不同密度下入侵植物北美车前生物量分配与异速生长关系
2021-11-22张世航戈玉莹1志1江浩然刘纪元冶1
张世航,龚 莉,戈玉莹1,,洪 志1,,江浩然,刘纪元,陶 冶1,
(1. 中国科学院新疆生态与地理研究所 荒漠与绿洲生态国家重点实验室,新疆 乌鲁木齐 830011;2. 中国科学院大学,北京 100049;3. 安庆师范大学 生命科学学院 / 皖西南生物多样性研究与生态保护安徽省重点实验室,安徽 安庆 246133)
生物入侵常常导致一系列负面影响,如物种快速丧失、巨大经济损失、生态效益急剧降低,以及不可逆的环境破坏等[1]。外来入侵植物侵占本地植物生长所必要的资源(如水资源),局部改变了本地种原有的空间分布格局,甚至造成了一些土著种濒临灭绝,严重影响生物多样性和生态安全[2]。北美车前(Plantago virginica)是一种恶性入侵植物,原产北美地区,1951 年始见于江西南昌市莲塘区,目前在淮河以南十余个省份均有发现,正迅速扩散[3-4]。该植物种子多,繁衍力强,扩张迅速,常常入侵和危害草地生态系统,在荒山和草坡等生境也有大量分布。与三裂叶豚草(Ambrosia trifida)类似,该植物也可能造成人体花粉过敏症[5]。北美车前种群在入侵地急剧增长,反映出较强的环境适应能力,具体体现在生理、形态和生物量分配的可塑性等方面[6]。因此,探讨北美车前种群密度、繁殖特征及生物量分配之间的关系,可以在一定程度上揭示该物种的入侵繁衍机制[7]。
生物量是研究植物生物学特征和功能性状的重要基础,是能量积累的基本表现,是研究生态系统中物质循环和能量流动的重要指标[8-9]。植物不同的生物量分配策略反映了植物在不同选择压力(如自然环境梯度、人类扰动及生态系统管理等) 下的生活史对策,通常最优分配假说认为植物会分配更多生物量给获取资源最受限制的器官[10-11],但植物依然会根据实际需要进行资源再分配,进而实现生物量分配策略调整[12]。生物量分配尤其是繁殖分配的大小对于入侵植物是否能产生更多种子、呈现更强的入侵能力,具有极为重要的作用[6-7]。当然,繁殖分配的前提是要有足量的叶片(营养分配)以实现繁殖生物量的增加。因此,光合(或营养)生物量与繁殖生物量的分配权衡,是植物尤其是入侵植物提升入侵能力的关键环节之一[1,13]。植物光合作用的产物在地上和地下各器官中的资源分配通常有较大差异,这与植物的个体发育、生长环境、植物种类(功能群)和群体大小相关[12,14-15]。异速生长关系是指生物体形态指标、生物量等不呈比例的生长关系,反映植物个体适应环境中各器官之间存在的权衡策略[16],这种现象表现出植物每个功能器官或构件生长的异步性[17]。通过异速生长分析可以深入了解不同功能群植物的表型可塑性,揭示植物生物学特征间的与尺度无关的潜在规律[18-19],如新陈代谢率和生物个体大小表现的是幂指数关系,而地上与地下生物量之间多为等速分配关系[17]。
