采伐,受到人为干扰,森林的生物多样性减少,生态环境恶化,生态功能下降[23]。目前,对于阔叶红松林的研究主要集中在对乔木群落的结构特征以及优势树种的空间格局及其关联性进行的分析[22,24],而针对小兴安岭地区阔叶红松林群落内幼树(1 cm ≤DBH<5 cm)群落结构和空间格局,较少进行长期的动态监测研究和分析。幼树阶段是植物生活史中动态变化最明显的阶段,幼树与小树和大树之间相互作用,受到竞争、干扰、光照、密度制约等环境因素,在不同空间尺度上具有不同的相关性[25]。群落幼树的物种组成和结构特征的动态变化决定着森林未来的演替趋势。小兴安岭作为我国阔叶红松林的主要分布区之一,研究其群落幼树结构动态变化,有利于恢复生物多样性,维护生态系统结构和功能。
因此,本研究以小兴安岭典型阔叶红松林固定监测样地为研究对象,利用2010年初测、2015和2020年复测的群落调查数据,分析此区域内1 cm ≤DBH<5 cm 的幼树10 a 间群落物种多样性动态变化,并分析3 次调查幼树的空间分布格局如何变化以及3 次调查结果下幼树同小树、大树的空间关联性是如何变化的,旨在为森林群落动态长期监测研究、维持群落结构、恢复群落生物多样性和合理经营管理提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于黑龙江省伊春市带岭区凉水国家级自然保护区(47°06′49″~47°16′10″N,128°47′08″~128°57′19″E),海拔为280~707.3 m,平均坡度为10°~15°,属于典型的低山丘陵地貌,年平均气温-0.3℃,冬长夏短,具有明显的温带大陆性季风气候,年平均降水量为676.0 mm,积雪期130~150 d,无霜期100~120 d,地下水资源丰富,研究区内物种丰富,几乎包含了小兴安岭山脉的所有森林植被类型,土壤为暗棕壤、草甸土、沼泽土和泥炭土[23]。研究区域内主要树种有红松Pinus koraiensis、冷杉Abies fabri、紫椴Tilia amurensis、青楷槭Acer tegmentosum、色木槭Acer mono、枫桦Betula costata、花楷槭Acer ukurunduense和云杉Picea koraiensis[26]。
1.2 样地设置与调查
小兴安岭阔叶红松林固定监测样地面积大约为30 hm²(380 m×780 m),于2010年在研究区内参照CTFS(Center for tropical forest science)的建设样地技术规范建立。采用全站仪将30 hm²大样地划分为20 m×20 m 的连续样方,再采用相邻网格法将20 m×20 m 样方细分为16 个5 m×5 m 的调查单元,对样地内所有DBH ≥1 cm 的木本个体进行每木检尺[26]。