背景噪声对清醒小鼠初级听皮层神经元听觉响应特征的影响
2021-12-16宋长宝
宋长宝,赵 岩,柏 林
南方医科大学1生物医学工程学院数学物理系,2基础医学院生理学教研室,广东 广州 510515
自然界声音信号常伴随着无序多变的背景噪声,如何从复杂的声环境中甄别具有生物意义的信息并对其加以处理、利用是听觉感知的重要任务之一。声环境中存在的背景噪声是如何影响我们的听觉感知的?听觉系统如何保持对目标声信号的听觉表征?对人类的听觉任务研究表明,连续的背景噪声会降低人类听觉行为测试的辨别能力[1-5],长时间高强度的噪音会损伤听觉感知,严重时甚至会导致短暂或永久性听力损失[6,7]。另一些研究则表明[8-12],某些背景噪声能够提高听觉系统的信号检测能力。因此,背景噪声对听觉感知的影响是双向的,信号源和噪声强度的匹配程度可能是影响因素之一。
既往的神经电生理学实验认为,高强度噪声对听觉中枢神经元的目标声反应往往表现为抑制,如阈值上移、动作电位数减少或电位幅度减小等[13-17]。我们前期在麻醉大鼠初级听皮层(A1)上进行的研究提出线性阈值平移模型,噪声背景相当于降低了声刺激的有效强度,听觉神经元对声音基本信息的表征是通过声强阈值的线性平移来进行,其平移值与背景噪声水平相关[14]。即使在听觉中枢的核心区域内,对噪声环境中的信号处理也具有显著的多样性。随着信噪比(SNR)的降低,噪声的加入抑制了松鼠猴听皮层某些核心部位对于调频声(FM)的反应,但听皮层的另一些区域对前景声刺激的表征仍然保持稳定,甚至在特定SNR时显著增强。研究者认为这些听皮层区存在特定的神经元群,在背景噪声中处理前景信号的方式不同,可能有助于复杂声学环境中的信号表征和辨别[18]。另有学者报道听皮层神经元采用差别增益控制模型来保持其对新刺激的有效应答,即听觉神经元能根据噪声环境中新旧刺激的差别,主动调节自身增益,以获取更好的敏感性[19-21]。
上述对于听觉中枢的电生理学研究[13-17,19-21]多以麻醉动物为研究对象,且多为多单位细胞外记录[13,15-20],或采取多单位记录后经数学拟合的模式进行分析[19]。随着多导电极埋置技术的发展,近年来在清醒动物听觉中枢进行的多单位记录时有报道,如Christensen等[11]对清醒小鼠听皮层、Malon等[18]对清醒松鼠猴听皮层的研究。对清醒动物进行在体单神经元记录难度大,少有研究采用单细胞贴附式记录探查噪声背景下清醒动物听觉中枢神经元的声信号处理特征。此外,噪声环境中听觉时间反应特性方面的研究较多集中于以人为实验对象的听觉诱发电位的延时变化[22,23],在听皮层的多单位记录证实神经元群体的听反应延时随噪声强度增加而延长[24],关于背景噪声对听觉中枢单神经元时间调谐的影响尚未见报道。本研究拟以清醒活动小鼠为主体,采用细胞贴附式记录技术,通过记录电极与A1单神经元之间形成松散封接,设计多样化声刺激序列,从声强调谐、频率调谐和时间调谐特性3方面分析背景噪声对清醒动物A1单神经元听觉响应的影响,以期更全面地了解清醒状态下听觉中枢对噪声背景中目标声的处理方式,为复杂声学环境中听觉感知的中枢机制提供理论基础。
1 材料和方法
1.1 实验动物
实验在6~8周的清醒状态C57BL/6小鼠上进行。所有的动物实验手术程序都遵循南方医科大学动物使用委员会标准。
1.2 手术准备
实验前,动物用戊巴比妥钠麻醉,并使用温控系统使动物体温维持在37 ℃,用立体定位仪固定,保持耳朵自由。麻醉完全后用牙科水泥将一个长约1.5 cm的平头金属钉粘合在颅骨前部正中位置,再将小鼠固定到屏蔽室的实验平台上,在立体显微镜下小鼠右侧初级听皮层经手术开颅(直径0.5~2 mm)暴露,同侧(右)耳道堵塞。在手术和记录过程中,暴露的皮质由恒温人工脑脊液(ASCF;单位mmol/L:NaCl,132 mmol/L;NaHCO3,20 mmol/L;ΚCl,2.5 mmol/L;NaH2PO4,1.2 mmol/L;MgSO4,1.1 mmol/L;CaCl2,2 mmol/L;Hepes,3 mmol/L;葡萄糖15 mmol/L)保持湿润。
1.3 声音刺激
声音波形由自编的Labview 程序合成,通过NI数据采集卡输出(PCI-4052E,NI),采用自由场喇叭(ES1)发声。喇叭被放置在小鼠左侧耳10 cm处。根据实验目的,设置5种声刺激序列。