种群密度是自然界重要的选择压力,密度制约则是植物种群普遍存在的特征之一,是种群调节的重要机制[20-21]。植物密度变化受到个体间强烈竞争的影响,竞争强度随植物发育阶段的变化而变化,而种群密度也随竞争强度的变化而变化[22]。密度制约对个体生长的调节是一个动态过程,从种子萌发到植株凋亡均有所体现,且草本植物往往比木本植物对密度压力更敏感[23]。密度引起的植株邻近体间遮荫效应将影响光照强度、光照时间和光质,促使植物调整资源利用和分配策略[24]。在密度压力下,植株会显著改变自身的株型发育系统,并影响各器官间的异速生长,进而权衡器官生物量分配以完成生活史[25]。除此以外,密度效应还能影响植物的繁育系统,如垂穗披碱草(Elymus nutans)能够进行有性繁殖的个体比例随密度的增加而下降[26]。因此,探讨不同密度下植物生物量分配策略,对于揭示不同类型植物的环境适应性和生存策略具有重要意义。
对北美车前而言,在繁殖阶段其地上、地下部分形态参数及繁殖分配均受种群密度的制约[27]。与低密度种群相比,高密度北美车前植株过氧化物酶活性、可溶性糖及脯氨酸含量较高叶绿素a/b 则较低,使高密度种群可较好地适应郁闭群体环境,并通过抗氧化酶的调节形成一定的抗逆能力[28]。北美车前形态可塑性强,其个体形态学指标随种群密度增加呈现规律性变化,但形态、生理和生殖分配等特征对种群密度表现出不同程度的表型可塑性[29]。不管如何,更多的繁殖输出是北美车前表型可塑性的最终目标,以达到定植、生长和扩散的目的,如入侵地北美车前果序生物量可达原产地的3 倍多[30],但其不同器官间资源分配的密度调节机制还有必要深入探究。
长江中下游地区是我国南北气候(即温带和亚热带)过渡区,也是生态系统类型(常绿阔叶林和落叶阔叶林) 和农业耕作方式(水田和旱田) 的过渡区,具有重要的生态价值[31]。然而,该地区也是北美车前入侵最早且态势极为严重的区域,其种群扩散迅速且具有明显的伴人特征,弃耕地、路边、村舍周边等地是其扩张的重灾区,且正逐步扩散到其他生境,对各类陆地生态系统已产生严重的直接或间接威胁[3-4]。尽管早期已对北美车前密度制约效应开展了一些研究,但其一方面关注于形态特征而忽视了资源分配的变化和响应,另一方面关注于上述指标之间的线性关系而忽视了异速生长关系的重要性,且缺乏明确的密度梯度及对比分析[7,29]。因此,从个体水平上探究不同种群密度下北美车前生物量(或资源)分配特征及不同器官间的异速生长关系具有一定的必要性,为深入解析北美车前入侵机制和表型可塑性提供科学依据。
1 研究地区与研究方法
1.1 研究区概况
采样地区位于安庆市宜秀区待建撂荒地(116°59′ E,30°36′ N)。安庆市地处长江中下游、皖鄂赣三省交界处,是安徽省西南部重要城市,属于温带与亚热带的过渡地带,平均气温14.5~16.6 ℃,年降水量1 300~1 500 mm,年日照时数2 012 h,四季分明,雨量充沛,光照充足,年无霜期247 d。该地区地貌多样,襟江带淮,山地、丘陵、河湖面积各占1/3。采样地的地被植物以北美车前为主,其他植物种类较少且组成相近,伴生植物有野胡萝卜(Daucus carota)、狗尾草(Setaria viridis)、野豌豆(Vicia sepium)和一年蓬(Erigeron annuus)等,也有一些藓类植物分布[32]。研究区土壤为黄棕壤、黄褐土;经测定,采样区土壤有机质、全氮和全磷含量分别为26.12、1.61 和0.81 g·kg−1,速效氮和速效磷含量分别为85.98 和26.48 mg·kg−1,土壤肥力一般;土壤pH 7.9,稍偏碱性。
