青藏高原多年冻土区活动层土壤入渗特征及机理分析
2021-12-17王一博吕明侠赵海鹏高泽永
王一博, 吕明侠, 赵海鹏, 高泽永
(1.兰州大学资源环境学院,甘肃兰州 730000; 2.中国科学院西北生态环境资源研究院冻土工程国家重点实验室,甘肃兰州 730000)
0 引言
土壤入渗是指降雨或者灌溉后,部分水分从土壤表层渗入深层土壤的过程,是土壤水文循环过程中水分运移的重要组成部分,它将陆地上的表层水流和地下水作为陆地水循环的一部分联系起来[1-2]。土壤入渗能力作为一个关键的水文参数,可以作为衡量土壤退化和干旱潜力的指标[3],同时也可以间接反映植被的生长发育、土壤的涵养水源以及抗侵蚀的能力[4]。土壤入渗能力可以通过初始入渗率、稳定入渗率、平均入渗率、入渗深度、累计入渗量等参数进行评估[5-6]。土壤入渗能力的量化对于草地、森林和农田等生态系统的承载力研究、明确地表径流的调节机制和土壤侵蚀的防治策略均具有十分重要的意义[2,7]。
影响土壤入渗能力的因子较多,如土壤质地、结构、矿物组成、土壤水分等土壤特性,植被类型、根系直径大小以及分布情况等植被特征[8]。目前,已有大量的研究对这些要素之间的关系进行了探讨。Alaoui 等[9]对瑞士高原4 种典型草原土壤的水文参数进行了研究,发现体积密度与大孔隙度之间的相互作用可以促进水的渗透。郑凯利等[10]对若尔盖湿地土壤入渗性能的影响因素分析时,发现土壤有机碳含量、含水量、毛管孔隙度和大于5 mm 水稳性团聚体与土壤入渗能力呈显著正相关关系。Scanlan 等[3]发现植物根系会堵塞土壤孔隙,降低土壤入渗率。Wu 等[11]发现与裸地相比,根生物量较大的人工草地更有效地提高了矿区土壤的入渗能力。Liu 等[2]发现干旱草地土壤入渗能力与土壤含水量呈负相关,与地下生物量呈正相关,而且Cui等[8]研究发现,在半干旱草地土壤中,细根较土壤含水量对土壤入渗潜力的决定作用更强。关于植被特征、土壤特性以及土壤入渗性能的研究尽管众多,但是对于青藏高原多年冻土区不同植被类型土壤入渗规律的了解依然较少。
青藏高原被誉为“亚洲水塔”和“世界第三极”,对气候变化非常敏感[12]。高寒草地占高原总面积的60%以上,根据不同的土壤和植物种类,可以将高寒草地分为三个主要的亚类——高寒沼泽草甸、高寒草甸和高寒草原[13],这些生态系统对全球气候变化非常敏感[14]。近年来,由于气候变暖,青藏高原活动层厚度增大,冻土面积减小,冻土厚度明显减小,而多年冻土对目前生态系统的维持具有非常重要的作用[15-16]。高山植被在短期内难以适应如此剧烈的气候、环境变化,出现了多种退化现象,这已经明显地影响了该地区的环境,改变了其水文循环以及水源涵养功能[13,16]。而分析不同植被类型土壤入渗能力的差异,对于认识气候变暖背景下,多年冻土区产流机制、地表水与地下水的转换、预测高寒生态系统变化及其保护具有重要意义[14]。本研究的具体目标是:定量描述高寒草原、高寒沼泽草甸、高寒草甸土壤的入渗能力,分析不同植被类型的土壤间入渗过程的差异;阐明高寒植被在土壤水分入渗中的作用机理;比较不同入渗模型在高寒草地植被类型土壤入渗过程中的适用性。
