黄淮海地区夏大豆(南片)46份大豆品种(系)农艺性状综合分析
2021-11-17常世豪舒文涛杨青春李金花张东辉张保亮张来成
李 琼,常世豪,舒文涛,杨青春,李金花,张东辉,张保亮,张来成,耿 臻
(周口市农业科学院,河南周口 466000)
0 引 言
【研究意义】大豆起源于中国,大豆种质资源丰富[1]。我国黄淮海地区是中国大豆的主产区之一[2]。大豆产量相关性状都是数量性状,遗传与环境因素共同作用使得其农艺性状间存在信息的相互关联、重叠,增加育种难度[3]。综合分析2020年国家黄淮海夏大豆(南片)中间试验参试品种(系)的农艺性状,有利于了解该地区近期大豆新种质的遗传信息,对下一步制定育种目标、杂交组合选配有重要意义。【前人研究进展】大豆种质资源农艺性状综合评价是资源合理利用的前提[4-5]。大豆农艺性状各个指标存在一定的相关性。主成分分析方法能够有效的将多个相互关联性状通过降维得出几个相互独立的主成分,用于分析各个农艺性状指标在表型性状中发挥的作用[6]。汪宝卿等[7]将87份大豆常规栽培品种的12个农艺性状分为4个主成分。林文磊等[8]将39份春大豆种质的7个农艺性状分为2个主成分。常世豪等[9]将60份夏大豆种质的12个农艺性状分为4个主成分。聚类分析更清晰地了解资源的群体组成,研究不同品种(系)之间亲缘关系,易于目标种质的筛选[10]。采用主成分分析(principal component analysis,PCA)和聚类分析(cluster analysis)相结合的综合评价分析法已经广泛应用于玉米[11]、甘蓝[12]、茶树[13]、大蒜[14]、杨梅[15]等。【本研究切入点】大豆品种种植区域狭窄。目前关于国家黄淮海(南片)夏大豆中间试验参试品种的遗传多样性及优良亲本的筛选研究报道较少。研究黄淮海地区夏大豆(南片)的遗传背景、遗传多样性和综合性农艺性状。【拟解决的关键问题】选择2020年国家黄淮海夏大豆(南片)中间试验参试的46个新大豆种质资源,综合分析与评价大豆农艺性状遗传多样性,并进行主成分分析与聚类分析,从多角度分析和评价黄淮海夏大豆(南片)种质资源多样性,为大豆品种选育提供理论基础。
1 材料与方法
1.1 材 料
供试材料为2020年国家黄淮海夏大豆(南片)中间试验参试的46个大豆品种(系)。2020年将各品种(系)种植在周口市农业科学院许湾乡育种基地同一地点种植,均在材料完全成熟后收获。表1

表1 供试大豆品种(系)Table 1 Details of tested soybean varieties(lines)
1.2 方 法