1.2 研究方法
1.2.1 野外样品采集与预处理
植物样品于2019 年5 月中旬植物开花结果期采集。依据前期实地调查,设定3 个密度梯度开展调查研究,分别为30 株·m−2(低密度)、100 株·m−2(中密度) 和220 株·m−2(高密度),3 个不同密度种群环境背景相同。3 种密度梯度分别调查9、13 和7 个样方(共29 个样方),样方大小均为1 m × 1 m,样方间隔5 m以上。详细计数样方内北美车前株数后,按照对角线法在每个样方中选取10 株有花有果植株,采用全株挖掘法获取整株,挖掘深度 > 20 cm,以保证根系完整。低、中、高密度分别有90、130 和70 株植物。
将植物样品带回实验室清洗干净,用滤纸快速擦干后测量相关形态指标;随后将每株植物分为繁殖器官(花序 + 花序轴)、叶片和根3 部分,置于70 ℃烘箱内烘干至恒重后用天平(精度均为0.000 1 g)测定干重。
1.2.2 生物量指标计算
花序轴和花序生物量之和为繁殖生物量(reproductive biomass,RB),每株植物的叶生物量(leaf biomass,LB)和繁殖生物量之和为地上生物量(above ground biomass,AGB),根 生 物 量 为 地 下 生 物量(underground biomass,BGB),地上与地下生物量之和为总生物量(total biomass,TB)。计数每株总叶片数,基于每株LB 计算单叶重(single leaf biomass, SLB)。计算根冠比(root to shoot biomass ratio,R/S) 及繁殖叶片生物量比(reproductive tissue to leaf ratio,R/L),以反映不同密度条件下地上与地下之间及繁殖器官与光合器官生物量分配的权衡特征。
1.2.3 统计分析方法
先对3 种不同密度条件下北美车前的LB、RB、AGB、BGB、TB 以及R/S、R/L 等指标进行描述统计分析,随后进行One-Way ANOVA 和回归分析,用Duncan 法进行多重比较(α= 0.05)。生物量指标之间的异速生长关系通过幂函数表示。

式中:Y或X为北美车前器官生物量,b为标准化常数,k为异速生长指数。k= 1 为等速关系,k≠ 1 为异速关系,分别表示Y和X间为等比例或不等比例变化。通常先将幂函数对数化,利用降主轴回归(RMA,即Model Type Ⅱ) 计算异速生长指数(a,即线型回归的斜率)、截距(logb)、95% 置信区间(95%CI)及决定系数(R2)[33]。依据异速生长指数和截距的差异性,植物不同群体间的异速生长关系的差异性可分为以下几种类型[34-36]:A 型漂移(a差异显著)、B 型漂移(a相同,截距差异显著但具有共同拟合轴,称为截距漂移)、C 型漂移(a相同,截距相同但拟合轴发生漂移,称为共轴漂移) 和D 型漂移(B 和C 同时发生),而如果斜率和截距相同且共轴,表明这些类群的相关指标间具有完全相同的异速生长关系[37-38]。