1 材料与方法
1.1 研究区概况
研究区位于青藏高原腹地风火山流域内,北麓河一级支流左冒西孔曲(图1),属于典型的高原高寒干旱半干旱气候,区域内多年冻土与地下冰较为发育。多年冻土平均厚度为50~120 m,活动层厚度在0.8~2.5 m 之间;年平均气温-5.2 ℃,极端最高气温为24.7 ℃,极端最低气温为-38.5 ℃;降水主要集中于6—8 月,年均降水量为269.7 mm;年均水面蒸发量为1 447.9 mm[17]。研究区主要发育四套不同世纪的沉积地层,分别有晚白垩世、老三纪、中新世和第四纪的砂砾岩、砂岩、泥岩和沉积地层[18]。活动层土壤类型主要为第四纪沉积物,包括红砂土、潜育土和黑钙土[13]。活动层高寒草甸为该区域主要的植被类型,建群种多为寒冷中生、湿中生的密集短根茎蒿草植物。植被群落以矮嵩草(Kobresia humilis)、高山嵩草(Ko-bresiapygmaea)和线叶嵩草(Kobresiacapillifolia)等寒生植物为主[19]。
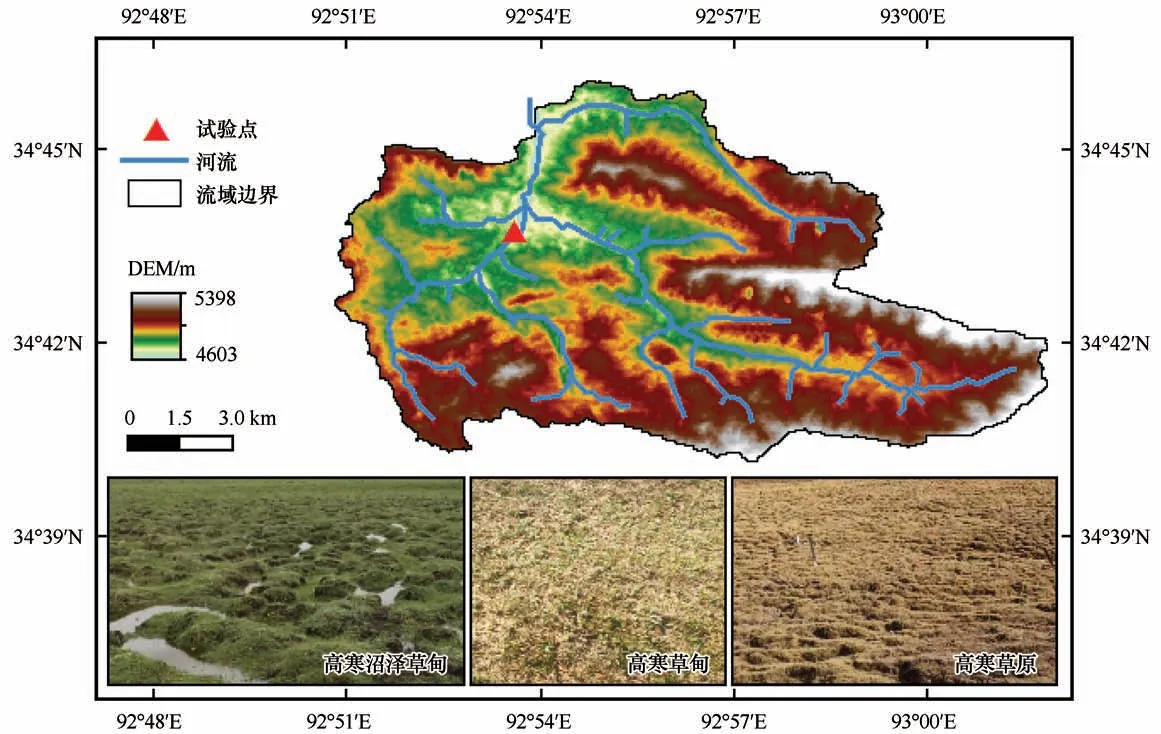
图1 研究区概况Fig.1 Situation of the study area
1.2 实验设计与土壤采样
在青藏高原多年冻土区选择植被生长良好、坡度相近、位置靠近、无人类活动干扰的高寒沼泽草甸、高寒草甸、高寒草原植被类型样地为试验样地,以裸地为对照组进行土壤入渗试验和现场采样。植被类型不同,其对应土壤剖面(0~100 cm)的土壤质地、有机质含量、土壤结构等土壤的基本性质不同[20]。在每个试验样地开挖20 cm×20 cm×50 cm 的土柱,在每10 cm 深度上用采样袋分装土样,然后带回实验室自然风干后用于地下生物量和土壤基本性质的测量。同时,在相应的深度处(10 cm 间隔),用环刀沿三个随机的方向采集土样,然后密封带回实验室用于测定土壤干密度和土壤初始含水量。
1.3 实验室分析
用烘干法(105 ℃)测土壤初始质量含水量和土壤干密度;土壤粒径采用Malvern Mastersizer 2000激光粒度仪测定,测量范围为2~2 000 μm,误差小于2%;有机质通过重铬酸钾容量法测定。提取的植物根系在烘箱中65 ℃恒温条件下烘干72 h,运用称重法测定根生物量。
1.4 土壤入渗试验