2015 和2020年分别对研究区进行复查,同时对新增树木进行补充编号,对死亡个体的状态进行更新。由于随着坡位的升高,含水量降低,土壤有机质减少,植物个体分布数量减少[27],本研究在固定监测样地低海拔地区选取研究样地。Zenner[28]研究发现,在森林结构调查中样地面积为2 500 m²时,群落的结构属性和指标趋于稳定;在研究区固定监测样地选取最小样地面积为40 m×40 m 时,几乎涵盖了群落的大部分物种,也可以比较全面地反映林分情况[27]。综合以上因素,本研究在固定监测样地低海拔区域内选取生长情况较好的200 m×200 m 样地进行后续的研究分析。此样地最高海拔是392 m,最低海拔341 m,平均坡度为6°。以所选样地东北点为原点,以40 m×40 m 为基本单元设置样方,沿着样地边长依次增加40 m,样方面积分别为40 m×40 m、80 m×80 m、120 m×120 m、160 m×160 m、200 m×200 m,分别计算不同尺度下的物种多样性。
1.3 研究方法
1.3.1 胸径等级划分
树木的胸径结构可以反映树木生长年龄[29],按照胸径结构将所测量的DBH ≥1 cm 的乔木划分为3 级:幼树(1 cm ≤DBH<5 cm)、小树(5 cm ≤DBH<20 cm)和大树(DBH ≥20 cm)[24],采用单因素方差分析法分析不同年份对不同生长阶段胸径的影响。
1.3.2 群落结构特征
重要值(Pi)是表示物种在群落中重要性的指标,计算样地内物种重要值公式如下[30]:

物种多样性指数是表示群落物种多样性程度的指标,可以用来判断群落和生态系统稳定性,其中n表示此区域内物种的数量,各指数计算公式[31]如下。
丰富度指数(S):

Shannon-Wiener 多样性指数(H'):

Simpson 优势度指数(D):

Pielou 均匀度指数(E):

指数变化率(R)是表示群落物种多样性指数在不同调查时间段的变化率,其中X表示指数值,t表示调查的时间点,T表示两次调查的时间间隔,变化率公式如下:

1.3.3 空间分布格局
以样地内幼树、小树和大树的空间点坐标为基础,运用单变量O-ring 函数分析样地内幼树的空间分布格局,对于单变量O-ring 分析来说,使用完全随机零模型分析幼树的空间分布格局[23];运用双变量O-ring 函数分析幼树与小树、幼树与大树的空间关联性,对于双变量O-ring 分析来说,考虑到幼树和小树、幼树和大树之间相互作用,采用前提条件零模型,认为大树和小树对幼树的生长和布局具有影响,使小树和大树位置不变,对幼树的空间位置进行随机化处理,分析两个研究对象之间的关联性[32]。本研究通过100 次Monte Carlo 函数拟合计算99%置信区间,空间尺度为100 m,步长为1 m。本研究规定小尺度为0~33 m,中等尺度为34~66 m,大尺度为67~100 m。
