锦鲤与草金鱼杂交子代肠道形态和酶学特性
2021-11-23刘君恒冯晓飒高建忠
刘君恒,冯晓飒,张 源,高建忠
( 1.上海海洋大学,水产科学国家级实验教学示范中心,上海 201306; 2.上海海洋大学,上海水产养殖工程技术研究中心,上海 201306; 3.上海海洋大学,农业农村部淡水水产种质资源重点实验室,上海 201306 )
鱼类是现存脊椎动物中最大的类群,距今已有6亿多年的存在历史,它们代表了广泛的生理学、生态学和自然历史[1-3]。杂交作为物种强化和基因重组的主要驱动力,在物种历史演变中发挥重要作用[4]。近50年来,由于过度捕捞,现有鱼类已无法满足人类对水产品的需求,杂交被广泛应用于水产养殖领域中[5]。但早期人们对于鱼类杂交研究主要集中在杂种优势的应用上,对于相关基础遗传学的报道较少[6]。
研究发现,不同鱼类因为食性和摄食习惯的差异会导致其消化道在结构和功能上有所不同[7-8]。如草鱼(Ctenopharyngodonidellus)♀和鳙鱼(Aristichthysnobilis)♂杂交F1代在食性上更接近于草鱼[9];Li等[5]在对团头鲂(Megalobramaamblycephala)和翘嘴鲌(Culteralburnus)进行正、反杂交后发现,所获得的两种杂交后代在肠道形态上都更接近于团头鲂。
锦鲤(Cyprinuscarpio)和草金鱼(Carassiusauratus)是重要观赏鱼类,深受广大水族爱好者的喜爱。为选育观赏价值更高的长尾锦鲤,笔者进行了锦鲤与长尾草金鱼的杂交。同时发现,锦鲤与草金鱼虽同为杂食性鱼类,但锦鲤肠道粗大,草金鱼肠道细长,且排布状态存在差异,而锦鲤与草金鱼杂交子代的肠道性状也不同于双亲,推测二者杂交子代肠道遗传特性可能介于锦鲤与草金鱼之间。为查明锦鲤和草金鱼杂交子代的肠道参数和功能特征,笔者选取昭和锦鲤与长尾草金鱼作为亲本,进行自交及正、反交,并对子代的肠道参数、组织形态、pH环境及消化酶活性进行比较分析,以期为锦鲤与草金鱼杂交育种和饲养提供理论基础和科学参考。
1 材料与方法
1.1 试验材料
1.1.1 亲本来源及苗种生产
亲本昭和锦鲤和长尾草金鱼均来自上海海洋大学观赏水族良种培育与健康养殖实验室。于2019年5月8日挑选品相良好、体质健壮的亲本进行人工注射催产。亲本于5月9日开始产卵受精,后经自然孵化得到昭和锦鲤自交F1(昭和组)、长尾草金鱼自交F1(草金组)、正交F1(长尾草金鱼♀×昭和锦鲤♂,正交组)和反交F1(昭和锦鲤♀×长尾草金鱼♂,反交组)。
1.1.2 养殖管理
分别选取昭和组、草金组、正交组和反交组各150尾试验鱼,每组分成6个平行,每个平行25尾,进行分缸饲养。试验过程中养殖用水为曝气后的自来水,溶解氧>5.0 mg/L,水温25~28 ℃,pH 7.1。各组鱼苗开口饵料为卤虫无节幼体。待试验鱼转食颗粒饲料后移入5 m×3 m水泥池中,按照鱼体质量的3%日投喂3次(8:00、13:00、18:00),喂养周期为45 d。