双环入渗仪是由改进的马里奥特容器(高:180 cm,内径:14 cm)和20 cm 高的圆形同心双套环(内、外环的直径分别为25.7 cm 和64.0 cm)构成的,在水头保持不变的情况下运用达西定律测定土壤一维入渗过程的变化规律。本研究在每个试验样地采用精度高而应用广泛的双环入渗仪在青藏高原多年冻土区活动层土壤开展土壤入渗过程试验[21]。土壤入渗试验中,首先将水向内、外环同时、缓慢地注入,并在整个试验过程中保持内、外环水位相齐并不变,分别在0 min,1 min,3 min,5 min,…,的时刻记录数据,直到变化率达到稳定结束试验。土壤水分入渗率计算公式如下:
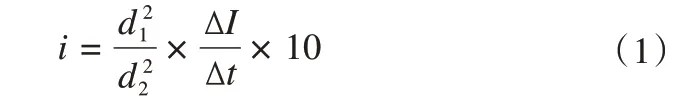
式中:i土壤入渗速率(mm·min-1);d1、d2分别为内环和外环的直径;ΔI为时间间隔是Δt的时间段内的累积入渗量;Δt为入渗时间间隔,10为转换系数。
根据活动层土壤入渗过程的试验结果,发现30 min后不同植被类型土壤的入渗率均逐渐达到稳定。因此,根据土壤入渗速率的变化情况,将入渗过程划分为三个阶段:分别是瞬变阶段(0~10 min)、渐变阶段(10~30 min)和稳定阶段(30~71 min)。此外入渗开始第一分钟的入渗速率被认为是初始入渗速率,0~71 min 内的平均入渗速率被定义为总的平均入渗速率。
1.5 土壤入渗模型
水分在土壤中的入渗是复杂的物理过程,各国学者在对土壤入渗的探索过程中,建立了许多模型来模拟土壤水分入渗速率随时间的变化规律。根据模型是否具有物理意义分为两类:一类是物理模型,包括Horton[22]模型和Philip 模型[23];另一类是经验模型,包括Kostiakov 模型[24]和Kostiakov-Lewis模型[25]。本研究采用上述四个模型对青藏高原多年冻土区不同生态类型土壤入渗过程进行模拟,对比入渗过程的现场试验对不同植被类型高寒草地上土壤入渗过程模拟的实用性和准确性进行对比。关于上述四个模型的简述如下:
(1)Horton模型:
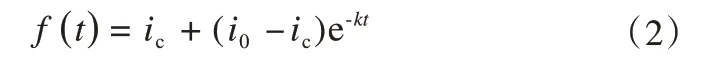
(2)Philip模型:

(3)Kostiakov模型:

(4)Kostiakov-Lewis模型:

式中:f(t)为入渗速率,mm·min-1;t为入渗时间,min;i0为初始入渗率,mm·min-1;ic为稳定入渗率,mm·min-1;S为土壤吸湿率;a、b、a1、b2、n和k为拟合参数。
依据相关系数(r)、均方根误差(RMSE)、决定系数(R2)来进行评估模型模拟的适用性。
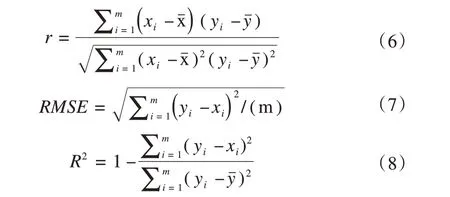
式中:y和x分别为模拟值和实测值;和分别为模拟值和实测值的平均值;i为第i个实测数据;m为实测数据总数。
1.6 统计分析