异速生长分析使用SMATR 软件完成,数据处理和作图使用Excel 2010 完成,方差分析使用SPSS 20.0 软件实现。
2 结果与分析
2.1 不同密度下北美车前各器官生物量的差异性
不同密度梯度下北美车前大部分器官生物量均有较强的变异性,其中低密度下北美车前各器官生物量的变异系数为0.709~0.964,中等密度下变异系数为0.201~0.918,高密度下变异系数为0.154~0.850,中、高密度下繁殖生物量变异程度最小(表1)。低密度下单株地上生物量为0.026~0.742 g,平均为0.243 g;单株地下生物量为0.017~0.130 g,平均为0.028 g;全株总生物量为0.027~0.872 g,平均为0.270 g。中密度下单株地上生物量为0.032~0.315 g,平均为0.166 g;单株地下生物量为0.002~0.091 g,平均为0.016 g;全株总生物量为0.033~0.406 g,平均为0.182 g。高密度下单株地上生物量为0.013~0.254 g,平均为0.066 g;单株地下生物量为0.001~0.026 g,平均为0.005 g;全株总生物量为0.014~0.280 g,平均为0.071 g。无论各器官生物量还是全株生物量,随密度增大均显著降低(P< 0.05),其中高密度下的个体大小(总生物量) 仅为低密度的26.2%。除此以外,单叶重也具有显著密度差异性(P< 0.05),低、中和高密度下分别为每片叶重为31.75、18.78 和8.95 mg,也呈递减趋势(图1)。
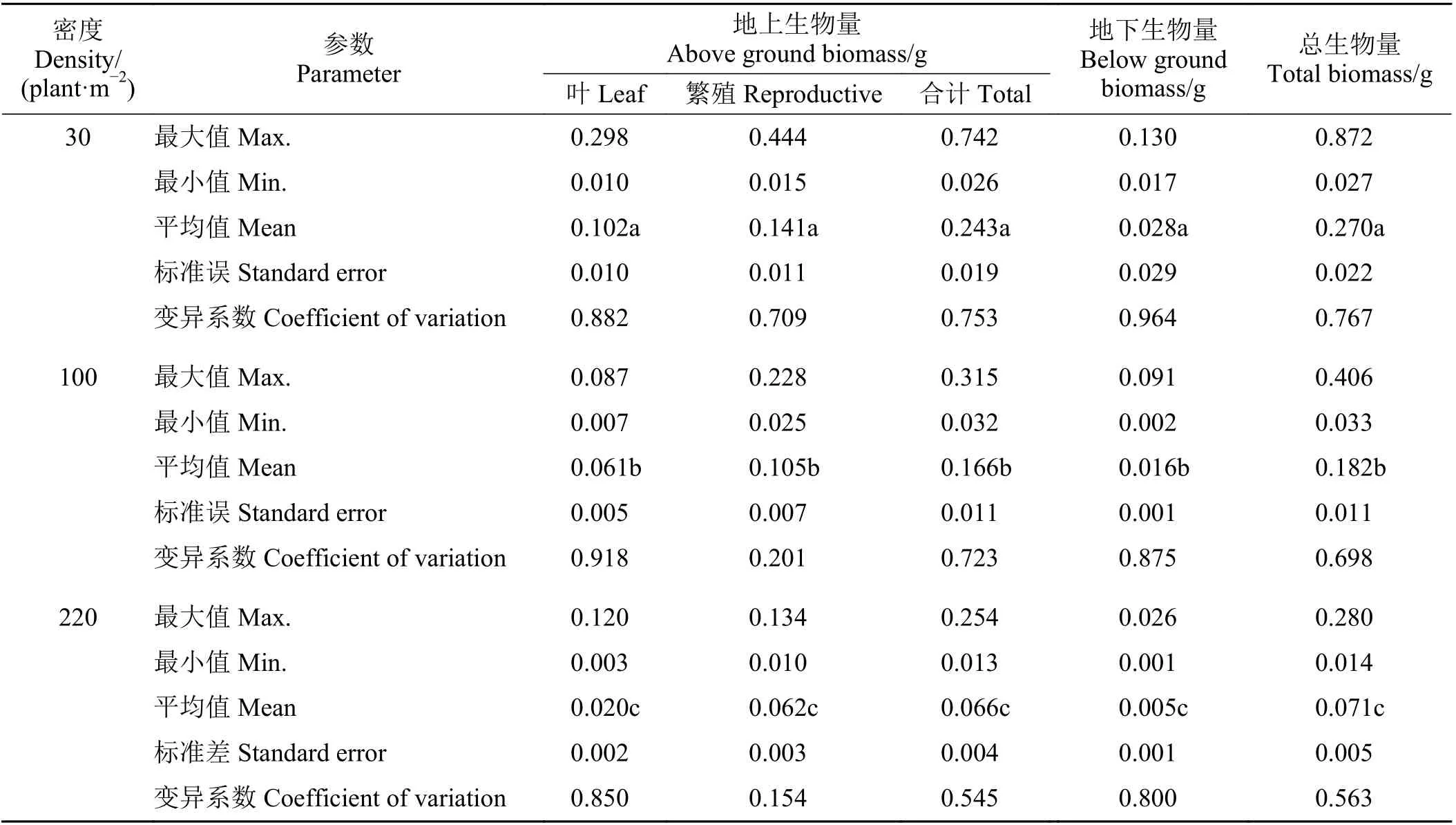
表1 不同密度下北美车前不同器官生物量的统计Table 1 Description of biomass of different Plantago virginica organs at different densities

图1 不同密度下北美车单叶重的差异性Figure 1 Effect of plant density on single leaf biomass of Plantago virginica
2.2 不同密度下北美车前生物量分配比例的差异性
3 种密度下北美车前的繁殖器官、叶及根3 个部分的分配比例均具有显著差异(P< 0.05),其中根和叶生物量分配比例随密度增大显著降低,而繁殖生物量比例则呈显著增加的趋势,低、中和高密度下分别为53.9%、58.6%和66.2% (图2)。
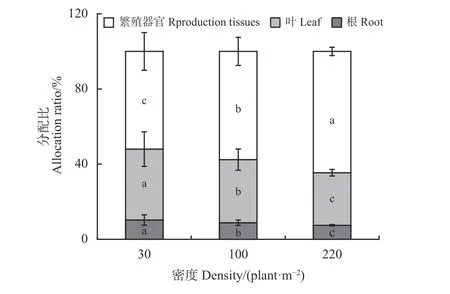
图2 不同密度下北美车根、叶、繁殖器官生物量比例的差异性Figure 2 Differences in biomass allocation ratios of root, leaf,and reproductive tissues of Plantago virginica grown in different density areas