1.2 样品采集
养殖试验结束时,将试验鱼进行8 h饥饿处理,每组鱼随机选取10尾用间氨基苯甲酸乙酯甲磺酸盐(100 mg/L)麻醉后,用游标卡尺和天平测量鱼体长(L, mm)、体质量(m, g)后在冰面上进行解剖,取鱼体整个肠道,测量其肠长(LG, mm)、肠质量(mG, g)。用解剖刀轻轻刮取鱼体前肠、中肠和后肠内容物1 g置于无菌离心管,用于测量肠道pH;另取肠壁用0.68%无菌生理盐水冲洗后置于无菌离心管内,保存于-80 ℃用于消化酶活性测定;再分别选取10尾鱼,将肠道按折点划分为前肠、中肠和后肠,置于波恩氏液中固定[10],用于肠道切片组织形态测定。
1.3 样品处理
肠道pH测定:将肠道内容物解冻后,参照文献[11]的方法测定。肠道蛋白酶、脂肪酶和淀粉酶活性参照南京建成生物工程研究所的试剂盒说明书进行测定[12]。将肠道组织从波恩氏液中取出用75%乙醇洗涤后,经脱水、包埋(取各肠段中间1 cm)、切片和苏木精—伊红染色等步骤进行观察测量。
1.4 数据统计计算及分析
RGL=LG/L
(1)
RGD=LG/m
(2)
wGM=mG/m
(3)
IZ=LG/[10×(m1/3)][13]
(4)
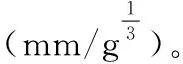
切片形态数据采用Image J软件进行测量,所测得试验数据采用SPSS 20.0软件进行单因素方差分析,并用Turkey方法进行多重比较(显著性水平为0.05)。试验统计分析结果用平均值±标准差表示。
2 结果与分析
2.1 昭和锦鲤与长尾草金鱼自交及正、反交F1肠道参数比较
昭和组和正交组的肠质量差异不显著(P>0.05),草金组和反交组的肠质量差异不显著(P>0.05),但昭和组和正交组的肠质量显著高于草金组和反交组的肠质量(P<0.05)(表1)。昭和组和反交组的肠长差异不显著(P>0.05),但均显著低于草金组和正交组的肠长(P<0.05)。昭和组与草金组的相对肠长、Zihler′s指数以及相对肠密度差异显著(P<0.05),但正交组和反交组之间差异不显著(P>0.05),介于两自交组之间。昭和组与两杂交组的相对肠质量差异不显著(P>0.05),但草金组的相对肠质量为0.12,显著高于这3个试验组。
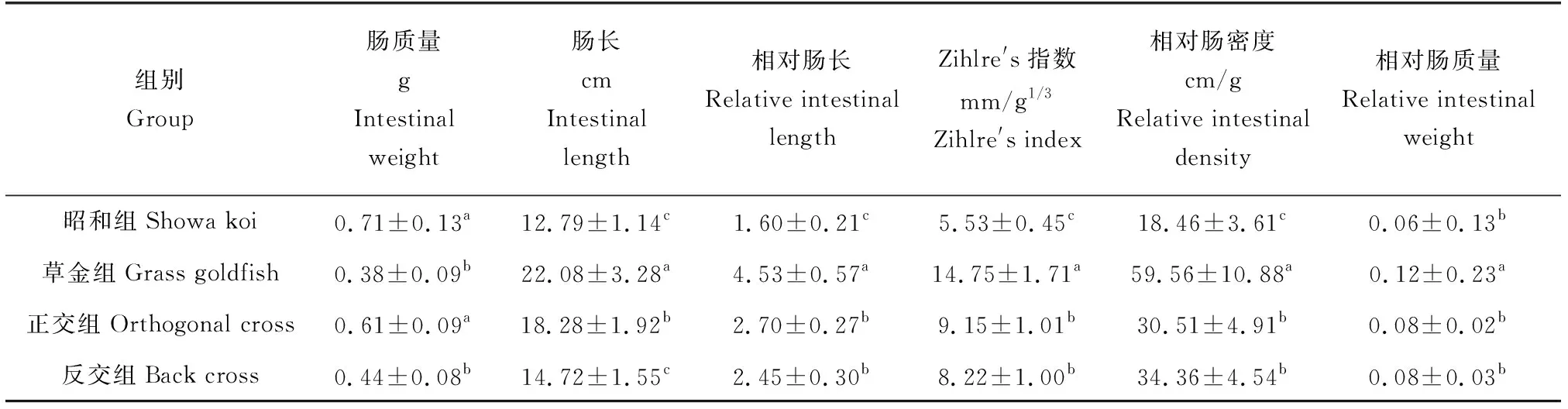
表1 昭和锦鲤与长尾草金鱼自交及正、反交F1肠道参数
2.2 昭和锦鲤与长尾草金鱼自交及正、反交F1肠道组织形态比较