本研究中采用单因素方差分析(ANOVA)对土壤理化特性以及植被根系生物量的变化差异进行分析,显著差异在0.05水平上确定。采用相关性分析方法分析土壤理化特性以及植被根系生物量与不同入渗阶段入渗速率的相关关系。泰勒图以及入渗曲线拟合通过MATLAB 软件实现,其他图由Origin软件绘制。
2 研究结果
2.1 不同植被类型条件下土壤入渗特征
野外原位实验中,在多年冻土区,不同植被类型的活动层土壤入渗过程随时间的变化如图2 所示。实验结果显示,不同植被类型土壤入渗过程都有相同的变化趋势,随着入渗时间的增加,入渗速率均减小并趋于稳定。但是,高寒草原与裸地入渗速率减小梯度相对于高寒草原和高寒沼泽草甸更大。而且仅通过初始入渗速率和稳定入渗速率来研究土壤的入渗能力不能准确反映土壤理化特性以及植被特征差异对入渗过程的影响。所以对入渗过程划分成不同的入渗阶段,进而分析不同植被类型的土壤入渗过程差异。
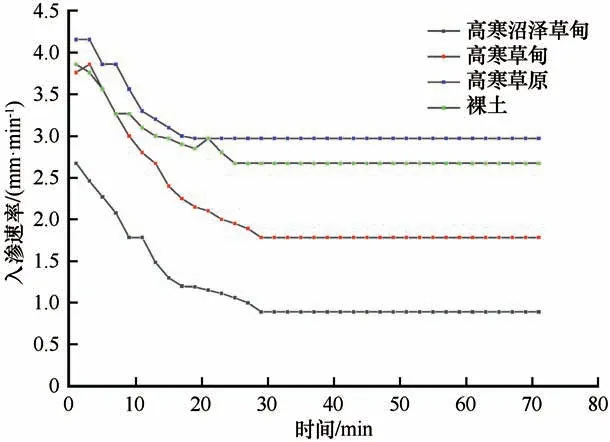
图2 不同植被类型土壤入渗速率随时间的变化Fig.2 Variation of soil infiltration rate with time for different vegetation types
本研究中,选择活动层土壤的初始入渗速率(第1 min 的入渗速率)、瞬变入渗速率(0~15 min 快速减小阶段)、渐变入渗速率(15~30 min 缓慢减小阶段)、稳定入渗速率(30~71 min 趋于稳定阶段)以及平均入渗速率(0~71 min 的平均值)作为分析土壤入渗能力的指标。如图3 所示,不同植被类型的土壤初始入渗速率、渐变入渗速率、稳定入渗速率的大小顺序均为:高寒草原>裸地>高寒草甸>高寒沼泽草甸。相对于裸土,高寒草原草地的初始、渐变、稳定、以及平均入渗速率分别升高了6.7%、7.9%、6.3%、5.1%,而高寒草甸草地分别降低了2.6%、22.0%、29.1%、20.0%,高寒沼泽草甸草地下降了30.8%、47.0%、64.5%、52.3%。在瞬变阶段,高寒草甸的土壤入渗速率几乎超过了裸地土壤,较裸地土壤升高了0.6%。
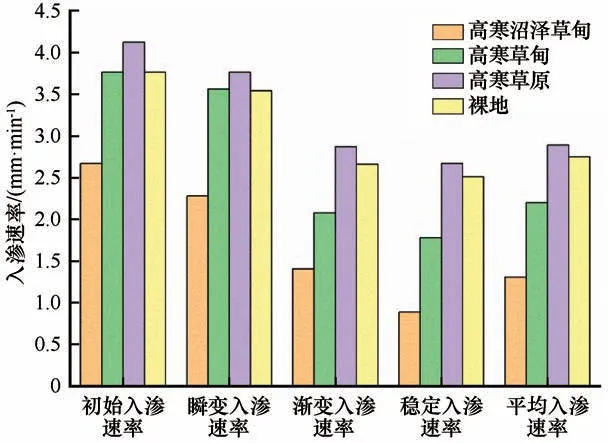
图3 不同植被类型土壤入渗能力指标Fig.3 soil infiltration capacity index of different vegetation type
2.2 土壤入渗速率影响因子分析
土壤是多孔介质,当水分在土壤中运移时,入渗速率必然会受到初始含水量、土壤质地、有机质含量、水稳性团聚体含量等土壤理化特性的影响。土壤理化特性对入渗过程的影响机制分析是不同植被类型土壤入渗性能差异研究的关键。由表1可知,青藏高原多年冻土区活动层土壤干密度和砂粒含量与不同入渗阶段的入渗速率均呈正相关关系。土壤初始含水量、根系生物量、有机质含量、粉粒和黏粒含量与不同入渗阶段的入渗速率均呈负相关关系。而且有机质含量与初始入渗速率、瞬变入渗速率、平均入渗速率呈显著的负相关关系,黏粒含量与稳定入渗速率呈显著的负相关关系。