1.3.1 频率-声强序列 所有神经元以短纯音序列(频率2~64 kHz,1/10 octave递增;0~70 dB声压级,10 dB递增;稳态时长50 ms)扫描5遍以获取单神经元的声音感受野(TRF),初步确定该神经元的特征频率(CF)和最低阈值(mTh)。
1.3.2 持续宽带噪声序列 所有神经元给予持续宽带白噪声(频率范围1~32 kHz;0~48 dB声压级,12 dB递增;时长10 s),重复5遍,检测单神经元对不同强度持续噪声的反应情况。
1.3.3 声强-噪声强度序列 短纯音(频率为CF;0~90 dB声压级,10 dB递增;稳态时长50 ms)与不同强度背景噪声(频率范围1~32 kHz;0~48 dB声压级,12 dB递增,时长50 ms)经TDT 3混合,重复5遍。
1.3.4 频率-噪声强度序列 短纯音(频率2~64 kHz,1/10 octave递增;60 dB声压级;稳态时长50 ms)与不同强度背景噪声(同1.3.3)经TDT 3混合,重复5遍。
1.3.5 延时-噪声强度序列 短纯音(频率为CF,声强为mTh+20 dB声压级,稳态时长50 ms)与不同强度背景噪声(同1.3.3)经TDT 3混合,重复20遍。
1.4 电生理记录
电声理实验在听觉屏蔽室中进行。采用细胞外贴附式记录法,记录皮层下400~700 μm(听觉皮层第四层和第五层浅表)的神经元,记录用电极(P-97拉制,尖端直径1~2 μm,电阻1~9 MΩ),电极内液为ACSF(同1.2)。记录信号(采样率为20 kHz,滤波2 kHz)经过200B放大器(Axon Instruments)放大及信号转换,通过MATLAB软件提取数据。
1.5 数据分析
通过自写Matlab程序读取每个声刺激下响应的动作电位发放数、延时及波形等数据。采用Matlab、OriginPro及Excel对数据进行分析和作图。
1.6 统计学分析
所有记录到的神经元需完成1.3.1和1.3.2声序列扫描才纳入后续记录。由于清醒动物在体单神经元记录难以长时间维持,根据每个神经元的声强响应特征和后续3种声序列扫描完成情况,将神经元分为4组:单调性-声强组20例,非单调性-声强组6例,单调性-频率组25例,单调性-延时组15例。
采用SPSS 19.0软件进行统计分析,正态分布计量资料资料以均数±标准差表示,不同强度噪声组和0 dB组比较采用配对样本t检验,两组间比较采用独立样本t检验,观察两组计量资料的相关性采用Pearson相关性分析,并进行线性回归,求得相关系数、斜率和R2值。P<0.05表示差异具有统计学意义。
2 结果
2.1 基本情况
采用细胞贴附式记录技术共记录清醒小鼠A1神经元44例,封接电阻平均为45±13.5 MΩ。记录位置为皮层下400~700 μm,相当于小鼠听觉皮层第四层和第五层浅表,根据动作电位波形特点(动作电位峰/谷值小于1,且峰-谷时间>0.4 ms),这44例神经元均为锥体神经元。根据神经元对声强的反应特点,将兴奋性神经元分为单调性和非单调性两类:单调性神经元的放电率随声强增大而增加;非单调性神经元在某一声强时放电率达到最大,当声强超过一定限度时,神经元放电率反而逐渐减小。44例神经元均进行1.3.1声刺激序列扫描和1.3.2持续噪声扫描,以获取单个神经元的基本声响应特征以及对持续背景噪声的响应特点。单调性神经元38例,其中20例神经元完成了1.3.3声序列扫描,25例神经元完成了1.3.4 声序列扫描,15 例神经元完成了1.3.5声序列扫描;非单调性神经元6例,完成了1.3.3声序列扫描。
2.2 持续噪声下清醒小鼠A1神经元的背景发放率
结果显示A1听神经元仅在给予噪声刺激后10~40 ms内出现瞬态的尖峰响应(图1A),不同强度的持续噪声对44例神经元的发放率无影响(图1B,C)。

图1 A1神经元对连续噪声刺激的背景发放率Fig.1 Background firing rate of A1 neurons in response to continuous noise stimuli.A:The firing PSTH diagram of the sample neuron to continuous noise shows that A1 neurons only have transient spike response within 50 ms after noise stimulation onset (5 repeats).B:Background firing rate (the number of action potentials/time