种植绿肥对作物产量和细菌群落稳定性的影响
2021-12-01李正鹏严清彪孙小凤
韩 梅,刘 蕊,李正鹏,严清彪,孙小凤
种植绿肥对作物产量和细菌群落稳定性的影响
韩 梅,刘 蕊,李正鹏,严清彪,孙小凤*
(青海大学农林科学院,青海 西宁 810016)
为深入研究青海高原地区长期种植绿肥,减施化肥条件下小麦/油菜产量及土壤理化性状及微生物群落的变化,通过2011年建立的定位试验,设GF0(毛苕子作为绿肥,不施用化肥)、GF60(毛叶苕子配施60%化肥)、GF70(毛叶苕子配施70%化肥)、GF80(毛叶苕子配施80%化肥)、GF90(毛叶苕子配施90%化肥)、GF100(毛叶苕子配施100%化肥)、F0(休耕并且不施化肥)、F100(休耕并且施用100%化肥)共8个处理,利用高通量测序等技术,研究了种植利用绿肥减施化肥条件下小麦和油菜的产量、土壤性质和微生物群落结构的变化特征.结果表明,在保证小麦、油菜不减产的基础上,种植绿肥后茬作物减施化肥达到30%左右.绿肥配施减量化肥对土壤肥力有显著的提升作用,特别是土壤有机碳和全氮含量有明显的提高,分别提高1.34%~7.46%、2.16%~7.48%.化肥与绿肥配施增强了土壤微生物群落多样性,其中酸杆菌门和变形菌门是本研究土壤中的优势菌群.共生网络分析表明,绿肥处理提高了细菌群落的稳定性和抗干扰能力,同时绿肥应用增加了细菌群落中的关键物种.本研究表明,不同土壤微生物丰度受种植模式、施肥量影响很大,长期种植绿肥、减施化肥提高了土壤微生物丰度和多样性,增强微生物群落的稳定性.种植绿肥条件下化肥减肥量30%左右保证作物稳产可为当地的施肥管理提供参考.
毛叶苕子;减肥;微生物群落;共发生网络;关键物种
种植绿肥是减少化肥投入、提高作物产量的传统有效方法[1-3].在许多农业系统中,使用绿肥可以降低土壤侵蚀风险、改善土壤特性、保证土壤健康、增加土壤微生物的丰度和活性[4-8].豆科植物因其生物固氮能力而成为农田绿肥中最常用的绿肥作物.毛叶苕子(Roth)因其适应性广、新鲜生物量大、固氮能力强等特点,被广泛应用为北方绿肥作物.在旱地地区,种植豆科绿肥比休耕更能提高土壤肥力,促进作物可持续生产[9-10].我国各种耕作制度下存在着大量季节性闲置耕地,并且不同区域有着多样的气候环境,适宜间作、轮作、混作等多种种植方式,发展绿肥具有很大潜力[11].青海省河湟地区位于青海高原的东北部属典型的两季不足、一季有余.为充分利用光热土资源,在青海高原建立了豆科绿肥-小麦/油菜轮作技术体系.毛叶苕子的种植不仅解决了青海高原休耕和裸地问题,而且改善了高原环境.经过多年的实践证明,该技术在优化作物栽培结构、提高复种指数和土壤质量、缓解牧区冬季牧草短缺问题等方面是一项有效的实践.种植绿肥有利于土壤微环境的改良、提升作物产量和品质等[12];翻压还田对后茬作物的生长与产量、养分吸收利用以及土壤物理、化学、生物特性等同样存在一定的影响[13].其在养分供应、培肥土壤、生物固氮、涵养水源等[14]方面均体现了其为农田提供的有价值的生态系统服务功能.绿肥作物在饲用价值方面也发挥着重要的作用,尤其在夏季饲料不足、冬季饲料缺乏的青藏高原地区.畜牧业的发展最重要的是饲料及饲草的质量,饲料短缺问题是畜牧业快速发展的阻碍[15].土壤微生物群落结构可以作为土壤肥力和土壤质量的有效指标.前人研究发现,绿肥可以改变微生物群落,从而影响土壤微生物诱导的土壤养分循环过程[16-17].持续种植绿肥可提高土壤微生物量碳、氮和土壤酶活性[18-19]. 而且种植翻压绿肥可以增加土壤微生物的数量和多样性,改变土壤微生物的组分和结构[20].土壤微生物群落结构的变化可能与长期种植绿肥改善土壤肥力、环境等有关[21]本研究在2011年建立的长期试验基础上,利用高通量测序技术,对小麦和油菜产量、土壤性质和微生物群落进行了研究.本研究在种植绿肥条件下减施化肥,调查毛叶苕子-小麦/油菜轮作系统作物产量、土壤性状及微生物群落结构和稳定性的影响.