2.3 入渗模型评估
本研究中采用相关系数、均方根误差、决定系数、标准差等统计参数以及各个入渗公式中的经验参数来评估4种入渗模型在多年冻土区活动层土壤入渗过程模拟中的适用性。根据不同入渗模型模拟的统计参数可知(图4,表2),四个入渗模型均能较好地模拟不同植被类型草地活动层土壤的入渗过程,其相关系数在0.85~1.00 之间,决定系数≥0.80。其中,Horton 模型在不同样地的实验点均具有最高的相关系数(>0.95)、决定系数(≥0.97)和最小的均方根误差值(图4,表2),说明Horton 模型最适用于模拟多年冻土区活动层土壤的入渗过程。而Philip 模型在模拟多年冻土区活动层土壤入渗过程的适用性较差[相关系数(<0.90)和决定系数(≤0.85)最低,均方根误差值最大](图4,表2)。
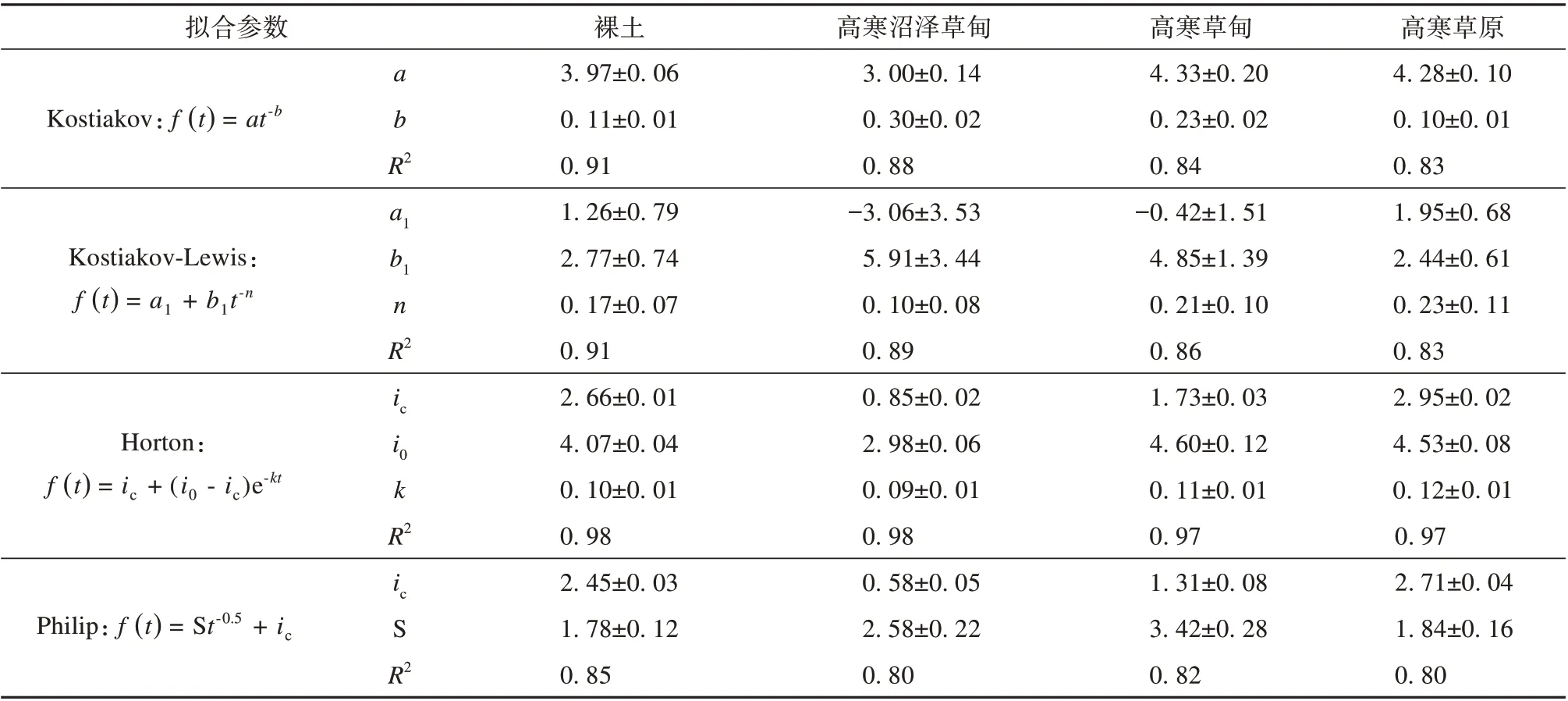
表2 不同植被类型土壤入渗模型的拟合参数Table 2 Fitting parameters of soil infiltration models for different vegetation types
根据不同入渗模型中的经验参数可知,Kostiakov 模型中参数a表示初始渗透率,不同植被类型土壤的a值介于3.00与4.33之间,与实测初始入渗率较为接近;参数b表示渗透速率随时间的减小梯度,不同植被类型土壤入渗的模拟值大小顺序为高寒草原>裸土>高寒草甸>高寒沼泽草甸,与不同植被类型条件下的实测入渗速率变化情况基本相同(表2)。Kostiakov-Lewis 模型中,a1值表示稳定入渗速率,b1值与初始入渗速率有正相关关系,n值表示初始入渗率到稳定入渗率的减小梯度。模拟结果中a1值介于-3.06 和1.95 之间,而且在高寒沼泽草甸和高寒草甸试验点,a1为负值,这与实际下渗理论不相符,在高寒草原和裸地试验点,a1与实测值差异较小。在不同试验点,n值大小顺序为高寒草原>高寒草甸>裸地>高寒沼泽草甸,与瞬变阶段土壤入渗率的大小一致(表2 和图2),而与其他下渗时段的下渗率不同,说明n与瞬变阶段的入渗率有关。Horton 模型对初始、稳定入渗速率均可以较好地模拟,模拟结果显示i0介于2.98和4.60 之间,ic介于0.85 和2.95 之间,在不同试验点的拟合值均接近实测值(图5)。Philip 模型中,不同样点稳定入渗率的实验值介于0.58 和2.71 之间,均表现出较好的拟合效果(表2)。另外,不同入渗模型在裸地的模拟效果均最优(表2)。