根据Wiegand[32]等的定义,单变量O-ring 函数(Ow(r))中,Riw(r)是以点i为圆心,r为半径,w为宽度的圆环,Point[X]表示在区域X内的点的个数,Area[X]表示区域X的面积的大小;双变量O-ring 函数((r))中,n1为物种1 的点总数,(r)表示以物种1 第i点为圆心,r为半径,w为宽度的圆环,Point2[X]表示在区域X中物种2点的个数,计算公式如下:
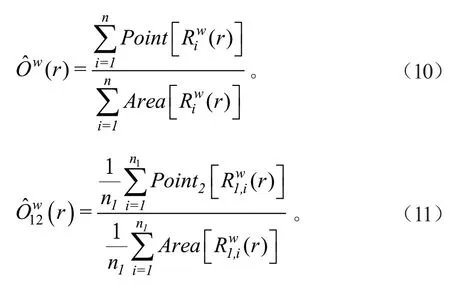
1.3.4 数据处理
本研究使用Excel 2010 软件进行统计分析,使用SPSS 23.0 软件对不同生长阶段胸径进行单因素方差分析(One-way ANOVA)和Duncan 法进行方差分析(α=0.05),使用Programita for point pattern analysis 2010 软件进行幼树的空间分布格局及幼树和小树、幼树和大树的空间关联性分析,利用Sigmaplot 12.5 软件进行作图。
2 结果与分析
2.1 不同生长阶段乔木物种及数量变化
由群落调查结果可知,2010、2015 和2020年分别在200 m×200 m 研究样地内监测到胸径≥1 cm的乔木物种数和个体数分别为20 种3 813 株、20种4 110 株、20 种3 971 株。不同调查时间点下乔木个体在不同生长阶段的数量变化如表1所示。其中,幼树在3 次调查中分别为169、173 和200株·hm-2,小树分别为621 和567 株·hm-2,大树分别为210、233 和227 株·hm-2。2015年从幼树进入到小树阶段的个体有78 株·hm-2,从小树进入到大树阶段的个体有22 株·hm-2;2020年从幼树进入到小树阶段的个体有56 株·hm-2,从小树进入到大树的个体有18 株·hm-2。
对于针叶树来说,2010—2015年不同生长阶段乔木数量均有增加,幼树、小树和大树分别新增20、10、6 株·hm-2;2015—2020年,不同生长阶段乔木数量均呈减少趋势,幼树、小树和大树分别减少5、4、14 株·hm-2。对于阔叶树来说,2010—2015年不同生长阶段乔木数量呈现不同的变化趋势,幼树数量减少15 株·hm-2,小树和大树的数量分别增加37、18 株·hm-2;2015—2020年,小树数量减少51 株·hm-2,幼树和大树的数量分别增加32、8 株·hm-2。3 次调查中,阔叶树的物种数均大于针叶树,小树的物种数多于幼树和大树。对于针叶树来说,不同生长阶段物种数随着时间的增加没有发生改变;对于阔叶树来说,2010年幼树阶段未见山杨Populus davidiana,2015 和2020年幼树阶段未见山杨、旱柳Salix matsudana和山桃Amygdalus davidiana,在3 次调查中大树阶段均未见鼠李Rhamnus davurica和暴马丁香Syringa reticulata。