根冠比(R/S)在低密度(0.101)与中等密度(0.104)间无明显差异,但均显著高于高密度(0.079) (P<0.05),表明高密度条件下北美车前生物量向根部分配的比例受到较强抑制(图3)。尽管低密度条件下的个体更大,但其繁殖叶片比(R/L) 仅为1.677,显著低于中密度(2.045)和高密度(3.166) (P< 0.05),表明随植株密度增加,在地上光合器官和繁殖器官生物量分配的权衡上,北美车前更倾向于增大繁殖器官的资源投资而减小光合器官资源分配(图3)。

图3 不同密度下北美车前根冠比(R/S)及繁殖叶片比(R/L)的差异性Figure 3 Differences in root to shoot (R/S) and reproductive tissue to leaf (R/L) ratios in Plantago virginica grown at different plant densities
2.3 不同密度下北美车前器官生物量间的异速生长关系
3 种密度下北美车前各器官生物量之间均具有显著的异速生长关系(P< 0.05) (表2),6 对器官生长关系中均有2 对符合等速关系。4 个异速生长关系中,不同密度下仅有3 对相同,分别为地上-地下(均指X-Y,下同)、地上-叶片及地下-繁殖生物量;而这3 个异速生长关系中,前两个异速生长指数均大于1.0,第3 个异速生长指数小于1.0,表明地下生物量和叶片生物量分配速率高于地上生物量,而繁殖生物量分配速率低于地下生物量。两对等速关系中,不同密度下仅地上-繁殖生物量是相同的,表明二者生物量分配速率相同。地下生物量与叶生物量间的异速生长指数在不同密度下有显著差异,其中低密度下异速生长指数最小且呈小于1.0 的异速生长关系,而中、高密度下二者呈等速生长关系,表明密度升高显著增加了北美车前地上光合器官的生物量分配速率。此外,在地上光合器官(叶)与繁殖器官之间,低密度下呈等速关系,而中、高密度下则呈小于1.0 的异速关系,表明地上部分的光合与繁殖资源分配受到了明显的密度制约,密度升高北美车前会提高光合器官的生物量分配速率并降低繁殖生物量分配速率(非绝对量)。
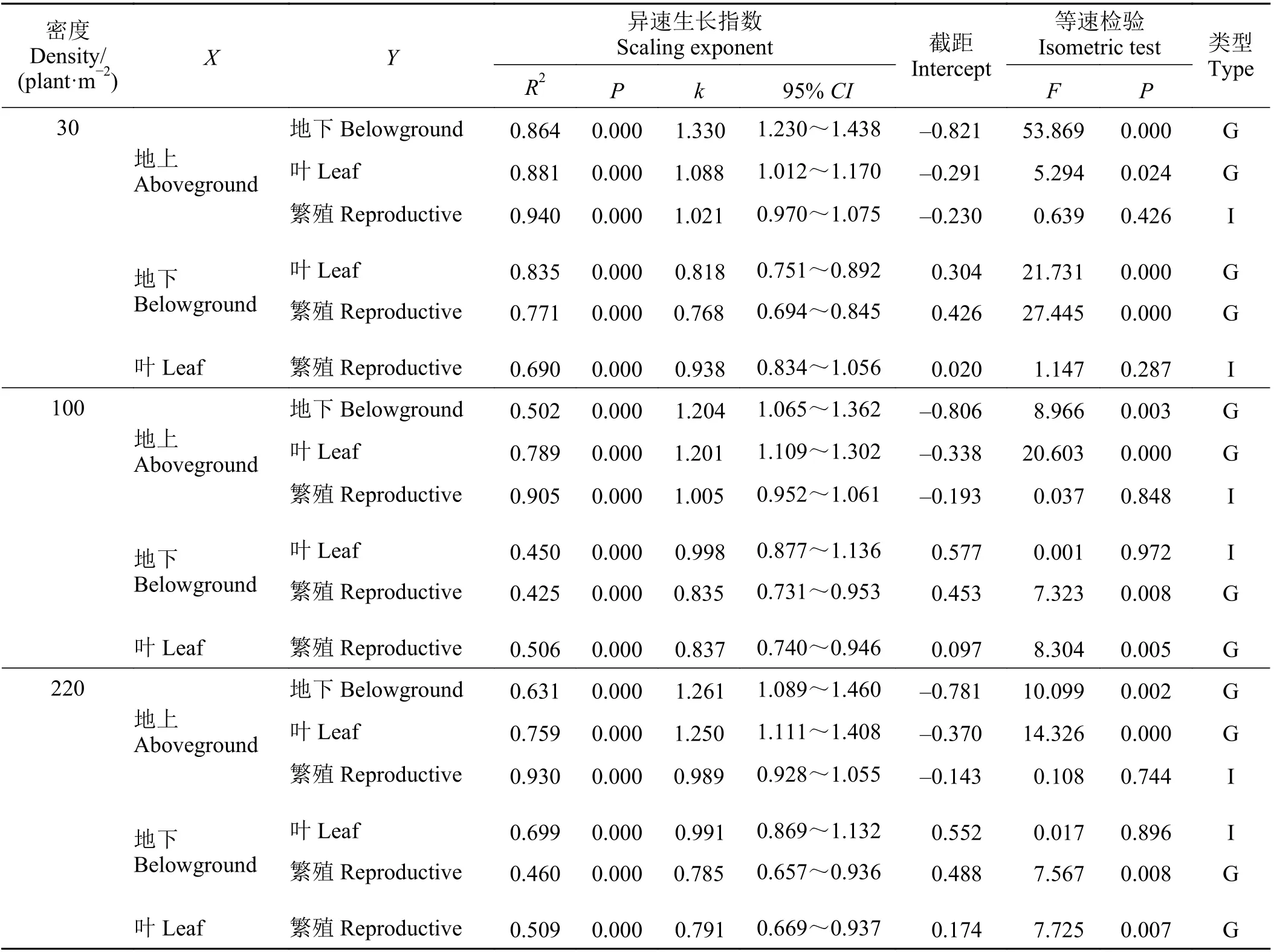
表2 不同密度北美车前各器官生物量间的异速生长指数及等速检验Table 2 Allometric scaling exponent and isometric test among organ biomasses of Plantago virginica grown at different densities
进一步对不同密度下北美车前器官生物量间的异速生长关系进行共同斜率检验和漂移类型分析发现,地下-叶生物量之间异速生长指数差异显著,即没有共同的斜率,属于A 型。地上-地下及地下-繁殖生物量间具有共同斜率和截距,但发生了共轴漂移,属于C 型。其他3 对(即地上-叶、地上-繁殖及叶-繁殖生物量)属于B 和C 同时发生的情形,即属于D 型(表3)。

