短叶罗汉松雌雄株叶片形态结构特征的比较研究
2021-12-13杨凤兰邓冬丽招礼军朱栗琼
杨凤兰,邓冬丽,招礼军,朱栗琼*
(1.广西大学 林学院,广西 南宁 530004;2.广西国际商务职业技术学院,广西 南宁 530000)
雌雄异株是自然植物界中常见的一种现象,在植物界30多万种被子植物中就存在着4%~10%的雌雄异株植物[1]。Renner等在对24万种被子植物资源的调查中发现雌雄异株的植物有959个属,共14620种[2]。在长期的演化过程中,雌雄异株植物的生长特性、资源分配、生理生化特征和基因表达等方面存在着一定的性别差异[3]。目前国内有关雌雄异株植物的研究主要集中在基因[4]、生理生化[5-7]和对环境因子的响应差异[8]等方面。研究雌雄异株植物,不仅可在植物早期进行性别鉴定,为生产实践提供技术支持,而且还有利于揭示雌雄异株植物在演化过程中的生存适应策略。
短叶罗汉松(Podocarpusmacrophyllusvar.maki)为罗汉松科(Podocarpaceae)罗汉松属(Podocarpus)的一种雌雄异株的常绿小乔木,是罗汉松的一个变种植物,原产于日本和中国云南省,目前广泛分布于我国各地,栽植的最北界为淮河流域和山东半岛,它是罗汉松科植物在我国露地栽培纬度最高的种类[9]。短叶罗汉松的根皮、果实、叶与罗汉松(Podocarpusmacrophyllus)相似,可用作药材,具有杀虫、治癣疥、治疗心胃气痛等功能[10]。目前对短叶罗汉松的研究较少,主要集中在叶片结构的发育过程[10]、生药性状[11]和微形态结构[12]等方面,而对雌雄株叶片形态结构特征差异的研究鲜见报道。鉴于此,本文以营养生长期的短叶罗汉松雌、雄植株的成熟叶片作为研究对象,对雌、雄株叶片的解剖结构及表皮形态进行了比较分析,明确了叶片的性别差异,并且探讨了短叶罗汉松雌雄株叶片在不断进化过程中对生存环境的适应策略,可以为进一步了解和认识短叶罗汉松的第二性征,为其育种、栽培等生产实践提供理论依据。
1 材料与方法
1.1 试验地概况
实验材料采自广西壮族自治区南宁市隆安县那桐镇金穗园林花木场内生长的短叶罗汉松。该地位于广西的中部偏西南,地理坐标为107°21′~108°6′E,22°51′~23°21′N,气候类型为南亚热带季风气候,夏季炎热多雨,冬季温暖少雨,降水多集中在夏季,年均降水量为1300 mm,年均气温为21.8 ℃,其中,月平均最高气温约为28.4 ℃,月平均最低气温约为13.2 ℃。
1.2 实验材料
在样地中选取生长健壮和生长势相同的8年生雌、雄短叶罗汉松各10株,在不同方位、树冠外围中上部(顶芽下的第4~5节)分别剪取新鲜成熟健康的雌、雄株叶片各60片。用于测定叶片植纹和解剖结构的30片样品立即用FAA固定液保存;用于扫描电镜和透射电镜观察的30片样本置于保鲜盒中带回室内,先用缓冲液冲洗干净,再迅速置于浓度为2.5%的戊二醛固定液中,进行抽真空后于4 ℃冰箱中保存。