试验地点位于周口市农业科学院许湾试验基地,供试材料为2020年国家黄淮海夏大豆(南片)中间试验参试材料。2020年6月20日一播全苗,按随机区组排列,3次重复,各重复每个小区7行,行长6 m,行距0.4 m,株距0.13 m,每个小区面积16.8 m2。各小区进行统一田间管理。每个小区收获期连续选择10株按照《大豆种质资源描述规范和数据标准》[16]室内考种。试验地年度内雨水频繁,未施肥、灌溉,整个生育期治虫2次。
1.3 数据处理
利用Microsoft Excel 2010软件进行遗传多样性分析(Shannon-Wiener’s多样性指数(Shannon-wiener diversity index,H’)[17-18]。利用SPSS Statistics 25.0软件对数据进行相关性分析、主成分分析,分别对农艺性状标准化数据和综合主成分值进行聚类分析(欧氏距离-瓦尔德(Ward)法)并比较。
变异系数的大小表明品种变异程度的大小,反映出各性状的变异丰富程度,遗传变异系数越大,变异越丰富[19]。利用Microsoft Excel 2010对国家黄淮海夏大豆(南片)中间试验参试的农艺性状遗传多样性信息指数进行计算,得出平均值、变幅、标准差、变异系数和遗传多样性等参数。
3个主成分对表型变异解释的贡献率(表)采用综合得分计算公式[20]。
2 结果与分析
2.1 大豆主要农艺性状的遗传多样性
研究表明,不同材料的农艺性状出现不同程度差异。有效分枝数(74.61%)和底荚高度(26.65%)的变异系数最大,主茎节数(12.41%)变异系数最小,株高、单株有效荚数、单株粒数、单株粒重、百粒重的变异系数在17.27%~23.95%。单株粒数(2.064)、单株有效荚数(1.989)和底荚高度(1.978)的遗传多样性指数最高,平均产量(1.697)的遗传多样性最低。各农艺性状遗传多样性指数介于1.697~2.064。9个农艺性状平均变异系数较大(24.67%),平均遗传多样性H’指数(1.918)较高。46份大豆品种资源的多样水平较高,黄淮海夏大豆(南片)种质资源具有丰富的遗传多样性。表2
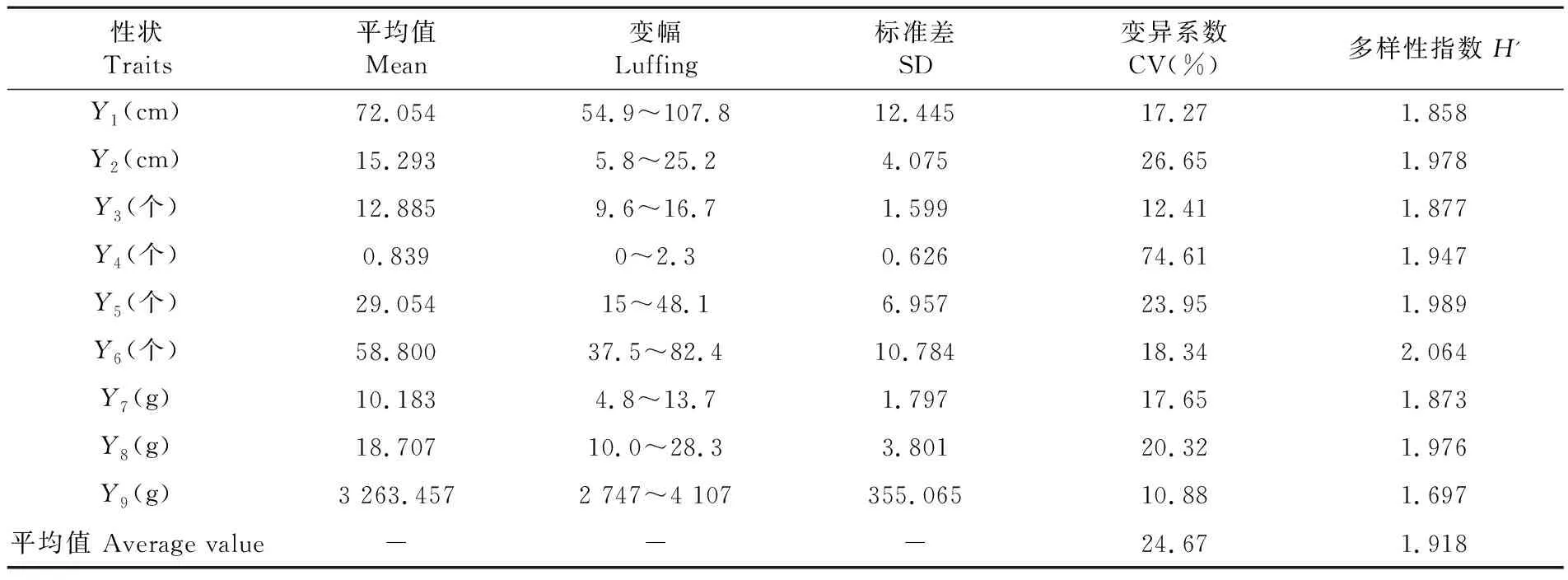
表2 大豆主要农艺性状的遗传多样性Table 2 Genetic diversity analysis of main agronomic traits
2.2 大豆主要农艺性状的相关性
研究表明,产量与单株粒数、单株粒重呈极显著正相关(P<0.01),与百粒重呈显著正相关(P<0.05);单株粒重与单株有效荚数、单株粒数均呈极显著正相关(P<0.01);百粒重与单株粒重呈极显著正相关(P<0.01),与有效分枝数呈显著负相关(P<0.05);分枝数与底荚高度呈极显著负相关(P<0.01)。表3

表3 主要农艺性状相关性Table 3 Correlation analysis of main agronomic traits