1 材料与方法
1.1 试验地点
试验地点位于青海省西宁市青海省农林科学院(36°62′N,101°77′E, 海拔2300m),大陆性半干旱气候,年平均气温5.9℃,年平均降水量367.5mm,年平均蒸发量1729.8mm.土壤类型为栗钙土.2011年试验前,土壤有机碳、全氮、全磷和全钾含量分别为9670,1110,2180,26330mg/kg,速效氮、速效磷和速效钾分别为98,13.1,139mg /kg,土壤pH值为8.31.
1.2 试验设计
于2011年布置长期定位试验.该试验采用完全随机区组设计(表1、图1),共8个处理,(1)GF0,毛苕子作为绿肥,不施用化肥;(2)GF60,毛叶苕子配施60%化肥;(3)GF70,毛叶苕子配施70%化肥;(4)GF80,毛叶苕子配施80%化肥;(5)GF90,毛叶苕子配施90%化肥;(6)GF100,毛叶苕子配施100%化肥;(7)F0,休耕并且不施化肥;(8)F100,休耕并且施用100%化肥.休耕代表不种绿肥.各处理重复4次,试验中,100%化肥用量为当地农田的推荐用量,施N量为225kg/hm2,施P2O5量为112.5kg/hm2,不施钾肥.尿素(N 46%)和过磷酸钙(P2O512%)作为氮磷肥.

表1 试验方案

图1 大田照片
青海省的种植制度是一年一熟.本试验采用毛叶苕子-小麦/油菜轮作制度.小麦、油菜和毛叶苕子品种分别为“青春38号”、“浩油11号”和“土库曼毛苕”.毛叶苕子干物质中碳(C)、氮(N)、磷(P)、钾(K)平均含量分别为46.32%、2.45%、0.31%和1.82%.小麦或油菜在3月中旬施肥后播种,7月下旬收获,小麦、油菜播种量分别为600kg/hm2,7.5kg/hm2.小麦、油菜收获后,以105kg/hm2播种毛叶苕子.十月中旬,收割毛叶苕子,地上部分作为饲料移出田地,根茬还田.
1.3 样品采集与指标测定
2017年土壤采样共3次,分别为播种前(3月15日)、抽穗期(6月15日)和收获后(7月25日).采样后的土壤分取部分保存于4℃用于测定铵、硝酸盐氮,部分自然风干进行用于测定其他理化性状,部分保存于-80℃用于提取DNA.
采用重铬酸钾外加热法测定土壤有机碳(soil organic carbon,SOC).凯氏定氮法测定土壤全氮(total nitrogen,TN).用0.01mol/L CaCl2浸提NH4+-N和NO3--N,然后用连续流分析仪(AA3, SEAL, 德国)测定.采用碱解扩散法测定土壤速效氮(available N).采用电位法测定土壤 pH值(水土比 2.5:1)[22].
1.4 土壤微生物DNA提取及测序
采用E.Z.N.ATMMag-Bind Soil DNA Kit (OMEGA,美国)提取土壤微生物DNA.使用Nanodrop 2000分光光度计测定DNA含量.16S rRNA基因v3-v4序列引物为341F/805R (CCCT ACAC GACG CTCT TCCG ATCTG / GACT GGAG TTCC TTGG CACC CGAG AATT CCA)[23].利用Illumina MiSeq PE300平台进行测序[24].
1.5 生物信息学和统计分析
测序获得的原始reads使用FLASH (version 1.2.7, http://ccb.jhu.edu/software/FLASH/)进行提取、裁剪和质量筛选.利用Mothur软件(http://www.mothur.org/ wiki/Schloss_SOP)进行各处理Alpha多样性分析.使用Usearch(7.1版http://drive5.com/ uparse/)将剩余的高质量reads以97%的相似性进行操作分类单元(operational taxonomic units,OTUs)聚类[25].使用RDP分类器将OTUs的代表性序列与Sliva数据库进行比较[26].计算Good’s coverage[27]来估计样本的多样性和丰度.原始数据上传至国家基因组科学数据中心,序列号为SAMC392370- SAMC392373.