表3 不同密度北美车前各器官生物量间共同斜率检验及异速生长关系漂移类型Table 3 Test for common slope, and type of shift in allometric relationship among Plantago virginica organ biomasses from plants grown at different densities
3 讨论
3.1 密度对北美车前生物量分配的影响
本研究中,北美车前各器官生物量、单叶生物量及单株总生物量均随密度升高而显著降低,表明高密度压力下即使是入侵植物,其个体大小也会受制于密度带来的竞争压力,即呈现出“竞争-密度效应”[39]。对于无性系繁殖的羊草(Leymus chinensis)而言,随着密度的增加,其在种间竞争中采取优先延长根茎长度,然后再增加分株数量和地下生物量的繁殖适应对策[40]。对四季竹(Oligostachyum lubricum)而言,随着密度增大,1~3 年生立竹地上生物量、构件生物量均呈先升高后显著降低的变化趋势;而随密度增大,枝、叶生物量分配比例呈降低趋势,秆生物量分配比例呈升高趋势,表明高密度下四季竹增加主要支撑构件秆的高度,以获得更多光资源[41]。随密度增大,沙生植物猪毛菜(Salsola collina)株高、根长、一级分枝数、二级分枝数、三级分枝数及总分枝长均呈减少趋势[42]。羌塘雪兔子(Saussurea wellbyi)随海拔升高其植株密度、个体大小、繁殖生物量及管状花数量均逐渐降低,但其管状花重量及繁殖分配比例却显著升高[43]。大果虫实(Corispermum macrocarpum)株高呈减小趋势,其分枝数及分枝长度明显减小,各器官生物量随密度增大而显著减小,该结果与本研究相同[21];但随密度增大,大果虫实茎和繁殖器官生物量分配呈减小趋势,根和叶片生物量分配呈增大趋势[25],而沙蓬(Agriophyllum squarrosum)的繁殖分配比例无显著变化,但根和茎生物量分配增加[23]。这与本研究结果不同,即北美车前叶、根的分配比例及根冠比均随密度增大而降低,而繁殖分配比例及繁殖叶片比随密度增大呈显著增加趋势,表明高种内竞争压力下北美车前优先将资源分配给繁殖器官,以保证后代繁育。此外,在竞争条件下,植物形态的改变往往比资源分配的调整更快[17],但本研究未涉及形态指标,因而尚不能证实北美车前是否具有相同特征。
密度主要通过影响邻近植物间器官对资源的竞争强度来调节资源分配。密度变化会导致植物种内对空间与资源发生竞争,进而导致各器官的生物量分配比例发生变化[23,44-45]。生物量分配有多种理论解释,其中最优分配理论表明,当光资源受限时,植物将增大茎和叶的生物量分配,使根冠比减小;而当营养受限时,植物将增大根的分配,使根冠比增加[23,38]。由于本研究3 种密度下生境土壤理化特征相近,因此可以排除土壤异质性带来的影响。在自然生境中,植物生物量分配主要受生境中光照强度、土壤水分和土壤养分等因素的影响,而且植物会将生物量较多地分配到可获得生存资源的器官中。