2.3 大豆主要农艺性状的主成分综合评价
研究表明,前3个主成分因子(F1、F2、F3)的特征值为2.645、1.721、1.620均大于1,并且其累计贡献率为66.5%。前3个主成分综合因子在所有农艺性状变异中起较大作用,代表大部分的表观遗传变异信息。表4,表5

表4 主要农艺性状主成分因子的方差贡献率Table 4 Variance contribution rate of principal component factors of main agronomic traits

表5 主要农艺性状主成分得分系数矩阵Table 5 Principal component score coefficient matrix of main of agronomic traits
第一个主成分F1的特征值为2.645,贡献率为29.384%。表达式:F1=0.190X1+0.132X2+0.188X3-0.018X4+0.246X5+0.303X6+0.266X7+0.092X8+0.240X9。第一个主成分因子主要是由单株有效荚数(X5)、单株粒数(X6)和单株粒重(X7)组成,均与产量有关,故称为“产量因子”。在一定范围内,第一主成分因子中单株有效荚数、单株粒数和单株粒重的变化与有效分枝数成反比,但有效分枝数权重系数仅为-0.018。
第二个主成分F2的特征值为1.721,贡献率为19.119%。表达式:F2=0.263X1+0.238X2+0.266X3+0.097X4+0.031X5+0.127X6-0.331X7-0.461X8-0.189X9。第二个主成分因子主要是由株高(X1)、底荚高度(X2)和主茎节数(X3)组成,均与株型有关,故称为“主茎因子”。在一定的范围内,第二主成分因子中主茎节数、株高和底荚高度的变化与百粒重成反比。
第三个主成分F3的特征值为1.620,贡献率为17.998%。表达式:F3=-0.177X1-0.364X2-0.162X3+0.515X4+0.215X5+0.237X6+0.044X7-0.238X8+0.028X9。第三个主成分因子主要是由有效分枝数(X4)组成,称为“分枝因子”。在一定范围内,第三主成分因子中有效分枝数的变化与底荚高度、百粒重成反比。
Z=(0.293 8F1+0.191 2F2+0.180 0F3)/0.665,Z为样本综合得分。得分较高的前5个品种(系)为郑1440、周豆34号、濮豆5110、恒豆6号、南农47。表6
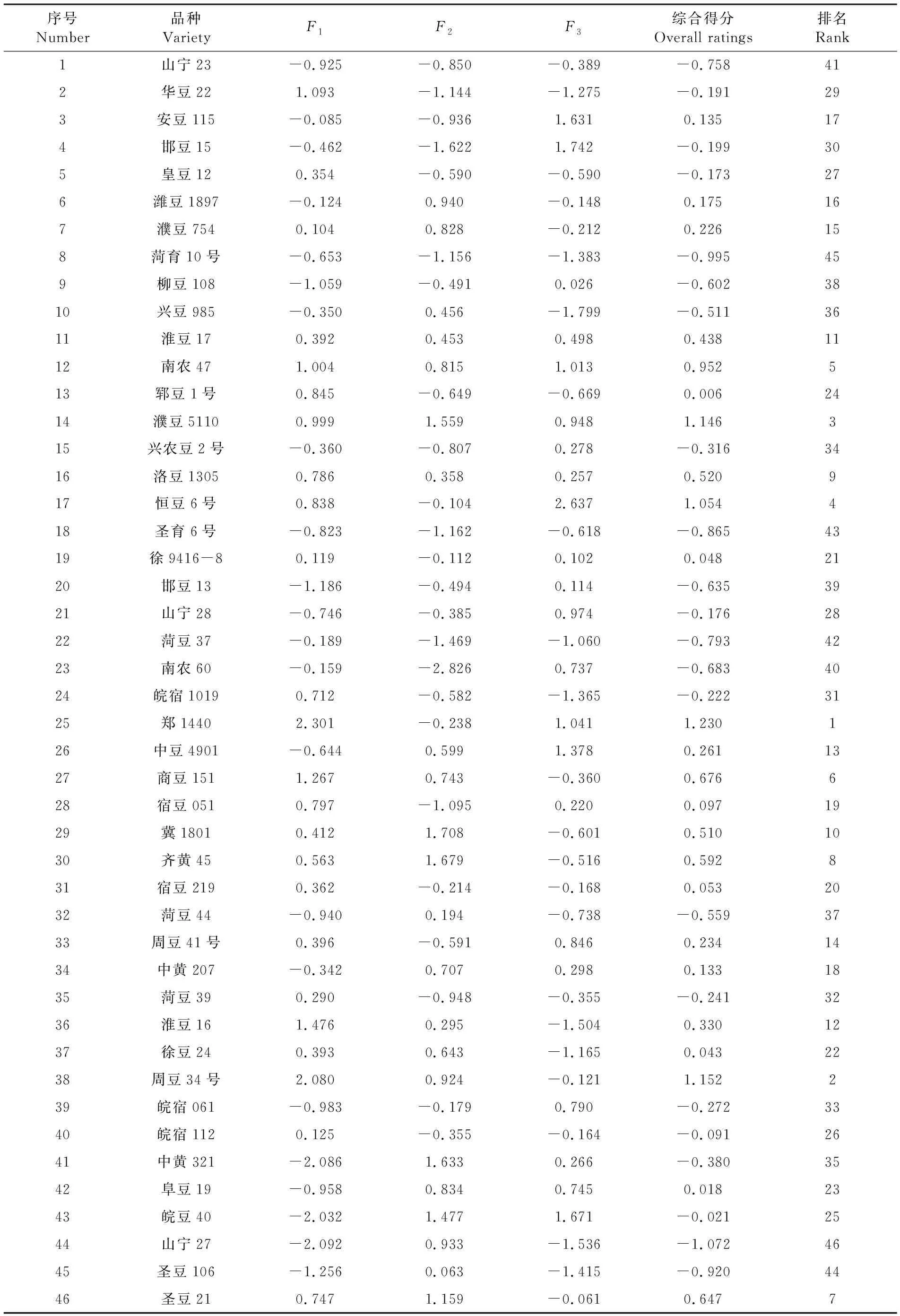