利用分子生态网络分析(Molecular Ecological Network Analyses pipeline,http://ieg2.ou.edu/MENA/)[28]对处理F0、F100、GF0和GF100进行共生网络分析..通过Gephi(http://gephi.github.io/)使生成可视化网络图.使用-score和-score定义关键物种(keystones),将网络中心点、模块中心点和连接节点作为分子生态网络中的关键物种[29-30].
2 结果与分析
2.1 绿肥配施化肥对小麦和油菜产量的影响

图2 2012~2018年小麦和油菜产量
由图2可见,绿肥配施化肥显著提高了油菜和小麦的产量.油菜GF100处理4a平均产量最高,达到1934kg/hm2,显著高于F100处理(1766kg/hm2).不同字母表示差异有统计学意义(<0.05).GF90、GF80和GF70处理的油菜产量与F100处理无显著差异,说明绿肥还田后减少10%~30%的化肥用量,而且油菜产量不减少.小麦GF100处理(6645kg/hm2) 3a平均产量显著高于F100处理(5873kg/hm2).GF90、GF80、GF70和GF60处理小麦产量均与F100处理无差异,说明保证小麦产量的前提下,绿肥根茬还田可减少小麦季化肥用量的40%.绿肥对提高作物高产和减少化肥的影响已得到广泛证实[2,31].在我国黄土高原建立的长期研究发现,施用豆科绿肥平均替代了31%的化学氮肥[32].在瑞士的田间研究发现,覆盖作物使玉米平均产量增加12%,种植毛叶苕子平均使玉米氮吸收增加79kg/ hm2[33].
2.2 绿肥配施化肥对土壤理化性状的影响
种植绿肥7年后,由于绿肥的生长和根茬的分解,土壤性状发生了改变(表3).在小麦播种,抽穗和收获阶段,土壤pH值的变化趋势是相似的,与F0相比,F100处理显著增加土壤pH值,而绿肥降低了土壤pH值.绿肥和化肥配施提高了土壤有机碳和总氮含量,GF70、GF60和GF80处理有机碳含量分别在播种、抽穗期和收获期最高.播期,GF60、GF100和F100处理土壤铵态氮含量显著高于其他处理. GF100和F100处理在抽穗期和收获期最高.绿肥处理降低了3个生育期的硝态氮含量.速效氮以播种期GF100处理最高,抽穗期和收获期F100处理最高.持续施用绿肥增加了土壤有机碳和易氧化碳的含量[10].西北地区夏季绿肥耕作优于休耕[10,32].将绿肥纳入农业轮作可以影响土壤碳库,并有助于土壤有机碳储量[34].

表3 三个生育阶段不同处理中的土壤理化性状
注:同一时期同一列内不同字母表示处理间有显著差异(<0.05).
2.3 绿肥配施化肥对细菌群落结构的影响
由图3可见,在属水平上,三个采样阶段Gp6、鞘单胞菌属()和芽单胞菌属()的丰度最高,分别被划分为酸杆菌门(Acidobacteria)、变形菌门(Proteobacteria和芽单胞菌门(Gemmatimonadetes).Gp6在播种、抽穗期和收获期分别占8.19%、8.08%和7.89%.鞘氨单胞菌的丰度在播种期高于芽单胞菌,而芽单胞菌的丰度在抽穗期和采收期高于芽单胞菌.在门水平上,变形菌门(Proteobacteria)最为丰富,在3个采样阶段占总reads的36%以上.酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes)和芽单胞菌(Gemmatimonadetes)的平均丰度分别为20.52%、8.25%和7.00%.在播种期,浮霉菌门(Planctomycetes)的丰度低于厚壁菌门(Firmicutes)和疣菌门(Verrucomicrobia),而在抽穗期和采收期则高于厚壁菌门和疣菌门.
为了进一步研究不同处理下的群落结构变化,分析了处理间丰度较高的8个菌门(相对丰度> 4%)的分布(图4).播种期,GF0处理的浮霉菌门丰度显著高于GF70和GF80处理;GF70处理的疣菌门丰度显著高于GF60和F100处理;F100处理的厚壁菌门丰度显著高于GF90处理.抽穗期不同处理改变了变形菌门、拟杆菌门、疣菌门、厚壁菌门和放线菌门(Actinobacteria)的分布.与GF90和F100处理相比, GF100处理显著增加了变形菌门的丰度;与GF0和GF60处理相比,GF90处理显著增加拟杆菌门的丰度;与GF60和F0处理相比,GF90处理显著增加了疣菌门的丰度;与GF70、GF80、GF90和GF100处理相比,F0处理显著增加了厚壁菌门的丰度;与GF0和GF90处理相比,GF80和F100处理显著提高了放线菌的丰度.收获期8个丰度较高的菌群在不同处理间差异不显著.绿肥的种植可以有效地提高土壤酶和土壤生物活性[19,35-37].土壤微生物驱动生物地球化学循环,特别是土壤碳、氮、硫等的转化[8,38].在藏西北地区的研究发现,在高海拔土壤中,酸杆菌门、绿杆菌门和变形菌门最为丰富,土壤总碳和碳/氮比值等土壤性质是该研究区域微生物分布的主要驱动因素[39].在本研究中,最丰富的门是酸杆菌门和变形菌门,这与邻近地区的研究一致.