1.3 实验方法
1.3.1 叶表皮植纹的观察 从外形测量完毕的样品中随机选取10片,剪取叶片靠近中脉中部位置约长0.5 cm的部分,放入装有蒸馏水的试管中,在水浴锅中加热15~30 min,然后将蒸馏水换成1∶1的冰乙酸和过氧化氢的混合液,放在60~80 ℃的恒温中离析24~48 h,当叶片出现泛白并有泡沫絮状物时将叶片取出,用蒸馏水反复清洗,直到叶肉组织完全去除;取上、下表皮置于番红-固绿溶液中染色,用蒸馏水装片,放于Nikon E100显微镜下观察并拍照。雌、雄株至少分别取30个视野,每个指标测30个观测值,统计值使用平均值。观测指标有气孔长度、宽度,表皮细胞长度、宽度等,并计算气孔密度和气孔指数,计算公式为:气孔密度=气孔数量/叶片面积;气孔指数=气孔数/(气孔数+表皮细胞数)×100%。
1.3.2 叶片解剖结构的观察 样本、采集的视野和观测数量以及方法同1.3.1节。使用双面刀片徒手切片方法,挑选出薄而均匀的切片,用蒸馏水装片,置于Nikon E100显微镜下观察并拍照。测量指标有:栅栏组织、海绵组织、角质层、表皮等的厚度,并用以下公式计算相关指标:栅海比(P/S)=栅栏组织厚度/海绵组织厚度;组织结构紧密度(CTR)=(栅栏组织厚度/叶片厚度)×100%;组织结构疏松度(SR)=(海绵组织厚度/叶片厚度)×100%。
1.3.3 叶面扫描结构的观察 取出置于4 ℃冰箱中固定24 h的样品15片,吸出原固定液,注入3%戊二醛溶液固定2 h,然后用0.1 mol/L的磷酸溶液冲洗3次,每次15 min;冲洗后加入2%的四氧化锇缓冲溶液,于室温固定2 h(用于能谱分析的材料省略此步骤);用0.1%磷酸溶液清洗3次,每次15 min;依次使用50%、70%、80%、90%、100%的乙醇溶液进行梯度脱水,该步骤进行3次,每个梯度滞留10 min;然后用100%六甲二硅胺烷浸泡3次,每次10 min,再放于真空干燥器中进行抽真空干燥;最后粘样本到样本座,用Eiko离子溅射仪喷镀后放在TESCAN VEGF3扫描电镜下观察拍照,并使用EDAX能谱分析仪对叶片进行能谱分析,选取15个视野进行分析并拍照。观测参数有角质层纹饰、蜡质纹饰、气孔类型和叶片蜡质层元素重量之比。
1.3.4 叶片透射结构的观察 取样、固定、清洗等步骤同上,之后使用乙醇-丙酮进行脱水:50%乙醇→70%乙醇→90%乙醇→1∶1混合液(90%酒精∶90%丙酮)→90%丙酮→3次100%丙酮,每个梯度脱水15 min;用环氧树脂对样品进行浸透和包埋;分别将样品放在38、45和60 ℃中聚合15、12和24 h;使用徕卡UC7超薄切片机进行切片;用醋酸铀-柠檬酸铅双重染色后放在日立H-1650透射电镜下进行观察,选取15个视野进行拍照。观测参数有细胞长、宽度,线粒体长、宽度和叶绿体的长、宽度。