4组鱼的前中后肠绒毛高度均存在显著性差异(P<0.05),前中后肠绒毛高度大小均为:昭和组>正交组>反交组>草金组(表2)。草金组与正、反交组的前肠隐窝深度差异不显著(P>0.05),但昭和组的前肠隐窝深度显著高于其他3组(P<0.05)。4组鱼的中、后肠隐窝深度存在显著性差异(P<0.05),中肠隐窝深度大小为:草金组>昭和组>反交组>正交组,后肠隐窝深度大小为:昭和组>反交组>草金组>正交组。正交组和反交组的前肠肌层厚度在两自交组之间无显著性差异(P>0.05),且昭和组的前肠肌层厚度显著高于草金组(P<0.05)。但草金组与反交组的中肠肌层厚度无显著性差异(P>0.05),与昭和组、正交组的中肠肌层厚度有显著性差异(P<0.05),且显著低于昭和组、正交组的中肠肌层厚度。4组鱼的前中后肠的黏膜厚度均存在显著性差异(P<0.05),杂交组在两自交组之间,昭和组显著高于草金组(P<0.05)。昭和组和正交组的前中肠绒毛高度/隐窝深度无显著性差异(P>0.05),且显著高于其他两组的前中肠绒毛高度/隐窝深度(P<0.05)。4组鱼的后肠绒毛高度/隐窝深度存在显著性差异(P<0.05),大小依次为:昭和组>正交组>反交组>草金组。

表2 昭和锦鲤与长尾草金鱼自交及正、反交F1肠道组织形态
2.3 昭和锦鲤与长尾草金鱼自交及正、反交F1肠道pH比较
正交组和反交组的前肠pH无显著性差异(P>0.05),但与两个亲本之间存在显著性差异(P<0.05),正反交的前肠pH低于草金组pH,而高于昭和组pH(表3)。4组鱼的中后肠pH均不存在显著性差异(P>0.05)。

表3 昭和锦鲤与长尾草金鱼自交及正、反交F1肠道pH
2.4 昭和锦鲤与长尾草金鱼自交及正、反交F1肠道消化酶活比较
4组试验鱼的淀粉酶差异显著(P<0.05),且正交组淀粉酶活性为107.43 U/mg,显著高于昭和组和反交组(图1a)。昭和组和草金组的脂肪酶活性差异不显著(P>0.05),但显著低于正交组和反交组的脂肪酶活性,且反交组脂肪酶活性最高(图1b)。正交组和反交组的蛋白酶活性无显著性差异(P>0.05),介于两自交组之间,但昭和组的蛋白酶活性显著低于草金组(P<0.05)(图1c)。
3 讨 论
3.1 昭和锦鲤与长尾草金鱼自交、正反交F1肠道参数
消化道作为鱼类食物消化、吸收的主要场所,其承载鱼类生命活动的基础[14]。研究发现,不同鱼类因为食性和摄食习惯的差异会导致其消化道在结构和功能上有所不同[10,15],这也使得人们在鱼类育种工作中对于亲本和子代之间的遗传差异有更好的了解。因此对鱼肠道指数的研究,有助于人们更好地研究鱼类摄食习性和健康状况。肠长指数和Zihlre′s指数常被用来反映鱼类的食性偏向性,对肠质量和密度的研究,进一步反映了鱼类对食物的消化能力。本试验结果显示,昭和锦鲤与长尾草金鱼虽同为杂食性鱼类,但长尾草金鱼的相对肠长、Zihlre′s指数、相对肠密度、相对肠质量均大于昭和锦鲤,说明长尾草金鱼为杂食偏草食性鱼类。通过杂交得到的后代,其肠道指标与亲本相比均发生改变,且介于两亲本之间,这说明杂交后代的形态结构受亲本遗传因素的影响,这与团头鲂和翘嘴鲌杂交及杂交鲟(Husohuso♀×A.schrenscki♂)的试验中得出的结论相符[5,16]。本试验结果显示,杂交组鱼肠道长度和质量更偏向于各自的父本,但肠道指数之间并没有显示出这种偏向性。