2.2 幼树群落结构特征动态变化
2.2.1 幼树物种组成
由表1可知,3 次调查中样地内幼树物种数分别为19、17 和17 种。幼树优势物种组成数量特征的动态变化如表2所示。可以看出,重要值排名在前12 位的优势物种组成基本保持不变,2015和2020年,仅白桦Betula platyphylla(2010年重要值0.057)退出优势位,枫桦进入优势位,其他优势物种不变。12 个优势物种的重要值总和分别为0.88、0.93 和0.95。3 次调查中,花楷槭、青楷槭和枫桦的重要值持续增加,色木槭、冷杉、云杉、红松的重要值呈现先增加后减少的趋势,毛赤杨Alnus sibirica、暴马丁香、紫椴、稠李Padus racemosa、春榆Ulmus davidiana的重要值先减少后增加。从幼树物种多度来看,3 次调查中色木槭、冷杉、花楷槭、青楷槭幼树的数量相对较多,总和占幼树总数量均超过69%。其中,2015 和2020年冷杉个体数量达到最大。由于个体数量较多,历次调查中色木槭、冷杉、花楷槭、青楷槭幼树的胸高断面积也相对较大,总和分别占样地总胸高断面积的70.19%,72.50%和73.20%。
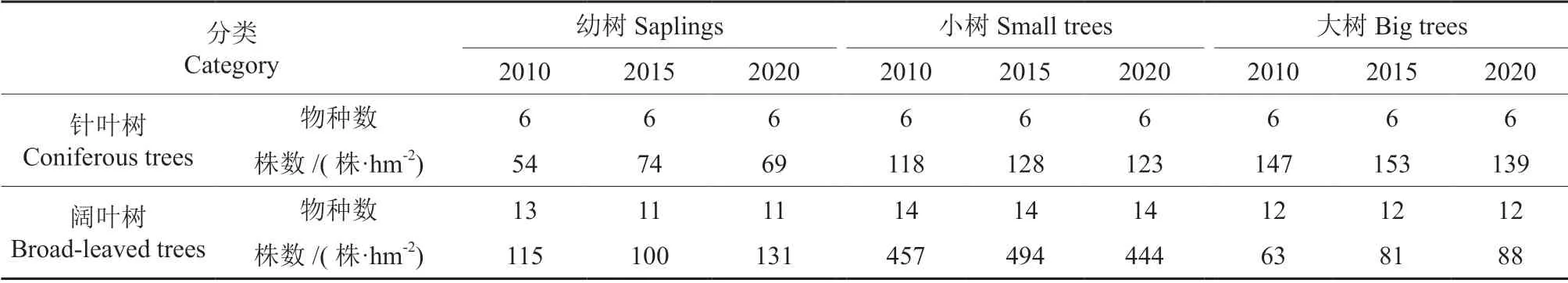
表1 2010-2020年典型阔叶红松林乔木层不同生长阶段物种及数量变化Table 1 Changes of tree species composition and quantity in different growth stages in typical broad-leaved korean pine forest from 2010 to 2020
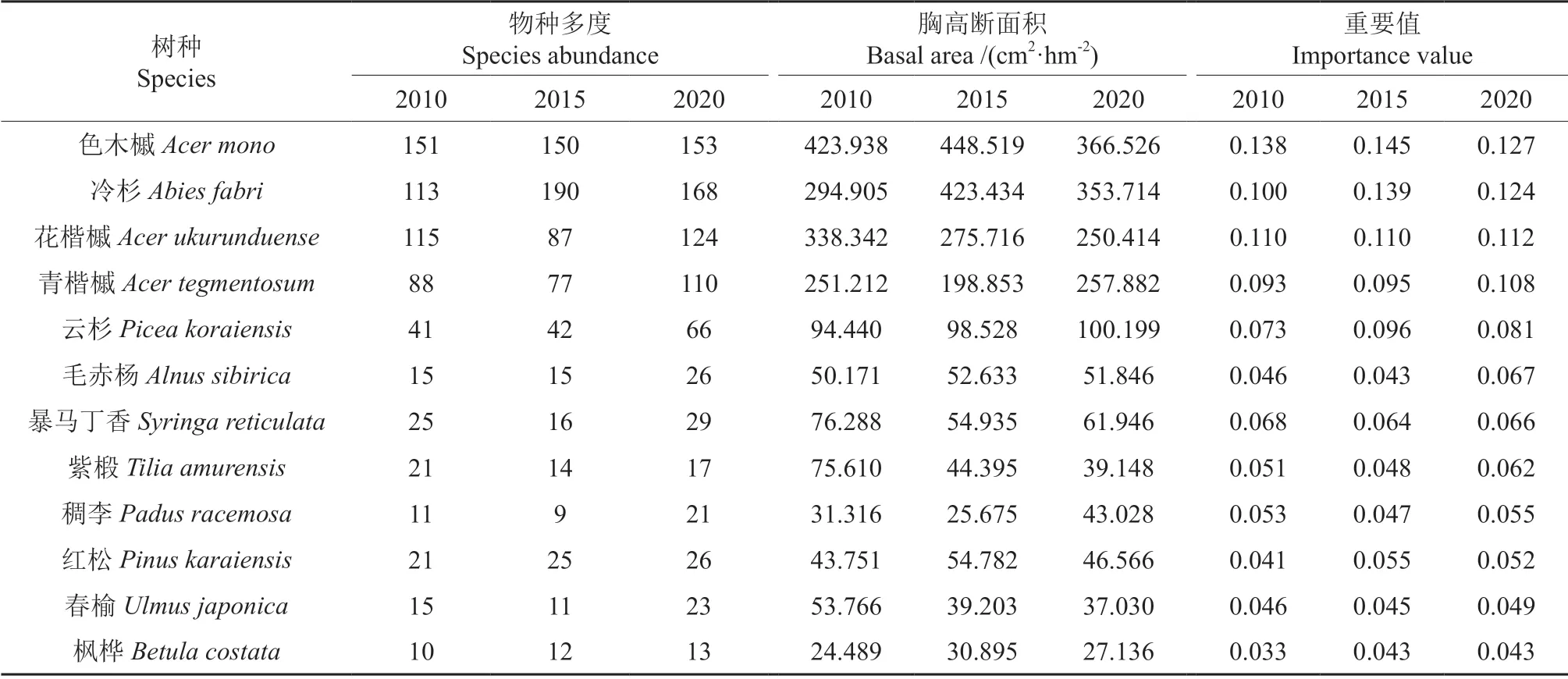
表2 幼树优势树种物种组成数量特征的动态变化Table 2 Dynamic changes in quantitative characteristics of species composition of dominant sapling tree species
2.2.2 幼树物种多样性