表6 参试大豆品种的主成分因子综合得分Table 6 Principal component factor comprehensive scores of tested varieties
2.4 基于主要农艺性状和综合得分聚类
研究表明,聚类可分为3个类群。第Ⅰ、Ⅱ、Ⅲ个类群分别包含25、13、8个品种(系)。其中,类群Ⅰ分2个亚类,第1个亚类包含15个品种(系),第2个亚类包含10个品种。3个类群中,类群Ⅰ属于株型较高大、分枝性较好、丰产性好、百粒重较大的品种。此类群品种(系)可作为选育多目标性状的综合性状较好的优良亲本。类群Ⅱ中百粒重最大,“产量因子”、“主茎因子”、“分枝因子”表现均差,且丰产性较差。类群Ⅲ“产量因子”中等,百粒重最小,株型中等偏小、分枝数最多,丰产性较好。
第Ⅰ、Ⅱ、Ⅲ个类群分别包含24、11、11个品种(系)。类群Ⅰ分2个亚群、类群Ⅲ分2个亚群。
相同点:2种聚类分析结果均把商豆151、圣豆21、齐黄45、洛豆1305、冀1801、淮豆17、濮豆5110、周豆34号、郑1440、南农47归到同一大类中,这10个大豆种质遗传距离比较近。2种聚类分析结果均把兴豆985、菏豆44、柳豆108、邯豆13、菏育10号、山宁27、山宁23、菏豆37、圣育6号、圣豆106归到同一大类中,这10个大豆种质遗传距离比较近。把中黄207、宿豆051、淮豆1897、徐9416-8、徐豆24、宿豆219、郓豆1号、濮豆754、周豆41号、淮豆16、皖宿1019、菏豆39、皇豆12、华豆22、皖宿112归到同一大类中,这15个大豆种质遗传距离比较近。把安豆115、皖豆40、中豆4901、皖宿061、中黄321、南宁60、恒豆6号、邯豆15归到同一大类中,这8个大豆种质遗传距离比较近。不同点:基于农艺性状的聚类分析法将山宁28、兴农豆2号、阜豆19归为类群Ⅱ,基于主成分综合评价的聚类分析法将这3个品种单独聚类到类群Ⅰ。图1
3 讨 论
3.1 黄淮海夏大豆(南片)种质资源丰富
黄淮海夏大豆(南片)种质资源具有丰富的遗传多样性。黄淮海夏大豆(南片)主茎节数一致性较强。株高、单株有效荚数、单株粒数、单株粒重、百粒重仍有相对改进的余地,可通过良种选配和栽培技术等方法使这些性状得到一定程度上的改善。有效分枝数、底荚高度有很大的遗传改良空间,这与林文磊等[9]对39份春大豆种质的7个主要农艺性状进行遗传变异分析得出底荚高度变异系数最高。常世豪等[10]对黄淮海60份夏大豆参试材料的12个主要农艺性状进行遗传变异分析。余飞等[21]对黄淮海11份夏大豆参试材料的12个主要农艺性状进行变异分析,均得出有效分枝数变异系数最大的研究结论相符。经过相关性分析得出,与大豆商品性息息相关的百粒重与有效分枝数呈显著负相关性,分枝数与底荚高度呈极显著负相关性。选育高产、大粒的品种要从有效分枝数和底荚高度两方面统筹兼顾。
