甘肃、青海和宁夏地区牛病毒性腹泻病毒Erns基因的变异分析
2021-11-24高闪电王锦明田占成独军政王建东常惠芸关贵全李有全
高闪电,王锦明,田占成,独军政,王建东,常惠芸,关贵全,李有全,殷 宏, 2*
(1.中国农业科学院兰州兽医研究所 家畜疫病病原生物学国家重点实验室,兰州 730046;2.江苏省动物重要疫病与人兽共患病防控协同创新中心,扬州 225009;3.宁夏农林科学院动物科学研究所,银川 750002)
牛病毒性腹泻病毒(bovine viral diarrhea virus,BVDV)属于黄病毒科(Flaviviridae)、瘟病毒属(Pestivirus),主要感染牛,导致腹泻、呼吸道症状、流产或黏膜糜烂[1]。病毒基因组RNA包含5′UTR、开放阅读框以及3′UTR保守的结构,编码C、Erns、E1、E2四种结构蛋白和7~8种非结构蛋白。Erns、E1、E2镶嵌在病毒脂质双层囊膜上,与囊膜内由C蛋白和病毒RNA构成的衣壳共同组成完整的病毒粒子[2]。E2是BVDV最主要的抗原蛋白,在诱导宿主体液免疫和细胞免疫中发挥重要的作用,是决定疫苗免疫保护的关键蛋白。Erns为高度糖基化蛋白,具RNA酶活性,除参与病毒粒子组成,还可从感染细胞分泌至胞外。Erns作为瘟病毒特有蛋白,也可诱导中和抗体的产生,可作为病毒持续感染的标志物,广泛用于BVDV持续与急性感染的鉴别诊断[3],但相关研究发现检测Erns的免疫组织化学法或商品化试剂盒存在漏检现象,表明Erns蛋白在毒株之间存在抗原差异[4]。
BVDV分离株间抗原性、基因组核苷酸存在较大差异,目前主要依据5′UTR、N基因核苷酸差异将其分为BVDV-1、BVDV-2、BVDV-3基因型以及BVDV-1a~BVDV-1w、BVDV-2a~2d等基因亚型[5-6]。根据分离株的细胞培养特性,可将BVDV分为非致细胞病变(ncp)和致细胞病变(cp)两种生物型,田间分离株多为ncp型且与牛急性感染、流产和持续性感染(PI)有关,cp型BVDV感染PI牛可导致黏膜病。在我国报道的BVDV分离株表现为较高的亚型多样性,可分为13种基因亚型(BVDV-1:11种;BVDV-2:2种)及BVDV-3基因型[6-12]。目前我国BVDV流行株的抗原基因E2的多态性已初步阐明[13],但关于毒株Erns基因的信息较少。为了系统了解我国BVDV流行株Erns基因的分子特征及遗传演化规律,本研究测定分析了2013—2019年间我国甘肃、青海、宁夏省区主要BVDV流行株的Erns基因序列,以期进一步丰富我国BVDV的抗原变异基础研究,为疫苗的分子设计提供必要的技术储备。
1 材料与方法
1.1 材料
2013—2019年,甘肃、青海、宁夏规模化牛场送检的疑似牛病毒性腹泻发病牛的EDTA抗凝血150份(表1),冻存于-80 ℃。致细胞病变生物型对照毒株BVDV-AV69 购自中国兽医药品监察所,无BVDV污染的牛肾细胞(MDBK)、BVDV-1p基因亚型分离株Camel-5,由本实验室保存,BVDV抗原检测试剂盒为爱德士公司产品,FITC标记的猪抗BVDV多克隆抗体为美国VMRD公司产品,RNAsimple总RNA提取试剂盒、DNA纯化回收试剂盒购自天根生化科技(北京)有限公司,一步法RT-PCR试剂盒HiScript Ⅱ One Step RT-PCR Kit、DH5α 化学感受态细胞、质粒提取试剂盒为南京诺唯赞生物科技股份有限公司产品,Trans2K DNA Marker购自北京全式金生物技术有限公司,pMD18-T 克隆载体购自宝日医生物技术(北京)有限公司。
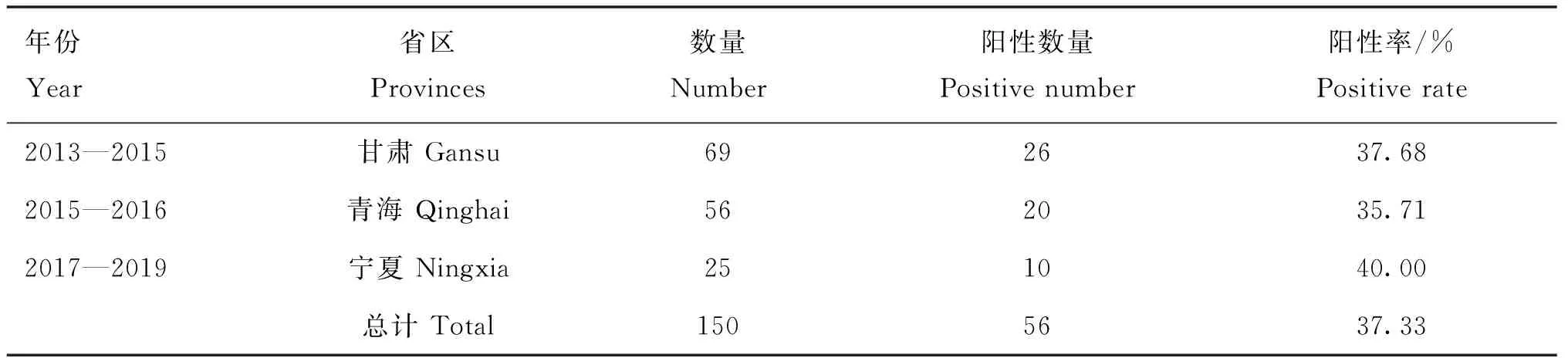
表1 样品信息及BVDV RT-PCR检测结果
1.2 引物