不同尺度样地下幼树物种多样性指数如表3所示。可以看出,在3 次调查中,随着空间尺度的增加,物种丰富度指数、Shannon-Wiener 多样性指数和Simpson 优势度指数总体上呈现先急剧增加后趋于平缓的趋势,Pielou 均匀度指数呈现先明显下降后趋于平缓的趋势。当尺度由40 m×40 m增加到80 m×80 m 时,各指数的变化速率最大。另外,在200 m×200 m 的空间尺度下,随着时间的增加,Shannon-Wiener 多样性指数和Simpson优势度指数呈先减小后增加的趋势,Pielou 均匀度指数持续增加。
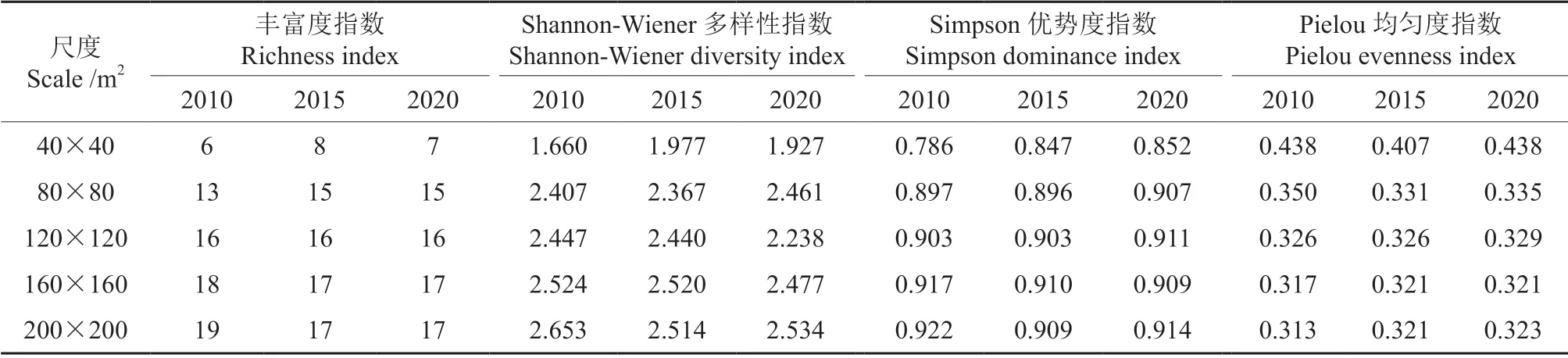
表3 不同尺度幼树物种多样性指数Table 3 Sapling species diversity index at different scales
2.2.3 群落径级结构
200 m×200 m 样地内林木的径级分布情况由图1所示。不同调查年份下,幼树、小树和大树的平均胸径分别在3.14~3.73,10.58~10.90,36.70~38.34 cm 之间。与2010年相比,2015年幼树的平均胸径有所增加,但是变化不显著(P=0.60>0.05),2020年幼树的平均胸径与2010 和2015年相比均显著减少(P=0.00<0.05);随着时间的增加,小树的平均胸径持续增加,其中2020年小树的平均胸径与2010年相比显著增加(P=0.04<0.05),大树的平均胸径也有所增加,但是增加的幅度不显著(P=0.80>0.05)。不同调查年份间样地内所有林木的平均胸径分别为15.11、15.58 和15.60 cm,群落整体处于稳定状态。
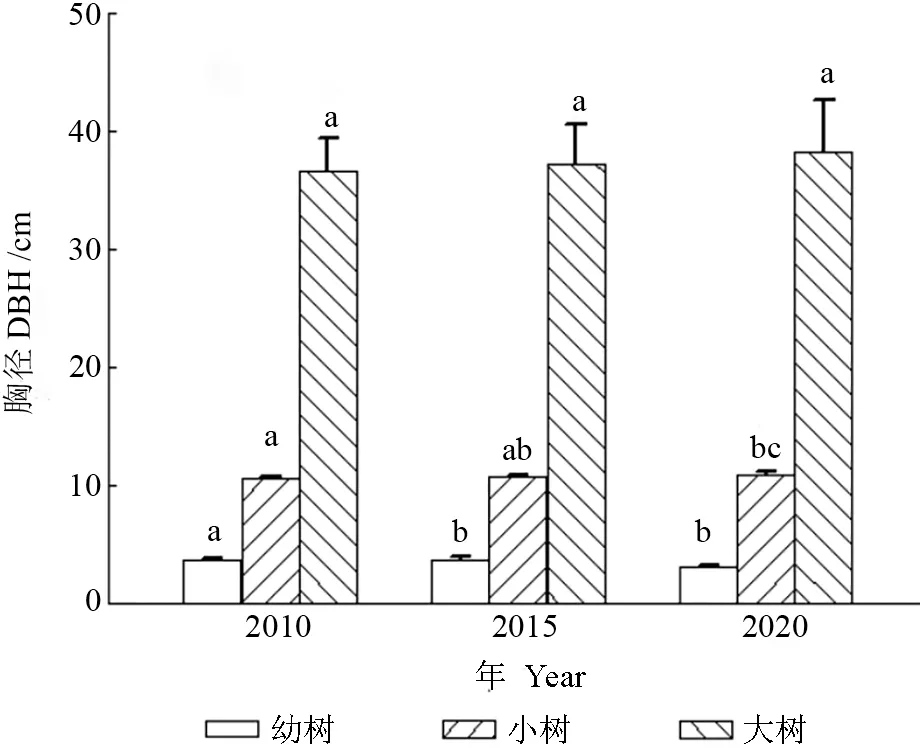
图1 样地内林木的径级结构Fig.1 Diameter class structure of trees in the sample plot
2.3 幼树的空间分布格局及其空间关联性动态变化
2.3.1 幼树的空间分布格局
不同调查年份下200 m×200 m 阔叶红松林样地内幼树的空间分布情况如图2所示。可以看出,2020年样地内的幼树空间分布比较均匀,2015年样地内的幼树空间分布和2010年幼树空间分布类似,在部分区域上存在聚集分布。运用单变量O-ring函数对样地内幼树分布进行空间点格局分析,结果如图3所示。由图3a 和图3c 可以看出,2010年和2020年幼树的空间分布格局均在小尺度表现为聚集分布,随着尺度的不断增大,种群的空间分布格局变为随机分布,但在部分尺度存在均匀分布;2015年幼树的空间分布格局在小尺度表现为聚集分布,但随着尺度的不断增大,种群的空间分布格局变为均匀分布(图3b)。
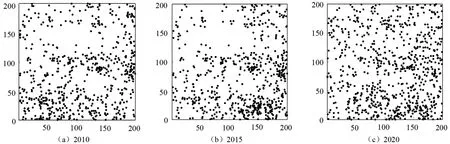
图2 样地内幼树的分布点Fig.2 Distribution of saplings in the sample plot