within 10-40 ms after the noise onset)of the sample neuron under continuous noise of different intensities.C:Based on the firing rate of neurons under 0 dB noise intensity,the firing rates of 44 neurons to different noise intensities are normalized.The gray line represents a single neuron and the red line represents Mean±SD(the same below).The background noise intensity has no significant effect on the firing rate of neurons(P=0.12,12 dB vs 0 dB;P=0.09,24 dB vs 0 dB;P=0.08,36 dB vs 0 dB;P=0.12,48 dB vs 0 dB).
2.3 噪声背景对清醒小鼠A1神经元声强调谐特征的影响
图2A、3A分别为1例单调性神经元和1例非单调性神经元的声强调谐范围和发放强度变化的示例图。不管神经元的声强调谐是否表现为单调性,PSTH和伪彩图均显示:当背景噪声增加到一定强度时,随噪声加大,神经元对目标声的探测阈值增高,声强响应范围缩小,发放率减小(图2A~C、3A~C)。对阈值改变10%以上的点进行线性回归,求出神经元的声强阈值随背景噪声变化的斜率(slope),R2值和P值。图2D、3D分别显示20例单调性神经元和6例非单调性神经元的阈值变化趋势,所有细胞的R2值均>0.90(P<0.05),表明阈值变化与背景噪声强度改变有良好的线性关系。我们用阈值线性变化的斜率来表示噪声水平对神经元声强响应的影响程度,图3F显示,神经元群体斜率的差异较大,而单调性和非单调性神经元之间的阈值变化斜率未表现出显著性差异。我们还观察了非单调性神经元的阈值变化和最佳声强(指单一频率纯音刺激时,引起神经元发放率最高的声强)变化之间的关系。图3E显示6例非单调性神经元最佳声强随背景噪声增强也逐渐增大,同样经线性拟合获得最佳声强随背景噪声变化的斜率。以同一神经元阈值变化斜率为横坐标,最佳声强变化斜率为纵坐标做散点图并进行线性拟合,结果显示:背景噪声水平改变时,A1非单调性神经元最佳声强的变化趋势与声强响应阈值的变化趋势呈正相关(图3G),拟合系数略小于1。

图2 不同背景噪音条件下单调性神经元声强调谐特征的变化Fig.2 Intensity tuning of monotonic neurons for tones in different background noises.A:Effects of different noise levels on intensity response range and firing rate of the example monotonic neuron.B:The threshold of the sample neuron to target stimulation increases with the increase of background noise intensity.C:The firing rate of the sample neuron to target acoustic stimulation decreases as the background noise increases.D:Effect of background noise levels on threshold of sound intensity response of 20 monotonic neurons (paired-sample t-test,*P=0.03,**P=0.006,***P<0.001, vs 0 dB).E:Effect of background noise on normalized firing rate of monotonic neuronal population(paired-sample t-test,P=0.17,12 dB vs 0 dB;P=0.06,24 dB vs 0 dB;***P<0.001,vs 0 dB).