根据GenBank中收录的BVDV-1、BVDV-2、BVDV-3基因组序列,设计引物ErnsF:5′-AGCATTGTTRGCRTGGGC-3′,E1R:5′-AACCAYAGTATRCCTTGYA-3′,用于扩增BVDV基因组Erns-E1区,预期目的片段为1 296 bp。同时合成引物5′UTRF(5′-CTAGCCATGCCCTTAGTAGGACTA-3′)和5′UTRR(5′-CAACTCCATGTGCCATGTACAGCA-3′),用于扩增BVDV基因组5′UTR区。
1.3 RNA提取及RT-PCR
参照RNAsimple 总RNA提取试剂盒说明书,分别从250 μL EDTA抗凝血和Camel-5株培养物提取总RNA。利用一步法RT-PCR试剂盒HiScript II One Step RT-PCR Kit,以50 μL反应体系扩增Erns-E1 DNA片段。电泳切胶回收后与pMD18-T 载体连接,转化DH5α感受态细胞,PCR鉴定并将阳性克隆摇菌,委托生工生物工程(上海)公司测序。同时选取测序正确的样品RNA扩增5′UTR区并克隆测序。
1.4 病毒分离
根据爱德士BVDV抗原检测试剂盒操作说明检测抗凝血中BVDV抗原,参照文献[14],利用阳性抗凝血样品制备牛淋巴细胞裂解液并接种至MDBK单层细胞,盲传3代后,接种MDBK单层细胞,利用FITC标记的猪抗BVDV多克隆抗体进行免疫荧光检测细胞培养物中的BVDV抗原。
1.5 序列分析
利用VecScreen、DNASTAR、BioEdit等软件对获得的序列进行分析。根据GenBank数据库中收录的SD-15毒株(登录号为M96751)的Erns基因和5′UTR区,以Clustal W方法比对获得Erns基因和5′UTR区核苷酸信息。利用DnaSP 5.10、BioEdit 分析Erns基因核苷酸以及推导氨基酸序列的变异位点。利用BLAST在线分析序列与GenBank中收录毒株Erns基因的同源性,并利用Mega7.0软件包构建系统进化树。
2 结 果
2.1 基因克隆及序列测定
从150份抗凝血提取总RNA,经RT-PCR扩增病毒Erns-E1区(1 296 bp),共获得56份DNA,测序均为BVDV特异序列,总体阳性率为37.33%,其中甘肃省、青海省、宁夏回族自治区BVDV阳性率分别为37.68%、35.71%、40.00%。比对并去掉相同序列后获得33条Erns序列,长度均为681 bp,无核苷酸插入或缺失,编号并提交GenBank数据库,登录号为KY675201~KY675227、MW560180~MW560185。
2.2 病毒分离与相似性分析
GenBank中未收录BVDV-1p亚型毒株的Erns序列,因此测定实验室保存的Camel-5株Erns序列(登录号为KY675228)并作为参考序列。分析发现这些流行株属于9个基因亚型:BVDV-1a(2株)、BVDV-1b(5株)、BVDV-1c(1株)、BVDV-1d(3株)、BVDV-1m(11株)、BVDV-1o(1株)、BVDV-1p(4株)、BVDV-1q(4株)、BVDV-2a(1株)。利用BVDV抗原阳性抗凝血GS5、QH4、QH9、NX1、NX3、NX201902进行病毒分离,在MDBK细胞未见细胞病变,经免疫荧光鉴定分离株均为非致细胞病变型,利用细胞培养物提取RNA并扩增Erns基因测序,与利用抗凝血样品扩增测序结果一致。在这些亚型株中,BVDV-2a亚型NX3株Erns基因序列与美国分离株9231(登录号:MH806437)相似性最高(97.75%),高于国内新疆分离株XJ04(FJ527854,94.73%)、山东分离株SD-1(MK599227,92.13%)以及吉林分离株JZ05-1(GQ888686,91.59%)。