另外,在试验过程中,还发现土壤入渗速率逐渐减小接近于稳定入渗率的过程中出现了快速降低的突然变化,而且模型的模拟曲线均偏离这一突变过程的实测值。说明这些模型均无法准确模拟下渗过程的这一突变情况(图5),这说明下渗在非饱和向饱和土壤过渡的过程中,由于土壤水分运移性质的突然变化引起了土壤入渗速率的不连续变化,导致土壤入渗模型难以准确模拟这一现象。
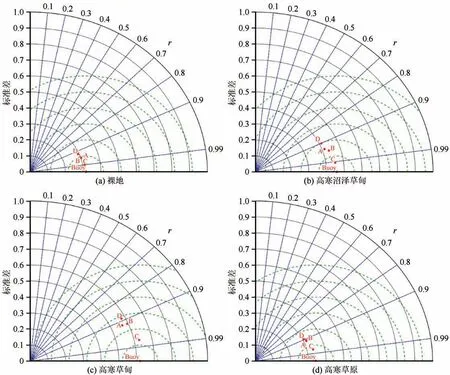
图5 不同入渗模型的模拟值与实测值之间关系的泰勒图Fig.5 Taylor program for infiltration rates among infiltration models with observations[bare land(a),alpine wet meadow(b),alpine meadow(c),and alpine grassland(d)]
综上所述,Horton 模型最适用于描述多年冻土区活动层土壤水分的入渗过程。尽管Philip 模型由统计参数得出的模拟性能较差,但较Kostiakov 和Kostiakov-Lewis 模型更符合实际下渗理论。虽然这些入渗模型可以模拟不同植被类型土壤的入渗过程,但它们均不能模拟入渗过程中的突变情况,而且不同入渗模型对裸地的模拟情况均优于其他植被类型(图4~5)。
3 讨论
3.1 土壤入渗能力影响因子分析
土壤入渗能力一般受土壤物理性质、土壤水力、热力性质和植被特征影响[26]。活动层土壤的地下生物量、土壤含水量、土壤有机质、土壤总孔隙度和土壤团聚体等土壤属性,均是影响土壤入渗能力的主要因素[2,27]。通过原位试验和模型的模拟研究发现,青藏高原多年冻土区活动层土壤的初始含水量、根系生物量、有机质含量、粉粒和黏粒含量与不同入渗阶段的入渗速率均呈负相关关系,土壤干密度以及砂粒含量与不同入渗阶段的入渗速率呈正相关关系(表1)。这些研究结果显示,在青藏高原多年冻土区,土壤的基本理化性质、生物性质及土壤结构和质地对土壤入渗的各个阶段都有强烈的影响。其中,土壤初始含水量以及土壤质地与土壤入渗速率的关系与其他学者的研究结果一致[28-29]。而土壤干密度与根系生物量以及有机质含量与土壤入渗速率的相关关系与前人的研究相悖[20,30-31],这主要是由于本研究样地选取了青藏高原多年冻土区高寒沼泽草甸和高寒草甸,该植被生态系统的根系呈草毡层,其根系致密,有机质丰富,土壤干密度偏小,土壤对水分入渗具有强烈的阻滞作用,土壤持水能力相对较高;而高寒草原的根系稀疏,土壤通透性良好,对入渗的阻滞作用相对较弱,土壤持水能力相对较差[13]。因此,高寒草甸和高寒沼泽草甸初始含水量显著高于高寒草原和裸地[图6(a)]。