2.4 背景噪声对清醒小鼠A1神经元频率调谐特征的影响
图4A~D为1例神经元的频率调谐范围、最佳频率(BF,指单一强度纯音刺激时,引起神经元发放最强的频率)、频率响应带宽(BW)和发放强度变化的示例图。当背景噪声增加到一定强度时,随噪声强度加大,神经元对目标声的频率响应范围缩小(图4A、C),平均发放率减少(图4D),而最佳频率BF基本不变(图4B)。BF近似于特征频率CF,可表征听觉神经元的频率选择性,以同一神经元在0dB噪声环境中的BF为横坐标,其他噪声强度下的BF为纵坐标,做散点图并进行线性回归(图3E),斜率为0.984,R2=0.986(P<0.001),与我们前期在麻醉动物上进行的实验结果相似,噪声强度并不影响清醒小鼠A1神经元对短纯音响应的频率选择性。将不同噪声强度下神经元频率响应带宽BW及平均发放率按照0dB噪声背景下的数值进行归一化处理,显示BW和平均发放率随噪声强度增大而逐渐减小。在不同强度的噪声背景中,A1听觉神经元表现出目标声频率响应范围的缩小和响应强度的降低,而神经元的频率选择性保持恒定。
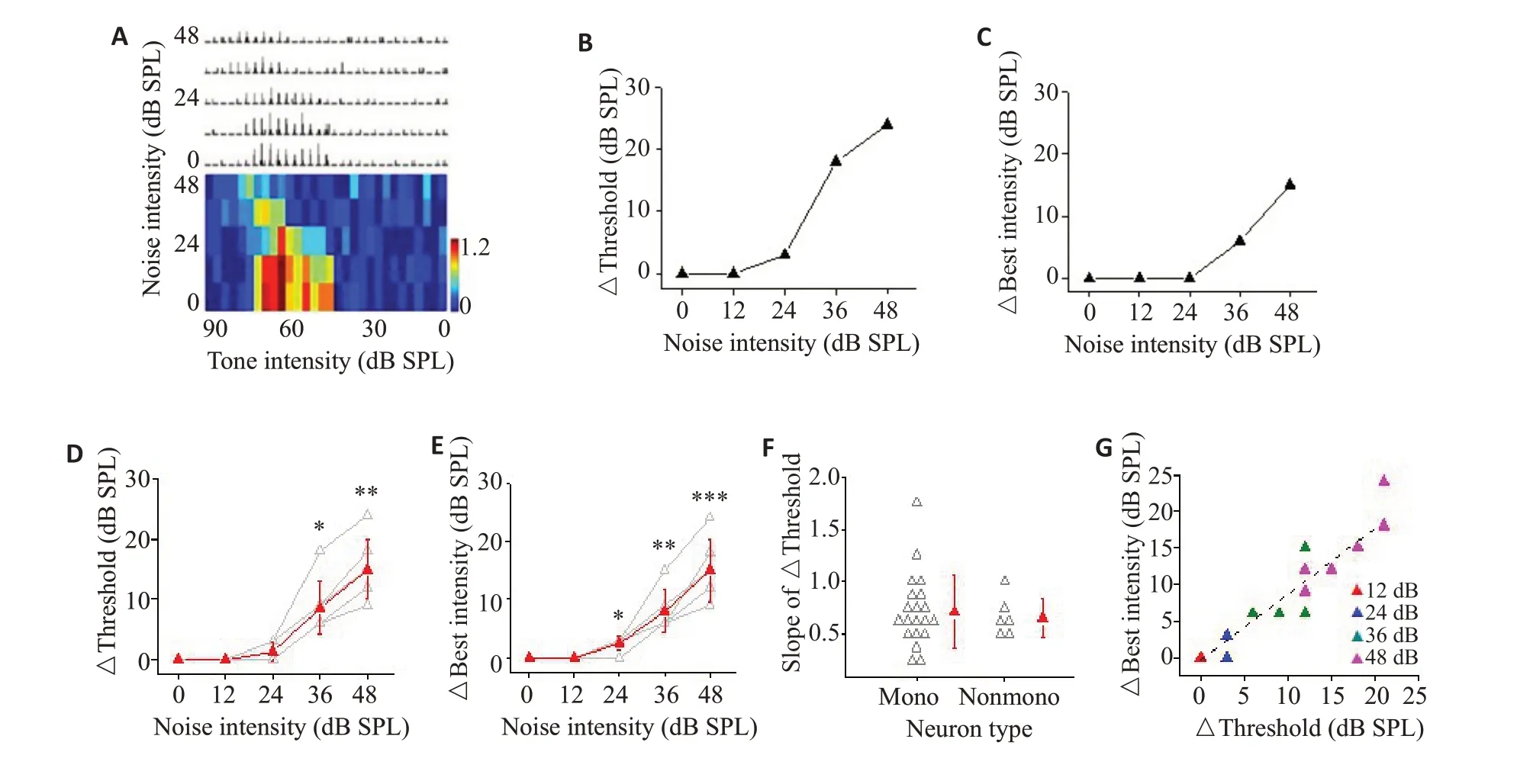
图3 不同背景噪音条件下非单调性神经元声强调谐特征的变化Fig.3 Intensity tuning of nonmonotonic neuron for tones in different background noises.A:Effect of background noise level on intensity response range and firing rate of the example nonmonotonic neuron.B:The threshold of the sample neuron to target acoustic stimulation increases with the increase of background noise level.C:The best intensity of the neuron increases along with the elevation of the background noise.D:Effect of noise level on the threshold of the intensity response of 6 non-monotonic neurons(P=0.76,24 dB vs 0 dB;*P=0.005,**P=0.001,vs 0 dB).E:Effect of noise level on the best intensity of nonmonotonic neurons (paired-sample t-test,*P=0.004,**P=0.003,***P=0.001, vs 0 dB).F:There was no significant difference in the slope distribution of threshold changes between monotonic and non-monotonic neurons (independent-sample t-test, P=0.70).G:The best intensity of nonmonotonic neurons shows a positive correlation with the change of threshold (correlation coefficient 0.944,slope=0.899,R2=0.890,P<0.001).