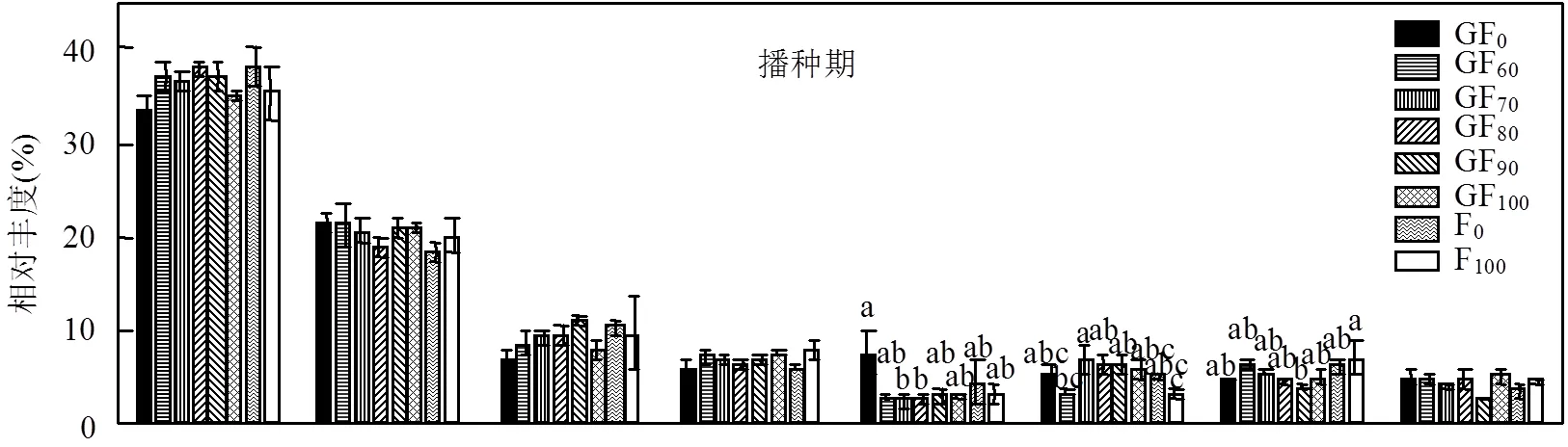
2.4 细菌群落的相互作用关系和网络拓扑特征
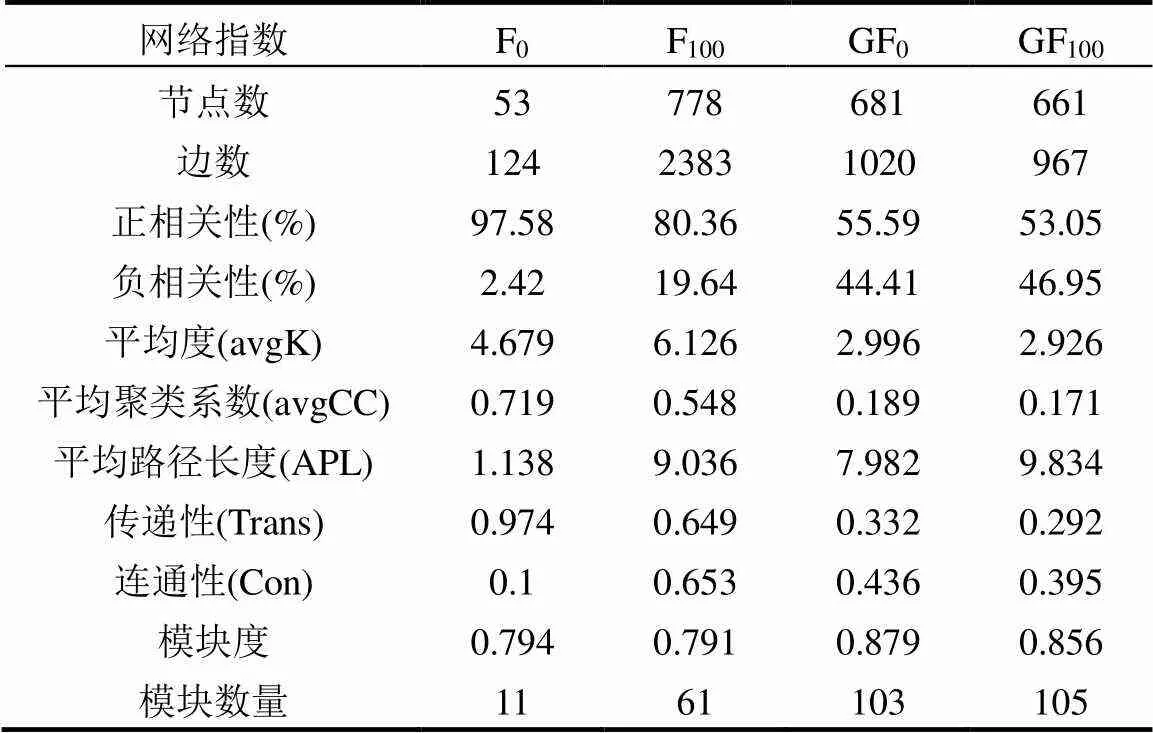
表4 细菌共生的网络特性
由表4和图5可见,F100的网络最复杂,其次是GF0和GF100,F0的网络最简单.F100处理的节点数和边数最高(分别为778和2383),F0处理的节点数和边数最低(分别为53和124).GF0和GF100处理的节点数和边数相似.绿肥处理(GF0和GF100)与无绿肥处理(F0和F100)呈负相关.GF0和GF100处理的平均连通性和平均聚类系数均低于F0和F100处理.GF0和GF100的网络模块度和模块数均高于F0和F100,说明绿肥的施用增加了网络模块度.通过构建并比较了添加绿肥和不添加绿肥处理下土壤细菌网络的结构,以评价物种间的相互作用.

图5 细菌群落的相互作用网络
不同颜色的节点代表不同的模块,节点大小代表节点的度,红色连接线代表节点间正相互作用,蓝色连接线代表节点间负相互作用
网络的模块化值在0.794到0.879之间,高于建议的阈值0.4,说明本研究构建的网络遵循模块化结构[21,40].节点(特定OTUs)和边(正或负链接)的数量是各种生物相互作用的表征[41].这些相互作用创造了复杂的微生物群落,并建立了特定生态系统中的群落特征[42].处理中没有绿肥(F0和F100)大多是正相关性,而施用绿肥不论是否添加化肥,都大大增加了网络的负相关比.