GS11、QH2、QH11、QH18株Erns序列相似性为89.4%~98.9%,与Camel-5株Erns基因核苷酸序相似性为90.0%~95.1%,同属BVDV-1p基因亚型。NX2019/02株与GenBank中收录的BVDV-1m亚型 SD-15 株Erns基因(登录号:KR866116)核苷酸一致性为84.73%,其5′UTR与1v毒株EN-19(登录号:MN417826)的核苷酸一致性为96.39%,因此暂定NX2019/02株为BVDV-1v基因亚型。各基因亚型株间Erns序列相似性矩阵结果表明,BVDV Erns核苷酸在1a~1d经典亚型相似性较高(79.8%~85.9%)、在1m~1v较新亚型也表现为较高相似性(81.0%~87.3%),以BVDV-1m和BVDV-1p流行株亚型间相似性最高(87.3%),与作者前期在各亚型BVDV株E2基因比较结果类似[13]。Erns基因在经典亚型组和新亚型组株间差异较大,以BVDV-1d亚型与BVDV-1p亚型流行株间相似性最低(76.7%)。BVDV-1v NX2019/02株与BVDV-1m~1q亚型BVDV株Erns序列相似性(82.0%~86.3%)高于经典亚型BVDV-1a~1d(77.8%~80.7%),表明BVDV-1m~1v等新出现毒株在起源上关系较为密切。
2.3 基因变异性分析
变异位点分析显示流行株Erns基因核苷酸序列和推导的氨基酸序列变异位点分别为49.34%(336/681)和26.43%(60/227),小于作者前期测定的BVDVE2基因(核苷酸和氨基酸变异位点分别为61.76%和60.69%)。在推导的Erns氨基酸序列,His30、His74、Glu75、Lys78、His79RNA酶活性位点以及139KKGK142dsRNA作用基序保守。在Erns蛋白存在7个N-糖基化位点,位于氨基酸第2、11、26、65、95、100、217位,其中第26位糖基化位点(26 NRSL)在BVDV-1m、1o、1p、1q、1v亚型株糖基化位点移位(24 NVSR)(图1)。此外,在1m亚型GS25株、1p亚型QH2株参与链内二硫键形成的第171位半胱氨酸发生变异,1 d亚型QH9株该位点半胱氨酸转位至第181位。

图1 各亚型BVDV株Erns N-糖基化位点(N95、N100、N217糖基化位点未显示)
2.4 系统进化分析
利用Mega7.0对获得Erns基因核苷酸序列与GenBank中收录的参考株进行序列比对并进行系统进化分析,结果表明,在西北地区BVDV表现为区域亚型多样性(甘肃省7种:1a、1b、1c、1m、1o、1p、1q基因亚型;青海省5种:1b、1d、1m、1p、1q基因亚型;宁夏区5种:1d、1m、1q、2a以及1v新基因亚型)。在3省区均存在1m、1q基因亚型BVDV株,以1m基因亚型株最为普遍(33.33%,11/33),且1m 亚型株Erns基因相似性较高(93.5%~97.9%),未发现明显地域性差异;其次为甘肃及青海2省共存的1b亚型(5/33)和1p亚型(4/33)(图2A)。为验证基于Erns基因的系统进化分析结果,进一步扩增测定NX2019/02株等19条5′UTR序列,构建系统进化树(图2B),证实NX2019/02株属BVDV-1v新基因亚型。1p基因亚型双峰驼分离株Camel5Erns基因与甘肃株GS11的Erns基因核苷酸相似性(95.1%)高于QH2、QH11等青海流行株(90.0%~90.4%),1q基因亚型GS26株、QH5株、QH8株分别与Camel-6双峰驼分离株以及SD0803猪源分离株存在较高Erns基因相似性(分别为98.3%和96.4%~98.6%)。利用Erns基因进化树和5′UTR进化树分析BVDV株亲缘关系结果较一致,但5′UTR 进化树揭示GS6株与青海分离株的进化关系效果略次于Erns基因进化树(图2A、B)。由于Erns基因在BVDV株间差异性略小于E2基因但显著高于5′UTR区段,因此在精细定位BVDV株的进化关系中具有显著优势。