图4 不同强度噪音背景下神经元频率调谐特征的变化Fig.4 Frequency tuning of A1 neurons for tones in different background noises.A:Influence of background noise level on the frequency response bandwidth and firing intensity of the sample neuron.B:The noise intensity has no significant effect on the BF of the sample neuron.C:The BW of frequency tuning of the example cell decreases with the increase of background noise intensity.D:The firing rate decreases along with the enhancement of background noise.E:The noise level has no significant effect on the BF of 25 neurons(slope=0.984,R2=0.986,P<0.001).F:An increased background noise compresses the normalized frequency bandwidth of the neuron (*P<0.001, vs 0 dB).G:Normalized firing rate of the neuron group decreases with the increase of noise level(P=0.18,12 dB vs 0 dB;*P<0.001,vs 0 dB).
2.5 背景噪声对A1神经元听觉反应时间及精确性的影响
图5A为示例神经元在不同背景噪声强度下响应的动作电位散点图。随着背景噪声强度的增加,神经元发生响应的FSL也随之增大(图5B),15例神经元表现出同样的变化趋势(图5D)。我们又进一步对FSL的Jitter值(表征神经元对同一声刺激发生响应的时间精确性)进行统计分析,发现在不同背景噪声条件下,FSL的Jitter值基本不变。

图5 不同背景噪音条件下听觉响应延时及精确性的变化Fig.5 First spike latency and jitter of A1 neurons for tones in different background noises.A:The response scatter of the sample neuron to the target tone under different noise background.The black dot represents the first action potential.B:The FSL of the sample neuron increases with the enhancement of background noise.C:The jitter of FSL showed no significant change along with the elevation of background noise intensity.D:The FSL of 15 cases to target stimulation are prolonged with the increase of background noise(*P=0.001,**P=0.002,***P<0.001,vs 0 dB).E:The background noise intensity has no significant effects on the jitter of group(paired-sample t-test,P=0.58,P=0.86,P=0.06,P=0.32,vs 0 dB).