1.3.5 数据处理 使用Microsoft Excel 2019对各项结构的测量数据进行归纳,运用SPASS 23.0软件对叶片形态结构数量化的参数进行统计分析。
2 结果与分析
2.1 叶表皮植纹
短叶罗汉松雌、雄株的上、下表皮细胞均排列整齐,均由长、短细胞组成,上、下表皮的长细胞多为长矩形,上表皮的短细胞多为正方形,而下表皮的短细胞多为长矩形(图1)。气孔为典型的平列型,由两个保卫细胞组成,雌株的气孔稍大于雄株,其中气孔宽、气孔密度和气孔器长等性状与雄株存在极显著差异,而气孔长、气孔指数和气孔器宽不存在显著差异(表1)。
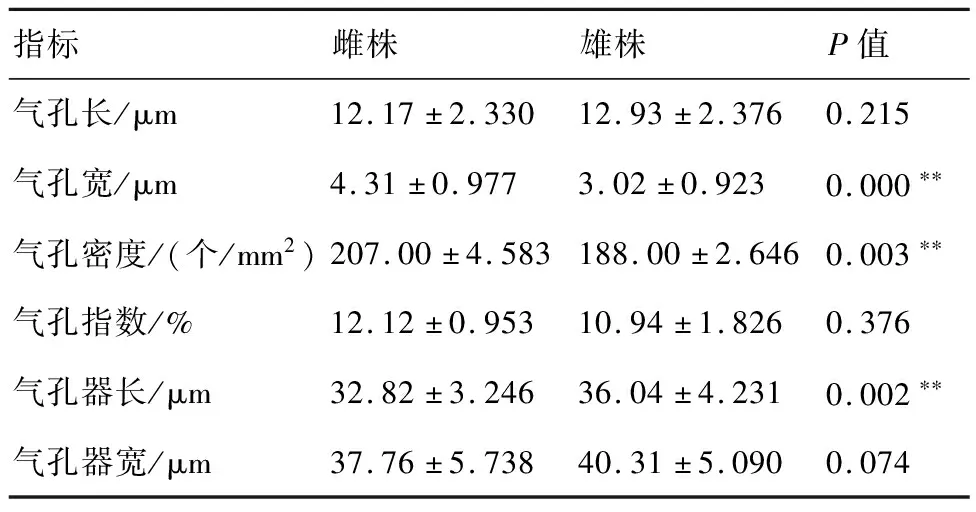
表1 短叶罗汉松雌、雄株叶表植纹结构的测量结果
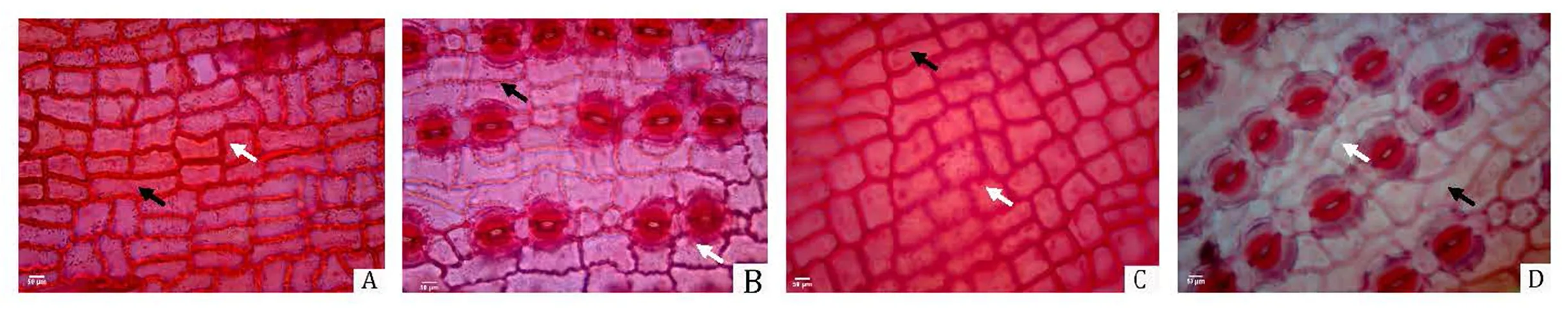
A:雌株叶片上表皮。B:雌株叶片下表皮。C:雄株叶片上表皮。D:雄株叶片下表皮。黑色箭头指长细胞;白色箭头指短细胞。
2.2 叶片解剖结构
短叶罗汉松雌、雄株的叶片都为异面叶,均由表皮、叶肉和叶脉组成(图2)。雌、雄株的叶片解剖结构存在差异(表2),其中雌株的上、下角质层厚度和叶中厚极显著大于雄株,而雄株的栅栏组织,上、下表皮厚度和致密度(CTR)极显著大于雌株,转输组织长和栅海比(P/S)则显著大于雌株;在雌、雄株之间海绵组织厚度、转输组织宽和细胞疏松度(SR)差异不显著。

A:雌株。B:雄株。黑色箭头指表皮;白色箭头指海绵组织;红色箭头指叶脉;蓝色箭头指栅栏组织。

表2 短叶罗汉松雌、雄株叶片解剖结构的测量结果
2.3 叶片表皮微形态结构
在扫描电镜下的叶片表面结构特征如表3所示,在上表皮中,雌、雄株的整体表面和角质层纹饰差异较大,雌株的表面为皱脊具近平行的凸起横纹,角质层纹饰为平滑的无片状(图3A);而雄株表面具有不规则的凸起状横纹和网状纹,角质层纹饰则为表面粗糙的鱼鳞状凸起与薄片(图3C)。在下表皮中,雌株表面为光滑的微凸横纹,细胞壁为凸起状(图3B);雄株表面有光滑的平整纹路,细胞壁呈凹陷状(图3D);雌、雄株两者的气孔器都呈平滑、成行排列状。
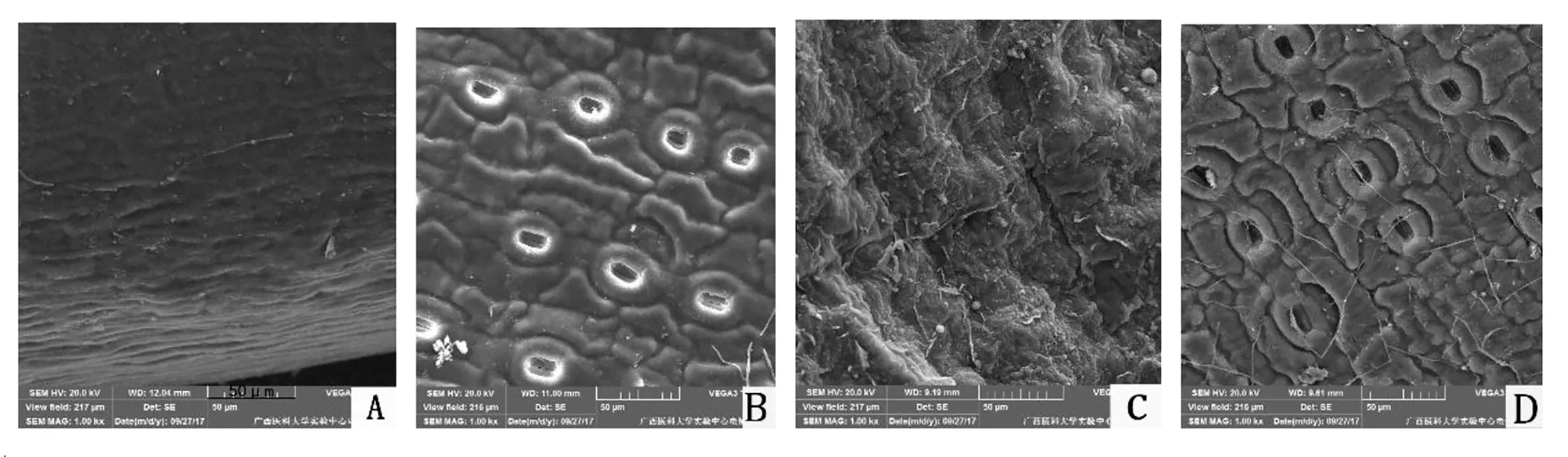
A:雌株叶片上表皮。B:雌株叶片下表皮。C:雄株叶片上表皮。D:雄株叶片下表皮。

表3 短叶罗汉松雌、雄株叶片表皮的扫描结构特征
2.4 叶片表面蜡质能谱分析
叶片表面蜡质能谱分析结果(表4)显示,雌、雄株的短叶罗汉松叶片中含有相同的9种元素,其中雌株的C、O、Si、Au等元素含量大于雄株,而雄株的Na、Mg、Al、Ca、K等元素含量则大于雌株。