图3 样地内幼树的空间分布格局Fig.3 Spatial distribution patterns of saplings in the sample plot
2.3.2 幼树与小树、大树的空间关系
运用双变量O-ring函数对样地内幼树与小树、大树的空间关系进行点格局分析,结果如图4所示。可以看出,在3 次调查中,幼树和小树的空间关联性类似,在小尺度上呈正相关,随着尺度的增加,正相关性减弱,两者变为无相关性,但是在中等尺度上和大尺度上存在部分负相关性;2010和2015年幼树和大树在小尺度上呈负相关,在大尺度上呈正相关性,在不同尺度均存在部分无相关性,2020年幼树和大树整体上呈无相关。2010年幼树和小树之间在0~11、14、17 和23 m的尺度上呈正相关,在56~67、70、72~74 和77 m 的尺度上呈负相关性,在其余尺度上无关联;幼树和大树之间在76、82~85、87~100 m 的尺度上呈正相关,在8~25、30~31、44、52和58 m 的尺度上呈负相关,在其余尺度上无关联。2015年幼树和小树之间在0~20、23、33、35 和37 m 的尺度上呈正相关,在57~79 m 呈负相关,在其余尺度上无关联;幼树和大树之间在73~100 m 的尺度上呈正相关,在8、11~13、16~37、40、42、44、47、49、51~52、55~56 和58 m 的尺度上呈负相关,在其余尺度上无关联。2020年幼树和小树之间在0~12、14~20、23~24、27、38 m 的尺度上呈正相关,在63~64、66~72、76~77 和80 m 的尺度上呈负相关,在其余尺度上无关联;幼树和大树之间在4 m 和83 m 的尺度上呈正相关,在其余尺度上无关联。
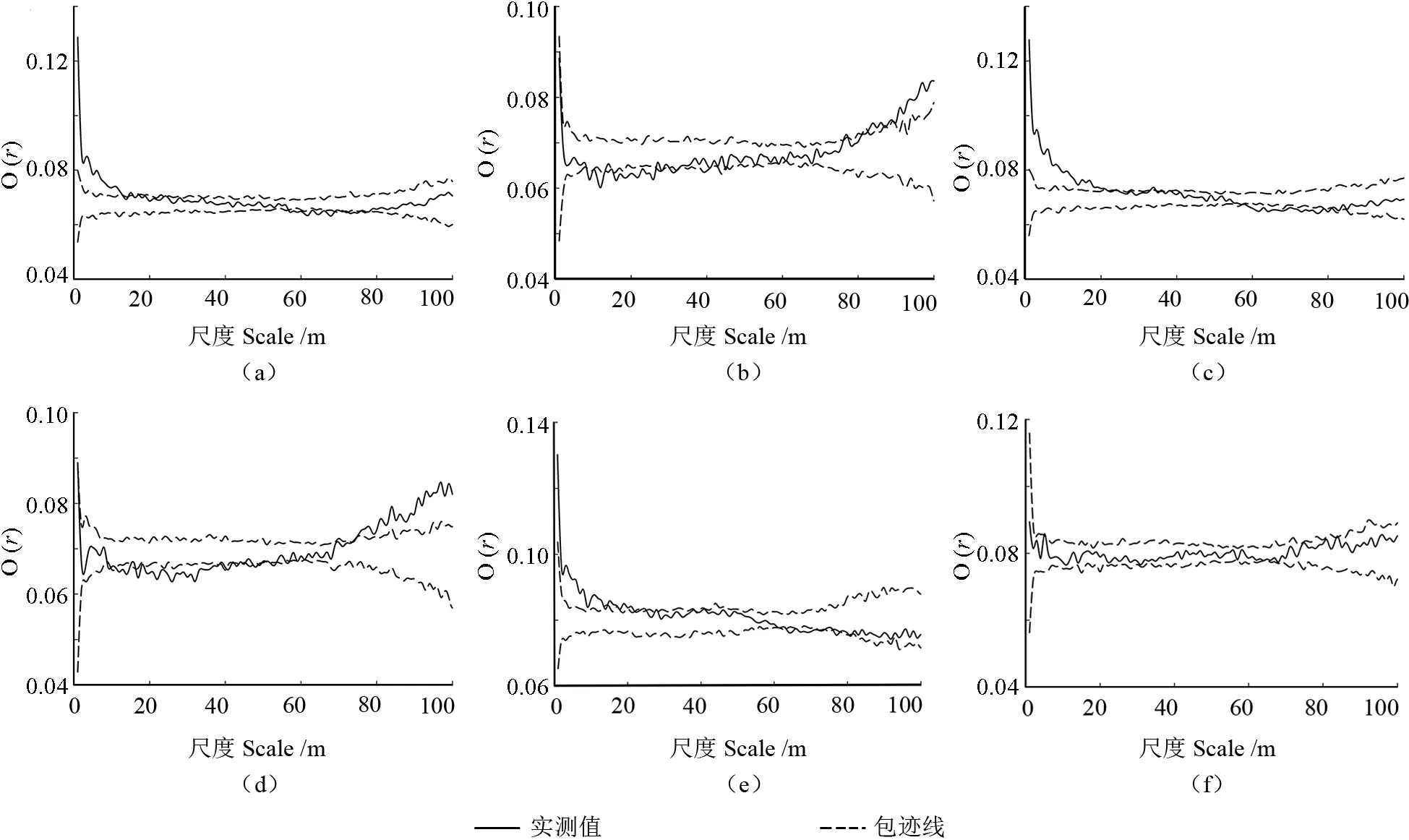
图4 样地内幼树与小树、大树的空间关联性。(a) 2010 幼树—小树,(b) 2010 幼树—大树,(c) 2015 幼树—小树,(d) 2015 幼树—大树,(e) 2020 幼树—小树,(f) 2010 幼树—大树。Fig.4 Spatial correlation between saplings and small trees and big trees.(a) 2010 saplings and small trees,(b) 2010 saplings and big trees,(c) 2015 saplings and small trees,(d) 2015 saplings and big trees,(e) 2020 saplings and small trees,(f) 2020 saplings and big trees.