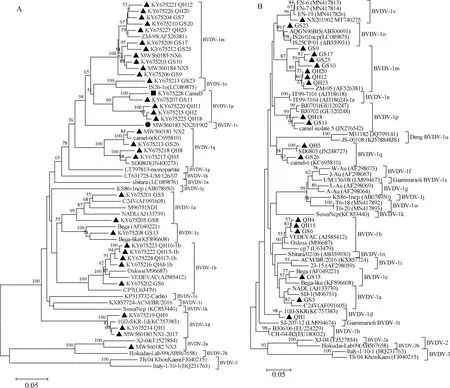
图2 基于Erns基因(A)和5′UTR核苷酸(B)的各亚型BVDV株系统进化分析
3 讨 论
自1946年美国首次报道牛病毒性腹泻,至今已有70余年。BVDV以牛为主要宿主,还可感染羊、猪、牦牛、鹿等家畜和多种野生偶蹄动物[15]。BVDV-1基因型在世界各地普遍流行,而BVDV-2 基因型在北美、南美、欧洲、亚洲流行,目前已发现BVDV-1a~1w、BVDV-2a~2d等基因亚型。BVDV-3基因型主要分布于巴西、泰国、意大利、孟加拉国、印度、阿根廷、土耳其等国家。相关研究表明,在我国牛BVDV总体抗体阳性率高达57%,RT-PCR阳性率约为27.1%[16],但病毒抗原阳性率较低(0.71%~3.66%)[17-18],流行株主要有BVDV-1a~1d、BVDV-1m~1q、BVDV-1u~1w、BVDV-2a~2b等亚型[6-12]。此外,在河南、山东省相继报道山羊、绵羊以及肉牛因BVDV-3自然感染发病[19-20]。本研究利用疑似BVDV感染牛抗凝血样本研究不同亚型株Erns抗原基因的差异,RT-PCR总体阳性率(37.33%)和抗原阳性率略高于前期普查性研究。作者发现在西北地区1a、1b、1d、1q亚型BVDV流行态势与前期报道类似[21-23],但BVDV-1m亚型株目前流行最普遍。BVDV-1p基因亚型株最早于2007年分离于北京奶牛[7],在我国江苏和西南地区山羊以及西北区双峰驼均有报道[24-26],在本研究中发现其在西北省区其检出率与1q株相当,仅次于BVDV-1m株。在宁夏首次检出BVDV-2a基因型,但其与我国前期分离株的亲缘关系相对较远,与之类似近期北美商品血清来源的BVDV-2分离株的报道[27],提示国外传入BVDV-2株的风险不容忽视。
目前,BVDV分离株的亚型鉴定大多基于5′UTR、N基因核苷酸相似性,由于不同亚型株的基因重组可能造成BVDV株遗传分析的偏差[28]。因此日本学者曾增加E2 N端编码区、NS3基因片段、NS5B-3′UTR 的系统进化分析以提高准确性[29]。抗原编码基因在BVDV株间变异最大,在揭示病毒遗传演化规律中更具优势。与作者前期发表的BVDV流行株E2基因的分析结果类似[13],本研究发现Erns基因也可作为研究BVDV遗传演化的靶标基因,精细定位BVDV株进化关系。此外,我们发现RNA酶活性位点在BVDV不同亚型株高度保守,但1m~1q、1v等新亚型株Erns糖基化位点错位,由于ErnsRNA酶活性位点、dsRNA结合活性位点、糖基化在影响病毒抗原性以及宿主细胞INF产生中具有重要调控作用[30-31],国内流行株Erns糖基化位点错位是否影响毒株抗原性或宿主细胞IFN反应的差异,还有待深入研究。
近期研究人员提出了1f、1g、1h、1m~1o、1q亚型株进化的关联性[32],本研究聚焦于国内流行株,首次全面获取了BVDV Erns编码基因信息,通过同源性分析进一步揭示了1m~1v亚型BVDV流行株的密切亲缘关系。前期研究人员在山东奶牛[6]和黑龙江肉牛[33]报道了BVDV新亚型1v,本研究进一步发现在宁夏区奶牛中也存在同亚型流行株,这一结果提示1v亚型株在我国的时空分布规律及其在我国BVDV株演化中扮演的角色值得关注。
4 结 论
首次选用Erns靶标基因对西北部分省区牛源BVDV株进行同源性及系统进化分析,发现10个基因亚型流行株,以1m亚型株最为普遍,1m~1q及1v等亚型BVDV株亲缘关系密切。