3.2 昭和锦鲤与长尾草金鱼自交及正、反交F1肠道组织形态
对肠道组织观察可进一步了解鱼类对食物的消化吸收能力,反映动物肠道消化功能的重要指标有绒毛高度、隐窝深度、绒毛高度/隐窝深度等[17-18]。其中肠绒毛对营养物质消化吸收起重要作用,绒毛越长,其吸收部位面积越大,消化能力越强。隐窝深度是反映肠细胞分化绒毛的能力,肠道绒毛高度越高、隐窝深度越浅表示绒毛高度/隐窝深度越高,其消化吸收功能越强[16]。黏膜层有很多皱褶,这些皱褶可增大肠壁面积,肠黏膜的正常结构与功能是营养物质充分消化与吸收的基本保证[19]。肌层厚度变化对营养物质的吸收和转运效率有重要的影响[20]。鱼类肠道根据肠道在体内粗细变化及折转方向分成前、中、后3段[21],前肠主要与消化吸收相关,中肠和后肠主要与免疫相关[22]。本试验中,正交组的绒毛高度、肌层厚度和黏膜厚度3个指标均高于长尾草金鱼组,且4组子代前肠绒毛高度均明显高于中、后肠。前肠隐窝深度依次为正交组、长尾草金鱼自交组、反交组、昭和锦鲤自交组。前、中肠的绒毛高度/隐窝深度正交组高于其他3组,且4组鱼的前肠绒毛高度/隐窝深度高于中、后肠;而反交组的各项指标均介于两个亲本之间。通过以上结果分析可知,正交组肠道绒毛密集、肌层厚实,收缩力强,对营养物质消化能力强,具杂种优势。从肠道健康方面考虑,应选择昭和锦鲤作为父本。这与徐莹等[23-24]所观察的草鱼♀×赤眼鳟(Squaliobarbuscurriculus)♂杂交F1肠道组织形态的结果相似。
3.3 昭和锦鲤与长尾草金鱼自交及正、反交F1肠道pH及消化酶活性
鱼类的消化酶活性是反映鱼类消化生理机能的一项重要指标[8],消化器官的组织结构和消化机能是与其食性相适应的,消化器官的组织结构不同其消化机能不同,消化酶的活性也有明显差异[25]。本试验正、反交子代肠道淀粉酶、脂肪酶活性均大于亲本,脂肪酶、淀粉酶活性反映鱼类对能量的利用情况[26],表明杂交子代对饲料中脂肪和淀粉等营养物质的利用率可能更高。长尾草金鱼蛋白酶活性大于昭和锦鲤。鱼类肠道(尤其是前肠)是鱼类消化吸收的主要场所[27],肠道蛋白酶的最适pH为8.0~10.0,甚至高达11.0[28-31]。本试验中,子代蛋白酶活性和前肠pH大小均为:长尾草金鱼自交组>正、反交组>昭和锦鲤自交组。较低的昭和锦鲤蛋白酶活性可能是因为前肠pH影响了蛋白酶活性[32-33]。肠道内pH的改变会影响消化酶活性,进而影响到杂交鱼的生长和发育[32-33],为保证食物的消化吸收,可以在其饲料中添加一定的酶制剂,提高饲料利用率。
4 结 论
昭和锦鲤与长尾草金鱼正、反交子代肠道特性高于或介于二者自交子代之间,综合了二者的肠道性状。