光是生境中最主要的环境因子,种群密度变化带来的光照资源差异及个体的邻近效应可能是北美车前个体生物量及各器官生物量分配差异的主要因素[46]。面对光改变时,植物体常通过改变生物量分配和形态,即通过形态可塑性来适应光照强度的改变[38]。叶片是植物进化过程中对环境变化较敏感且可塑性较大的器官,叶片也可通过多种途径改变外表特征以适应异质光环境的变化[23]。有研究表明,随密度增大,植株会增加地下部分生物量分配而减少地上部分生物量分配,表明植物对水、矿质营养和物理空间等地下资源的竞争能力随密度增加而增加,而对地上光资源的竞争能力随密度的增加而降低[25]。也有研究认为,在器官水平上,随密度增加植物根和叶生物量分配增加,茎和繁殖生物量分配减小,说明在高密度条件下,根生物量分配符合最优化理论中矿质营养受限的情况,而叶生物量分配的减少则可能是营养与繁殖权衡的结果[25,47]。而在本研究中,北美车前繁殖分配比例随密度增加而显著增加,至少证明在本研究所测定密度条件下,入侵植物北美车前仍以高繁殖输出来完成生活史,实现入侵和扩散。
3.2 密度对北美车前器官生物量异速生长关系的影响
异速生长分析表明,密度升高北美车前会提升叶片或减小繁殖生物量分配速率,但实际上,密度越高北美车前繁殖生物量比例越大而叶生物量比例越低。可见,这种分配速率的变化并未造成叶片实际分配比例高于繁殖分配比例,而仅仅是调整了生物量分配的速率,其繁殖生物量仍为增加趋势,但速率稍有降低。可见,高密度带来的种内竞争压力迫使北美车前不断提高繁殖分配比例以保证后代繁衍,而此时繁殖器官的构建也必然会提高对光合产物的需求,进而需要更高的光合器官投资速率。此外,生境条件对植物叶-繁殖器官异速生长关系存在一定程度的影响,但不同物种间并无一致规律[18,20,48]。对于不同生长期白刺(Nitraria tangutorum)而言,随繁殖生长持续推进,植物反而增加了对叶片的相对生物量分配比例[49]。不同固沙年限条件下,砂蓝刺头(Echinops gmelini)繁殖分配速率均高于地上生物量(本研究中为等速关系)[50]。镰叶锦鸡儿(Caragana aurantiaca)荚果数及荚果重与分株高及分枝数之间的异速生长指数均表现出生境差异性,即阶地生境高于河漫滩,表明土壤水分条件相对较差的阶地生境镰叶锦鸡儿具有更快的繁殖分配速率[51],该结果与本研究不同密度下同速分配的结果不同,本研究中北美车前在不同密度条件下,各器官之间更多体现为异速生长关系。虎尾草(Chloris virgata)生物量分配对不同降水条件的响应较为保守,其繁殖与非繁殖器官之间异速生长指数在不同降水条件下无显著差异[52],这与本研究结果相似。
研究发现,沙生植物猪毛菜(Salsola collina)的叶与地上、叶与根、叶与茎、根与地上和根与茎生物量间异速指数在不同种群密度间差异均不显著,表明叶与地上、叶与根、叶与茎、根与地上以及根与茎生物量间的变化是受个体大小制约的,不属于真正响应种群密度所发生的变化,属于表观可塑性;而种群密度对猪毛菜繁殖生物量和地上部分生物量间的异速指数和异速常数均产生了显著影响,表明种群密度的变化确实对猪毛菜繁殖器官生长产生了影响[42]。