这些结果结合节点和边的数目,表明应用毛叶苕子随着绿肥增加细菌群落的稳定性和抗干扰能力.更高的平均度(avgK)意味着更复杂的网络[28].平均度以F100处理最高,F0处理次之,均高于绿肥处理.化肥使细菌群落变得更加复杂而不稳定.环境特征,如温度和土壤pH值在构建网络交互作用中起重要作用[28].
进一步研究OTU的拓扑作用,以确定关键物种[30].从关系定义的各节点的网络角色可以看出,绿肥和化肥不同处理共生网络的拓扑性质和关键物种不同(图6).在F0处理中,没有找到关键物种,所有节点都被定义为外围节点.F100处理中, 0.77%被定义为模块中心点,0.26%的被定义为连接节点.在GF0处理中,定义为模块中心点和连接节点比例分别为1.32%和0.73%.在GF100中,0.76%被定义为模块模块中心点,没有连接节点.而GF0处理中模块集线器和连接器的比例最大,说明绿肥的利用增加了细菌群落的关键物种.与特定土壤过程相关的生物在微生物相互作用中发挥着更重要的作用[43],更多的关键物种在土壤地球化学过程中发挥着更重要的作用.绿肥的利用可以通过改变关键物种的分布来改善土壤质量.OTUs网络中关键物种间互作仍需进一步研究,以确定与绿肥和施肥管理相关的生理特征.

3 结论
3.1 绿肥提高作物产量和土壤肥力
在保持油菜、小麦产量的前提下,长期翻压毛叶苕子根茬能减施10%~40%化肥.绿肥配施减量氮肥可以化肥增加土壤有机碳和全氮含量.