表4 短叶罗汉松雌、雄株叶片表面蜡质元素质量百分比
2.5 叶片的超微结构
在透射电镜下观察,发现在雌、雄株叶片的栅栏组织内均存在线粒体和叶绿体(图4)。雌株中的线粒体和叶绿体稍大于雄株,但不存在显著差异。雌、雄株的细胞壁厚和细胞长存在显著差异,而细胞宽无显著差异(表5)。

A:雌株。B:雄株。Chl:叶绿体。CW:细胞壁。M:线粒体。

表5 在透射电镜下短叶罗汉松雌、雄株叶片超微结构的测量结果
3 结论与讨论
雌雄异株是植物适应生存环境变化的一种进化表现,雌雄异株植物对维持和稳定生态系统和保护物种多样性有着积极的作用,同时也是陆地生态系统的重要成员之一[8]。短叶罗汉松是罗汉松的一个变种植物,也是一种雌雄异株植物。在植物的进化过程中,叶片被认为是对环境变化最为敏感和具有较强可塑性的植物器官,可在各种压力下形成应对不同逆境的适应类型,其形态与解剖结构特征最能体现植物对环境的适应能力[13]。对雌雄异株的植物而言,雌、雄株的叶片在生理特性[14]、形态解剖结构[15-17]以及对环境因子的响应[8]等方面存在着显著的差异。在一般情况下,雄株的枝条大小、活力和克隆繁殖能力大于雌株[18],尤其是在条件艰苦的环境下,雄株的个体数量明显多于雌株,在空间上两者也存在着相互排斥的现象[19]。在本研究中,短叶罗汉松雌、雄株的叶片形态结构特征存在着明显的性别差异,这是短叶罗汉松雌雄株第二性征[20]在叶片上的表现。
一般认为植物叶片的表皮厚度、栅栏组织厚度、栅海比值和叶片致密度在一定程度上与其储水、控水和抗旱性成正比[17,21]。雄株短叶罗汉松叶片的表皮厚度、栅栏组织厚度、栅海比和致密度均大于雌株,表明雄株的保水能力和抗旱性强于雌株,这与前人的研究结果[15,22-23]一致。气孔是植物控制水分和进行气体交换的通道,不仅可以直接影响植物的蒸腾作用[24],还对植物的水分利用、营养吸收、物质代谢等方面以及整个生态环境的调节起重要作用[25]。雌株短叶罗汉松的气孔宽、气孔密度和上、下角质层厚度极显著大于雄株,说明雌株叶片不仅气体交换能力强,而且还可有效防止水分的过度蒸腾,从而促进植物的新陈代谢和保持水分。植物叶片的微形态结构为鉴别雌、雄株植物的差异提供了依据[12]。在扫描电镜下短叶罗汉松雌、雄株在叶片表面的蜡质纹饰和角质层纹饰等微形态方面存在着明显的区别,雄株叶片表面粗糙的鱼鳞状角质层纹饰的滞尘能力比雌株平滑无片状的角质层纹饰稍强一些,这可能是在叶片表面所含的9种蜡质元素中雄株有5种元素含量大于雌株的原因。
综上所述,短叶罗汉松雌、雄株叶片表皮的形态结构特征存在明显的性别差异,且叶片的各项指标表明雄株对环境的适应性和抗逆性强于雌株,这可能也是雄株短叶罗汉松在自然界广泛分布且应用频率高于雌株的原因。本研究结果有利于在实践中区分短叶罗汉松的雌、雄株,可为其育种和应用提供一定的理论参考。