而无论猪毛菜[42]、沙蓬[23]或是大果虫实[25],种群密度均显著改变了繁殖器官与其他器官生物量间的异速指数和异速常数,这表明种群密度改变了植物的繁殖分配策略。本研究发现,无论何种密度条件下北美车前地上与地下生物量均呈异速生长关系,即地下根系分配速率高于地上部分;尽管如此,随密度增加北美车前地下生物量分配比例却呈显著降低趋势,也呈现出生物量分配速率和实际分配比例之间的不一致性。上述不一致性与其叶和根实际生物量比例远小于繁殖和地上生物量有关(图2),即尽管生物量较小的器官其分配速率高于生物量较大的器官,但因其基数小,导致实际分配的量和比例仍越来越小。同样,不同密度下北美车前繁殖器官与地上、繁殖器官与地下及繁殖器官与叶之间虽然各具有共同异速生长指数,但不同密度下的异速生长常数却有显著差异(表3),证实种群密度显著改变了北美车前的繁殖策略。尽管如此,不同器官之间资源分配的权衡,尤其是繁殖器官与叶之间的权衡,可能也是北美车前保持强大入侵性的重要基础。
等速生长假说是解释植物生物量分配的重要理论,尤其是地上与地下生物量等速生长关系,且已得到大量证实[53-54]。但是,地上与地下生物量间并非始终呈现等速关系,植物会因物种、功能群、环境梯度(如土壤养分、水分等)、人类影响(如生态系统管理与资源开发利用)等因素的不同而出现资源分配的自我调节,进而表现为异速生长关系,符合环境限制分配假说[12,55]。本研究中,等速检验和漂移类型分析表明,两种C 型异速生长关系表明器官之间具有完全相同的异速生长指数和相同的截距,发生共轴漂移是不同密度下个体大小差异引起的。3 种D 型异速生长关系的重要特征是有共同斜率、发生了共轴漂移但截距差异显著,这表明在不同密度和环境影响下北美车前相关器官之间的资源分配模式和协调机制,尤其体现在光合器官与繁殖器官之间。尽管如此,除地下与叶生物量(A 型)外,其他器官之间的异速生长指数相同,体现了恒定的资源分配速率和增长速率,这是同一物种不同种群或不同环境梯度下保持种内性状稳定性和内在生长发育规律的重要保证[55-56]。
3.3 本研究的价值与不足
本研究从个体水平探究了3 种自然密度梯度下入侵植物北美车前各器官生物量的分配特征及生物量间异速生长关系,揭示了不同种群密度对北美车前各器官的资源分配特征的影响,为其入侵机制的深入解析及亚热带地区的生态系统管理提供了一定的科学依据。但本研究也存在一定的不足之处:1) 未结合形态特征开展可塑性研究;2) 选取的密度梯度相对较少;3) 缺乏明确的环境梯度(如土壤环境),尚不能完全区分环境影响和密度制约之间的关系。因此,未来在这些方面还需要加强研究。
4 结论
3 种密度梯度下北美车前各器官生物量及生物量分配比例具有显著差异,其中低密度种群各器官生物量均最大而高密度种群最小,随密度升高繁殖分配比例逐渐增大而叶和根分配比例逐渐降低;表明密度增加北美车前种内竞争激烈,在有限的资源下各器官之间的资源分配出现权衡。各器官之间多为异速生长关系,但除了地下-叶生物量外,其他器官之间具有共同的异速生长指数,体现了不同竞争强度下恒定的资源分配速率。总之,入侵植物北美车前会调整自身资源分配比例和速率从而更好地适应不同竞争压力和环境条件。