3.2 绿肥配施化肥对细菌群落结构的影响
播种期,绿肥配施70%的化肥处理的疣菌门丰度显著高于GF60和F100处理;抽穗期绿肥配施化肥处理显著增加了变形菌门、拟杆菌门、疣菌门、厚壁菌门、放线菌的丰度.
3.3 绿肥增加细菌群落的网络稳定性
绿肥不论是否添加化肥,都大大增加了网络的负相关比.绿肥增加细菌群落的稳定性和抗干扰能力,增加了细菌群落的关键物种.
[1] Yang L, Bai J, Liu J et al. Green Manuring Effect on Changes of Soil Nitrogen Fractions, Maize Growth, and Nutrient Uptake [J]. Agronomy. 2018,8(11).
[2] Xie Z, Tu S, Shah F, et al. Substitution of fertilizer-N by green manure improves the sustainability of yield in double-rice cropping system in south China [J]. Field Crops Research. 2016188:142-149.
[3] 曹卫东,包兴国,徐昌旭,等.中国绿肥科研60年回顾与未来展望[J]. 植物营养与肥料学报, 2017,23(6):1450-1461.
Cao W D, Bao X G, Xu C X, et al. Reviews and prospects on science and technology of green manure in China [J]. Journal of Plant Nutrition and Fertilizer Science, 2017,23(6):1450–1461.
[4] Abdalla M, Hastings A, Cheng K, et al. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity [J]. Global Change Biology, 2019.
[5] Drinkwater L E, Snapp S. Nutrients in agroecosystems: rethinking the management paradigm [J]. Advances in Agronomy, 2007,92:163-186.
[6] Elfstrand S, Hedlund K, M?Rtensson A. Soil enzyme activities, microbial community composition and function after 47years of continuous green manuring [J]. Applied Soil Ecology, 2007,35(3): 610-621.
[7] Tejada M, Gonzalez J L, García-Martínez A M, et al. Effects of different green manures on soil biological properties and maize yield [J]. Bioresource Technology, 2008,99(6):1758-1767.
[8] 高嵩涓,周国朋,曹卫东.南方稻田紫云英作冬绿肥的增产节肥效应与机制[J]. 植物营养与肥料学报, 2020,26(12):2115-2126.
Gao S J, Zhou G P, Cao W D. Effects of milk vetch () as winter green manure on rice yield and rate of fertilizer application in rice paddies in south China [J]. Journal of Plant Nutrition and Fertilizer Science, 2020,26(12):2115–2126.
[9] Yao Z, Zhang D, Liu N, et al. Dynamics and sequestration potential of soil organic carbon and total nitrogen stocks of Leguminous Green Manure-Based Cropping Systems [J]. Soil & Tillage Research, 2019, 191:108-116.
[10] Zhang D, Yao P, Zhao N, et al. Building up the soil carbon pool via the cultivation of green manure crops in the Loess Plateau of China [J]. Geoderma, 2019,337:425-433.
[11] 曹卫东,黄鸿翔.关于我国恢复和发展绿肥若干问题的思考 [J]. 中国土壤与肥料, 2009:1-3.
Cao W D, Huang H X. Ideas on restoration and development of green manures in China [J]. Soil and Fertilizer in China, 2019,337:425-433.
[12] 栾好安.三峡库区橘园绿肥的生态效应及其对柑橘产量和品质的影响 [D]. 武汉:华中农业大学, 2015.
Luan Hao-an. Effects of planting green manure on ecological environment, yield and quality of citrus in citrus orchard at Three Gorges Reservoir [D]. Wuhan: Huazhong Agricultural University, 2015.
[13] 李富翠,赵护兵,王朝辉,等.渭北旱地夏闲期秸秆还田和种植绿肥对土壤水分、养分和冬小麦产量的影响 [J]. 农业环境科学学报, 2011,30(9):1861-1871
Li F C, Zhao H B, Wang Z H, et al. Effects of straw mulching and planting green manure on soil water, nutrient and winter wheat yield on Weibei Plateau, China [J]. Journal of Agro-Environment Science, 2011,30(9):1861-1871.
[14] 韩 梅,胥婷婷,曹卫东.青海高原长期复种绿肥毛叶苕子对土壤供氮能力的影响 [J]. 干旱地区农业研究, 2018,36(6):104-109.
Han M, Xu T T, Cao W D. Effects of long-term green manurehairy vetch on soil nitrogen supply on the Qinghai Plateau [J]. Agricultural Research in the Arid Areas, 2018,36(6):104-109.