3 讨论
在噪声环境中,低强度的信号很难被检测到的,尤其是当噪声的强度接近我们辨识的阈值时,这是心理声学中的掩蔽现象[3,25,26]。早期研究描述了听觉通路各中枢神经元在背景噪声下的电生理特性,这些研究大多是采用多单位记录技术进行神经元群体水平上的研究。Rabinowitz等[19]通过对噪声环境中麻醉猫听皮层神经元的多单位记录结果进行数学拟合,提出差别增益控制模型,该模型并未实际检测不同噪声背景中单神经元对目标声反应特性的变化,因此缺乏直接证据证明听觉中枢神经元确实存在主动的增益调节。我们在前期实验中,采用贴附式记录来探究背景噪声对麻醉大鼠初听皮层单神经元声响应特征的影响,提出阈值平移模型[14]:听觉神经元对某种包含特定意义(如频率信息)的声信号的响应是其固有特性,背景噪声的加入改变了目标声信号的有效物理强度,使神经元对声信号的反应阈值增大,相应的感受野整体上调,但神经元不改变其特定的功能选择性,对阈值上相等声强的目标声,其反应特性如频响带宽、反应强度等也保持相应水平,提示噪声背景中的听觉中枢单神经元并没有出现主动的增益调节。本研究旨在前期麻醉动物结论的基础上探查清醒小鼠听皮层单神经元在持续噪声环境中听觉响应的频率、声强和时间特征,探讨复杂声环境中听觉中枢神经元对声信号表征的基本工作机制,是否存在主动的增益调节,为听觉系统的信号识别机制提供理论证据。
受清醒动物在体单神经元记录时长所限,本实验设置的宽带白噪声集中于中低强度(0~48 dB),是人群日常生活和工作环境中的常见噪声强度。结果表明,持续噪声背景中清醒动物听皮层单神经元处理目标声的方式与麻醉动物类似,较小的噪声强度不引起阈值改变或只产生极小变化;背景噪声超过某一水平后,阈值偏移与噪声强度间呈高度线性相关。这种变化趋势同样表现在A1非单调性神经元,其阈值也随噪声强度增大表现出线性升高。阈值随噪声强度变化的趋势呈现出神经元之间的差异性,但A1单调性和非单调性神经元阈值变化的斜率分布未见明显差异,表明背景噪声强度对听觉神经元声强调谐特征的影响是一致的,都是基于噪声背景中目标声有效强度的变化而产生相应改变。早期研究发现[27],噪声强度每增加1 dB,豚鼠下丘中央核神经元的声强反应范围就向高强度方向偏移0.97 dB,且这种变化趋势与神经元种类(单调或非单调)无关。猫下丘中央核和初级听皮层的研究也证实了这种线性变化规律[15-17,28]。因此,噪声背景下听觉神经元的声强反应特征在阈值增大的基础上发生线性推移可能是整个听觉系统的基本处理方式。不同的是,低位听觉中枢如下丘神经元,受背景噪声影响的声强反应区的推移幅度可能直接反映噪声对目标声强度的削减效应(斜率接近1),而A1神经元的阈值和声强反应区的偏移系数则表现出更大的差异化(斜率0.25~1.75),这可能归因于听皮层内复杂的环路联系,尤其是抑制性纤维的广泛分布,使得神经元的突触后总和出现明显的差异。Teschner等[24]在麻醉大鼠听皮层上进行的多单位记录显示,高强度噪声(40~85 dB)表现出对神经元反应更大程度的压抑,因此背景噪声对神经元群体反应的影响无法用单一的阈值平移模型进行拟合。对于噪声环境中清醒动物听觉中枢单神经元的声强调谐特征,还需要设置更宽泛的噪声强度进行进一步探查。