3 讨 论
3.1 群落幼树物种多样性的特征
研究森林群落结构和动态变化以及群落物种多样性维持机制一直是森林生态学的热点问题[31]。由3 次调查结果可知,乔木物种数保持不变,但个体数呈现先增加后减小的趋势;2015 和2020年小树新增个体数均高于转移个体数,但是2020年较高的枯死率逆转了小树的增长趋势,这可能与2020年研究区受到多种恶劣天气影响有关;2010年幼树阶段未见山杨,2015 和2020年幼树阶段未见山杨、旱柳和山桃,这是由于在小树和大树阶段旱柳、山桃和山杨为偶见种[26],种子数量较少,且旱柳和山杨属于阳性树种,随着森林的演替会逐渐衰退,也可能与其种子传播困难及生物学特性有关[33];在大树阶段未见鼠李和暴马丁香,可能与其生物学特性有关,因为鼠李和暴马丁香均为小乔木,无法生长到大树阶段。小树的株数最多,表明林下树种丰富,群落在进行进展演替;阔叶树种物种数和植物株数都较大,表明此区域是典型的阔叶红松林,阔叶树种在此群落中占有重要地位。这与侯红亚等[26]在小兴安岭阔叶红松林物种组成研究中的结果是一致的,符合地带性群落分布特征,体现了群落的稳定性。
通过对小兴安岭阔叶红松林幼树的物种组成和群落结构对比分析可知,幼树优势物种基本保持不变,物种组成的变化受到种子库、干扰以及研究时间间隔等因素的影响[12],研究中的群落处于演替中后期,研究的时间间隔相对较短,且人为干扰较少,因此幼树物种变化不明显。这与徐玮泽等[34]在长白山阔叶红松林树木种群动态研究中物种组成没有明显变化的结果是一致的,表明演替处于一个相对平衡的阶段。白桦的重要值降低,花楷槭、青楷槭和枫桦的重要值持续增加,这与李建等[22]在长白山阔叶红松林群落对其演替实质的分析是一致的,表明随着演替进行,阴性树种会取代阳性或半阴性树种,耐阴性是种群在大多数森林动态演替过程中长期存在的关键因素[35],群落处于正向演替过程中,而红松幼树的重要值相对较小,这可能是由于红松幼苗成活率低,球果无法借助重力和风力来进行传播等原因造成的[33]。3 次调查中色木槭、冷杉、花楷槭、青楷槭幼树的物种多度和胸高断面积相对较多,对群落的贡献均远大于群落其他物种,这在一定程度上反映了种群在短期内的稳定性[20]。随着尺度的增加,不同物种多样性指数出现了不同的结果,是因为不同物种多样性指数具有不同的性质[36],当尺度由40 m×40 m 增加到80 m×80 m 时,各指数的变化速率最大,随后变化速率趋于平缓,这说明群落的物种多样性在一定尺度范围内,会明显受到尺度的影响。随着尺度增加,Pielou 均匀度指数下降,是因为空间异质性会随着尺度的增加而增大。这与郭孝玉等[36]对石栎次生林多样性指数的尺度效应研究结果是一致的。3 次调查结果计算的丰富度指数、Shannon-Wiener 多样性指数、Simpson 优势度指数和Pielou 均匀度指数的变化主要是由偶见种引起的,说明群落处于稳定变化中,偶见种对维持群落生物多样性机制具有重要影响。随着时间的增加,小树和大树的平均胸径均有增加,但2020年幼树的平均胸径显著减少,说明幼树阶段易受环境和生物因素影响而引起死亡[25],其动态特征随时间的变异性较大。不同调查年份间样地内所有林木的平均胸径分别为15.11、15.58 和15.60 cm,该群落为相对稳定的森林群落。
3.2 群落幼树的空间分布格局
种群的空间分布格局受到生境条件、种内种间关系和生态习性等因素影响,对空间尺度格局具有明显的依赖作用[37],本研究运用O-ring 函数对幼树的空间分布格局进行分析,3 次调查幼树的空间分布格局类似,均在小尺度表现为聚集分布,随着尺度的不断增大,种群的空间格局变为随机分布或均匀分布,这与Stoll 等[38]在竞争对植物个体间隔的影响研究中、Kubota 等[39]在对阔叶林和针叶林动态格局分析的研究中、Law 等[40]在植物空间点格局理解的研究中以及陈贝贝等[41]在长白山阔叶红松林幼苗空间分布研究中的结果是一致的;这种分布格局可能是由种群的生物学特性造成的,同一种幼树个体对环境的需求基本一致,因此在小尺度上聚集分布,共同抵抗不利条件,增加对光照、水分等资源的竞争力,提高此种幼树成活率;随着尺度的增大,竞争减弱,群落幼树表现为随机分布或均匀分布[41]。但是在2015年中等尺度和大尺度范围内出现均匀分布较多,可能是因为与2010年相比较,2015年群落进行正演替,群落空间格局更加稳定和理想[22];与2020年相比较,2020年受到多种台风天气影响,树木死亡较多,枯倒木对幼树分布产生影响,均匀分布减少。