[15] 王宗芳,宋文涛.绿色、优质饲用农作物种植技术的推广应用分析 [J]. 农业开发与装备, 2020,(1):60-61.
Wang Z F, Song W T. Analysis on the popularization and application of green and high quality forage crop planting technology [J]. Agricultural development and equipment, 2020,(1):60-61.
[16] Gao S J, Katsu-Yoshi S, Thorup-Kristensen K, et al. Long-term rice-rice-green manure rotation changing the microbial communities in typical red paddy soil in South China [J]. Journal of Integrative Agriculture, 2015,(12):2512-2520.
[17] Wang Y, Liu X, Butterly C, et al. pH change, carbon and nitrogen mineralization in paddy soils as affected by Chinese milk vetch addition and soil water regime [J]. Journal of Soils & Sediments, 2013,13(4):654-663.
[18] Elfstrand S, B?Th B, M?Rtensson A. Influence of various forms of green manure amendment on soil microbial community composition, enzyme activity and nutrient levels in leek [J]. Applied Soil Ecology, 2007,36(1):70-82.
[19] Ye X, Liu H, Li Z, et al. Effects of green manure continuous application on soil microbial biomass and enzyme activity [J]. Journal of Plant Nutrition, 2014,37(4):498-508.
[20] Selvi R V, Kalpana R. Potentials of green manure in integrated nutrient management for rice—A review [J]. Agricultural Reviews, 2009, 30(1):40-47.
[21] Zhang X X, Gao J S, Gao Y H, et al. Long-term rice and green manure rotation alters the endophytic bacterial communities of the rice root [J]. Microbial Ecology, 2013,66:917-926.
[22] 鲍士旦.土壤农化分析.3版:中国农业出版社, 2000.
Bao S D. Soil and agriculturai chemistry analysis [M]. Beijing: China Agriculture Press, 2000.
[23] 安毅夫,孙 娟,高 扬,等.长期放射性环境下微生物群落多样性变化 [J]. 中国环境科学, 2021,41(2):923-929.
An Y F, Sun J, Gao Y, et al. Variation of microbial community diversity with long-term exposure of radionuclides in dry uranium tailings pond J]. China Environmental Science, 2021,41(2):923-929.
[24] 郭宁宁,李 希,周训军,等.亚热带丘陵区人工湿地底栖动物群落特征研究 [J]. 中国环境科学, 2021,41(2):930-940.
Guo N N, Li X, Zhou X J, et al. Characteristics of zoobenthos community in constructed wetlands in subtropical hilly area [J]. China Environmental Science, 2021,41(2):930-940.
[25] Edgar R C. UPARSE: highly accurate OTU sequences from microbial amplicon reads [J]. Nature methods, 2013,10(10):996-998.
[26] Wang Q, Garrity G M, Tiedje J M, et al. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy [J]. Applied and environmental microbiology, 2007,73(16):5261-5267.
[27] Gi J. The population faequencies of species and the estimation of population parameters [J]. Biometrika, 1953,(3/4):237-264.
[28] Deng Y, Jiang Y H, Yang Y, et al. Molecular ecological network analyses [J]. BMC Bioinformatics, 2012,13(1):1-20.
[29] Poudel R, Jumpponen A, Schlatter D, et al. Microbiome networks: A systems framework for identifying candidate microbial assemblages for disease management [J]. Phytopathology, 2016,106(10).
[30] Olesen J M, Bascompte J, Dupont Y L, et al. The modularity of pollination networks [J]. Proceedings of the National Academy of Sciences, 2007,104(50):19891-19896.
[31] Astier M, Maass J M, Etchevers-Barra J D, et al. Short-term green manure and tillage management effects on maize yield and soil quality in an Andisol [J]. Soil & Tillage Research, 2006,88(1/2):153-159.
[32] Dabin Z, Pengwei Y, Na Z, et al. Contribution of green manure legumes to nitrogen dynamics in traditional winter wheat cropping system in the Loess Plateau of China [J]. European Journal of Agronomy, 2016,72:47-55.
[33] Wittwer R A, van der Heijden M G. Cover crops as a tool to reduce reliance on intensive tillage and nitrogen fertilization in conventional arable cropping systems [J]. Field Crops Research, 2020,249:107736.
[34] Jja B, Xuan D C, Msra D, et al. A meta-analysis of global cropland soil carbon changes due to cover cropping [J]. Soil Biology & Biochemistry, 2020,143:108389.