土壤有机质是通过促进团聚体的形成而间接影响土壤的入渗性能[32],而土壤干密度是土壤紧实度和气相比的间接反映,决定着土壤内部结构的松散程度[33]。当土壤含水量较高时,毛管孔隙被水分填充,活动层土壤水分的水力梯度减小,从而降低了水分入渗的驱动力,表现出较小的初始土壤入渗速率[34]。因此,高寒植被根系和土壤初始含水量强烈影响多年冻土区活动层土壤的入渗能力。不同高寒植被类型根系生物量差异显著[图6(b)],在0~30 cm 深处,高寒沼泽草甸、高寒草甸根系生物量显著高于高寒草原,而根系生物量与土壤入渗速率呈负相关关系(表1),这也表明根系对土壤水分的入渗具有阻挡作用。而且当土壤入渗过程达到稳定时,稳定入渗速率的大小顺序为:高寒草原>裸地>高寒草甸>高寒沼泽草甸,而高寒沼泽草甸以及高寒草甸浅层有机质含量显著高于裸地和高寒草原,容重显著低于高寒草原和裸地。这充分表明高寒沼泽草甸、高寒草甸浅层致密的根系降低了土壤入渗性能,而高寒草原根系的阻水作用较弱,土壤水分向深层入渗的速率较大。
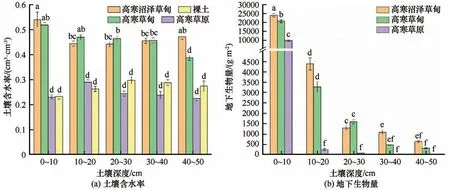
图6 不同植被类型土壤含水量和根系生物量分布特征Fig.6 Distribution of root biomass and soil water content in different alpine ecosystems
受气候变暖的影响,青藏高原多年冻土区活动层土壤厚度明显增加,冻结开始时间明显推迟,融化开始时间明显提前,一年中,活动层土壤冻融过程的频率明显增加[35-37],改变强烈。活动层土壤的这些变化强烈地改变了土壤的内部结构、根系分布、空隙大小及其分布、土壤干密度和有机质含量的变化[38],不断影响土壤水分的入渗、蒸散发、产汇流等过程,改变了土壤水文循环过程及水资源的空间分布。而且高寒生态系统由高寒草甸不断向高寒草原退化[39]。因此,在未来气候持续变暖的情况下,高寒生态系统将不断发生变化,强烈改变多年冻土区土壤的水文过程和高寒陆地水文循环过程。高寒区不同植被类型草地土壤入渗过程的研究为未来气候变暖下土壤水文过程的机理研究提供理论依据。
3.2 不同入渗模型的适用性分析
高寒植被变化与冻土活动层土壤变化相互影响,气候变化与冻土变化的协同作用是青藏高原植被退化和土壤水文过程变化的主要驱动力[40]。在当前青藏高原气温逐渐升高,冻土退化的大背景下,研究多年冻土区活动层土壤水文过程,对认识不同植被类型条件下的产流机制、进行冻土环境保护、研究高寒生态系统对全球气候变化的反映和评估其恢复的潜力等至关重要[14,41]。土壤入渗是活动层土壤水文过程的关键过程之一,准确分析和预测不同植被类型条件下,活动层土壤入渗过程是研究多年冻土区活动层土壤水文过程和水资源时空变化的主要内容。