3.3 群落不同林层的空间关联性
分析幼树与小树、大树的空间关联性对于更深层次了解不同林层之间相互作用和群落稳定性变化具有重要意义[42]。不同林层的关联性体现了对外界资源的分配利用能力,正相关则表明植物互利共生,生长条件相似,存在生态位重叠;负相关则表明植物之间存在竞争排斥,生长条件存在差异[5]。运用O-ring 函数对幼树与小树、大树的空间关联性进行分析,由图4可得,在3 次调查中,幼树和小树的空间关联性类似,在小尺度上呈正相关性,随着尺度的增加,相关性减弱,两者变为无相关性,但是在中等尺度上和大尺度上存在部分负相关性。这可能是因为幼树生长初期比较脆弱,在小尺度上两个阶段的树木竞争较小,由于处在不同的生长阶段,生态位存在分离,各自利用不同的资源而和谐共存,而在中等尺度上和大尺度范围内上层林木具有较强的竞争资源能力,从而在某种程度上呈一定的负相关性,说明种群的空间分布格局受到生境异质性和扩散限制的影响,不同尺度下林分具有不同的配置方式。幼树和大树在大多数尺度上呈无相关性和正相关性,这可能是由于阔叶红松林幼树数量丰富,林分相对成熟,林下土壤湿润,养分条件良好,也可能是因为大树产生大量的种子,受到种子传播限制影响,幼树多集中在大树周围[41],说明这两者之间也存在竞争较小,可以很好地共存。但是在小尺度和中等尺度上仍然存在负相关,可能是因为幼树和大树形体差异很大,较大的个体可能对较小的个体产生排斥。这与舒兰等[43]在帽儿山天然次生林内对主要木本植物空间分布格局及其更新特点的研究中关于幼苗和大树、中树、幼树的空间关联性的研究结果是一致的。3 次调查中幼树和小树、大树的变化趋势相似可能是因为幼树和小树、大树之间形成特定关系,在分布格局上表现为稳定性[44]。但是幼树与小树、大树在整个研究尺度内均存在负相关性,说明了此群落仍然需要处于进展演替中,需要一段时间才能使其更复杂化和稳定化[43]。
本研究基于2010、2015 和2020年对小兴安岭典型阔叶红松林固定监测样地的调查数据分析群落幼树的非空间结构和空间结构的动态变化,对今后阔叶红松林的演替趋势,优化森林结构、恢复生物多样性具有重要意义。但是北方树种生长相对缓慢,本研究研究结果并未发现群落演替中“质”的变化,群落幼树的群落结构和空间格局仍然处于变化之中,同时在短期的测量中由于人为测量可能会存在偏差,所以需要对大样地进行持续监测研究,在长期监测并多次复查的情况下校正数据,持续观察其动态变化。另外,对于该样地的研究仍处于起始阶段,后期仍然需要对植物个体的生长与死亡、植物的谱系结构、种子扩散与幼苗存活机制、环境因素与群落动态的相互作用、植物的功能性状等方面开展研究,以探讨生物多样性的形成和维持机制。
4 结 论
本研究以小兴安岭典型阔叶红松林固定监测样地的多年调查数据(2010、2015 和2020年)为基础,选取了低海拔生长情况较好的200 m×200 m的研究样地,分析群落幼树非空间结构和空间结构的动态变化,得出主要结论如下:
1)由3 次调查结果可知,乔木物种数保持不变,但个体数呈现先增加后减小的趋势;阔叶树的物种数和株数均远大于针叶树,阔叶树在此群落中占有重要地位,幼树优势物种组成基本保持不变。随着尺度的增加,物种多样性越来越丰富,而空间异质性增大,均匀度越来越差;群落中的偶见种引起物种多样性指数变化,在维护群落生物多样性方面起着关键作用。随着时间的增加,小树和大树的平均胸径均有增加,幼树的平均胸径先增加后显著减少,表明幼树阶段易受环境和生物因素影响而引起死亡;不同调查年份间样地内所有林木的平均胸径分别为15.11、15.58 和15.60 cm,说明该群落为相对稳定的森林群落。
2)3 次调查幼树的空间分布格局类似,均在小尺度表现为聚集分布,共同抵抗环境的不利因素,提高物种整体的存活率。随着尺度的增大,竞争性减弱,种群的空间格局变为随机分布或均匀分布,这可能是由种群的生物学特性造成的。
3)在3 次调查中,幼树和小树的空间关联性类似,在小尺度上呈正相关性,竞争较小,相互依赖,随着尺度的增加,相关性减弱,两者变为无相关性,但是在中等尺度上和大尺度上存在部分负相关性,这可能是因为在中等尺度上和大尺度范围内上层林木具有较强的竞争资源能力;幼树和大树在大多数尺度上呈无相关性和正相关性,可能是由于种子传播限制的结果;在小中尺度上存在负相关,说明在小中尺度上由于个体差异较大,大树对幼树也具有排斥。