另一方面,非单调神经元对声强的调谐可能出现改变。虽然非单调性神经元的最佳声强随着阈值变化也发生相应的增大,其最佳感受区向高声强方向偏移,但最佳声强的偏移斜率略小于阈值的偏移幅度,意味着非单调神经元的工作范围被压缩,影响听觉系统对目标声低声强部分的编码,造成低声强信号的感觉缺失。当背景噪声强度过高时,非单调神经元工作范围的偏移过大,可能使非单调神经元由闭合型转变为单调性响应的Ⅴ型感受野,即改变了该类神经元的声强调谐特征。有研究认为[17],背景噪声的加入可以移动声强选择性神经元的动态响应范围,但基本不改变神经元的声强响应特征,这可能是由于连续噪声对形成非单调性的兴奋性和抑制性输入具有相同程度的影响。而我们在前期实验中发现,背景噪声对兴奋性和抑制性神经元阈值变化斜率的影响可能不一致[14]。因此,有必要采用在体全细胞记录技术探查听觉神经元在噪声背景中对目标声刺激发生响应时的突触前输入,以明确听觉中枢在处理声强信息时的突触整合情况。
听觉神经元对纯音的频率选择性是声反应的基本特征之一。与麻醉动物中的研究相似,背景噪声的增强,强烈压缩了A1听觉神经元的频率响应范围,但并不影响神经元的最佳响应频率。背景噪声的加入,使得神经元的频响范围缩小,从而表现出更尖锐的频率选择性,有利于噪声环境中对特殊频率的辨别,Christensen等[11]在清醒小鼠A1进行多电极细胞外记录的研究也证实了这一点。另一方面,当环境过于嘈杂时,过高的背景噪声强度掩蔽了前景声信号,导致神经元只对围绕最佳响应频率的少数频段有响应,频段之间的叠加减小,可能削弱听觉中枢对某些频率的响应甚至造成缺失,必然会影响噪声环境中的复杂声信号如语言的辨识感知。
动作电位发放时间是听觉中枢编码的可能机制之一。早期研究即已表明[29,30],在单个神经元中,动作电位的时间特性作为信息载体,尤其是第一个动作电位延时FSL,可以忠实地表征声强,表现在神经元对同一个目标声刺激发生反应的延时非常稳定,其标准差(又称为FSL的jitter)往往只有零点几毫秒,变异系数非常小,这意味着更高的声刺激分辨能力。我们的研究认为,动作电位延时可作为听觉中枢神经元通过不同通路处理不同频率声刺激信息时的指标[31]。关于噪声环境中听觉中枢的时间反应特性方面的报道较少,Teschner等研究[24]显示当SNR减小时,听皮层神经元群体的动作电位延时随之增大,但SNR的改变对延时随目标声强度增大而减小的斜率没有显著影响,提示目标信号声强和SNR对于延时来说是相对独立的影响因素。我们的结果证实,固定前景声的频率(CF)及声强(阈上20 dB)不变,随着背景噪声强度的增加,SNR相应减小,清醒小鼠A1单神经元的FSL随之延长,而FSL的jitter值基本保持不变,提示背景噪声增大时,尽管A1神经元的响应时间出现延长,但对于相同目标声刺激,其动作电位发放时间的精确性不发生变化,表明延时的改变是基于背景噪声引起的阈值增大,仍然能忠实地反映阈值偏移后的实际目标声强。
综上,我们通过在体细胞贴附记录探查了清醒小鼠单个听皮层神经元对噪声背景中目标声响应的声强调谐和频率调谐,进一步证实了我们前期在麻醉动物上得到的阈值平移模型,即背景噪声通过削减目标声信号的有效声强,导致神经元声响应阈值增大,在此基础上发生神经元频率-声强感受范围向高声强方向偏移。这种平移使得神经元对于低声强信号失去感知能力,同时神经元频率选择性更尖锐,一方面有利于嘈杂环境中对重要声信号的探查,另一方面会损害中枢对复杂语言中细微变化的辨识。我们还观察了背景噪声下听皮层神经元的时间响应特性,FSL随噪声强度增大而延长,表明听神经元时间特性的变化也是基于声信号有效强度的减小,而听觉反应时间精确程度不会受到背景噪声的影响,这为我们探究复杂环境中听觉信号的识别机制提供了一定的理论基础。