通过对比不同入渗模型对多年冻土区活动层土壤入渗过程的模拟,发现Horton 模型对多年冻土区不同植被类型土壤入渗过程的模拟均优于其他三个入渗模型。这与Wang 等[42]对高寒草甸入渗过程的模拟结果以及Hu 等[43]对青藏高原多年冻土区不同下垫面土壤入渗过程模拟结果一致。尽管Philip 模型基于RMSE、R2、相关系数等统计参数得出的模拟性能较差,但相对于Kostiakov模型以及Kostiakov-Lewis 模型对高寒沼泽草甸、高寒草甸入渗过程的模拟,Philip 模型模拟的经验参数更符合实际下渗理论。虽然Hu 等[43]也发现Philip模型基于RMSE、R2等统计参数得出的模拟性能较差,但他们未对模型中的参数与实际下渗过程进行比较。不同入渗模型对高寒裸地土壤入渗过程的模拟均优于其他植被类型草地(表2,图4),说明植被下垫面条件对土壤入渗过程估算有一定的影响。Wilk 等[44]在分析土壤耕作和植被覆盖对土壤水分入渗的影响时,也发现类似的现象,这可能是因为这些渗透模型主要是针对裸地建立的[45],在初次构建模型时没有考虑到植被条件的影响。
尽管这些入渗模型能够较好地模拟不同植被类型土壤的入渗过程,但对入渗过程中突变过程的模拟较差(图5)。而且对质地均一的土壤的入渗过程模拟相对较好,而对不同植被类型土壤水分入渗过程的模拟较差,而且由于不同植被类型土壤其复杂的土壤结构、有机质含量、根系分布及其生物量、孔隙度的不同其下渗过程的模拟效果也有所不同。因此,通过野外现场实验结合土壤入渗模型对条件艰苦、对气候变化敏感脆弱的多年冻土区入渗过程的适用性研究是冻土区入渗模型进行改进和高寒区陆地水文过程模型改进具的重要理论基础。
4 结论
通过双环入渗仪对青藏高原多年冻土区高寒沼泽草甸、高寒草甸、高寒草原、裸地土壤入渗过程进行野外现场实验,分析了土壤有机质、容重、初始含水量、根系生物量以及土壤质地对活动层土壤入渗能力的影响,针对四种常用的入渗模型对多年冻土区活动层土壤入渗模型的适用性进行了分析研究,主要结论如下:
(1)多年冻土区活动层土壤的初始、渐变、稳定和平均下渗率的大小顺序为高寒草原>裸土>高寒草甸>高寒沼泽草甸,而在瞬变入渗阶段高寒草甸土壤的入渗速率几乎超过了裸土,而且这一变化与Kostiakov-Lewis模型中的参数n有关。
(2)多年冻土区植被根系强烈影响活动层土壤的入渗性能。高寒沼泽草甸和高寒草甸致密的毛毡层根系可能堵塞土壤孔隙,降低土壤入渗能力,而高寒草原根系则对土壤水分入渗的阻挡作用较弱。
(3)通过四种常用的入渗模型对高寒土壤入渗过程模拟发现,Horton 模型对不同植被类型土壤水分入渗过程的模拟均优于其他三种入渗模型。这些入渗模型均能较好地模拟不同植被类型活动层土壤的入渗过程,但对入渗过程中突变情况的模拟能力较弱。而且植被类型对入渗模型的模拟精度存在负面影响。