[35] Zhou G, Gao S, Lu Y, et al. Co-incorporation of green manure and rice straw improves rice production, soil chemical, biochemical and microbiological properties in a typical paddy field in southern China [J]. Soil and Tillage Research, 2020,197:104499.
[36] Nevins C J, Lacey C, Armstrong S. The synchrony of cover crop decomposition, enzyme activity, and nitrogen availability in a corn agroecosystem in the Midwest United States – ScienceDirect [J]. Soil & Tillage Research, 2020,197.
[37] Piotrowska A, Wilczewski E. Effects of catch crops cultivated for green manure and mineral nitrogen fertilization on soil enzyme activities and chemical properties [J]. Geoderma, 2012,189-190 (Complete):72-80.
[38] Soman C, Li D, Wander M M, et al. Long-term fertilizer and crop-rotation treatments differentially affect soil bacterial community structure [J]. Plant & Soil., 2017.
[39] Chu Haiyan, Sun Huaibo, Binu M, et al. Bacterial community dissimilarity between the surface and subsurface soils equals horizontal differences over several kilometers in the western Tibetan Plateau [J]. Environmental Microbiology, 2016,18(5): 1523-1533.
[40] Newman M E J. Modularity and community structure in networks [J]. PNAS, 2006,103(23):8577-8582.
[41] Jose M, Montoya J, Pimm S, et al. Ecological networks and their fragility [J]. Nature., 2006,442(7100):259-264.
[42] Jiang Y, Sun B, Li H, et al. Aggregate-related changes in network patterns of nematodes and ammonia oxidizers in an acidic soil [J]. Soil Biology and Biochemistry, 2015,88:101-109.
[43] Yan C, Ruibo S, Ting S, et al. Evidence for involvement of keystone fungal taxa in organic phosphorus mineralization in subtropical soil and the impact of labile carbon [J]. Soil Biology & Biochemistry, 2020, 148.
Green manuring increased crop yields and the stability of bacterial community.
HAN Mei, LIU Rui, LI Zheng-peng, YAN Qing-biao, SUN Xiao-feng*
(Academy of Agriculture and Forestry, Qinghai University, Xining 810016, China)., 2021,41(11):5382~5390
In order to provide theoretical support for the effects of reducedfertilizer in hairy vetch -wheat/oilseed rape rotation system on crop production and soil fertility in Qinghai Plateau. a location field experiment about hairy vetch -wheat/oilseed rape rotation established in 2011 was used in this study. The experiment including eight treatments, e.g., only green manure (GF0), green manure and 60% of chemical fertilizer (GF60), green manure and 70% of chemical fertilizer (GF70), green manure and 80% of chemical fertilizer (GF80), green manure and 90% of chemical fertilizer (GF90), green manure and 100% of chemical fertilizer (GF100), only 100% of chemical fertilizer (F100), and the no-fertilizer control (F0). The wheat and oilseed rape yields, soil properties and microbial community structures were measured. Results showed that the green manure -wheat/oilseed rape rotation system could reduce about 30% chemical fertilizer without reduction of wheat and oilseed rape yields, when compared with F100. Green manure combined with reduced chemical fertilizer improved soil fertility, especially increasingthe contents of soil organic C and total N by 1.34%~7.46% and 2.16%~7.48%, respectively.Green manure combined with reduced chemical fertilizer increased the diversity of soil microbial, and the most abundant phyla were Acidobacteria and Proteobacteria. The results of co-occurrence networks showed that green manuring increased the percentages of negative correlations andmodularity of the networks, indicating that application of hairy vetch as green manure increased the stability and anti-jamming capability of the bacterial communities. The network roles suggested that utilization of green manure increased the keystones in the bacterial community. It was concluded that the microbial diversity and abundance were affected by the cropping system and fertilization amounts. The utilization of green manure can reduce 30% of chemical fertilizer without reduction of wheat and oilseed rape yields. The reduced fertilizer amount could be a reference to the local fertilizer management practices and will give a theoretical support for this agricultural system.
hairy vetch;reduced fertilizer;microbial community;co-occurrence networks;keystones
X171.5,X172
A
1000-6923(2021)11-5382-09
韩 梅(1973-),女,青海湟中人,副研究员,硕士,主要从事绿肥方面研究.发表论文40余篇.
2021-04-02
财政部和农业农村部:国家现代农业产业技术体系资助(绿肥, CARS-22);青海省农科院基金项目(2019-NKY-06)
* 责任作者, 研究员, 610193056@qq.com