3.2 黄淮海夏大豆(南片)种质资源区分和筛选
从主成分分析来看,其遗传信息可分为3个主成分因子,累计贡献率为66.5%,与以前研究分析所得值相比稍微较低[7,9]。可能是由于测定样本量大小不同造成的。要实现提高产量、增大百粒重双重目标,就要把握产量因子、主茎因子、分枝因子3个主成分因子作用的同时,控制主茎因子和分枝因子对百粒重的影响。大豆优良亲本的株型要适中,且避免了倒伏在生产中对产量的影响。这与以往的结果基本一致[22-24]。鉴于底荚高度和分枝有极显著负相关的关系,且变异系数较大,可适当提高底荚高度避免分枝过多,也便于生产上机械收割。要达到提高产量、增加百粒重的育种目标,要把握3个主成分因子相互关联、相互制约的密切关系,从3个主成分中的各要素进行综合考量[24]。
赵朝森等[25]对431份江西省大豆种质资源进行多样性分析,筛选出21份优异种质可用于我国高蛋白大豆新品种选育及基因挖掘。赵斌等[26]对111份多棱大麦种质主要农艺性状进行多样性分析,将供试材料分为3个大类,验证了基于表型性状对大麦种质遗传多样性评价的简单可行性,有助于更快培育出符合育种目标的大麦新品种。研究基于农艺性状的聚类分析可知:类群Ⅰ和类群Ⅲ可作为选育产量高、多目标性状综合性优良亲本和产量高、分枝性好的多分枝性优良亲本,且郑1440、周豆34号、濮豆5110、恒豆6号、南农47这综合得分较高的5个品种中郑1440、周豆34号、濮豆5110、南农47处在类群Ⅰ,恒豆6号处在类群Ⅲ。基于主成分分析与基于农艺性状的聚类结果比较得出两者存在一定的差异。
育种过程中要选育高产、大粒商品性好的大豆品种,其优良亲本的选择把握“产量因子”、“主茎因子”、“分枝因子”三者之间相互联系、相互制约的关键。利用2种聚类分析方法能有效地实现对大豆种质间的区分。获得更为精准的大豆种质信息仍需进行深层次的分子标记遗传多样性分析。
4 结 论
黄淮海夏大豆(南片)资源的农艺性状出现不同程度的差异。有效分枝数(74.61%)和底荚高度(26.65%)的变异系数最大,单株粒数(2.064)、单株有效荚数(1.989)和底荚高度(1.978)的遗传多样性指数最高。9个农艺性状平均变异系数较大(24.67%),平均遗传多样性H’指数(1.918)较高。黄淮海夏大豆(南片)种质资源具有丰富的遗传多样性。产量与单株粒数、单株粒重呈极显著正相关,与百粒重呈显著正相关;单株粒重与单株有效荚数、单株粒数均呈极显著正相关;百粒重与单株粒重呈极显著正相关,与有效分枝数呈显著负相关;分枝数与底荚高度呈极显著负相关。主成分分析将9个性状转化为3个主成分,累计贡献率达66.5%;郑1440、周豆34号、濮豆5110、恒豆6号、南农47五个品种(系)综合得分最高,可作为优良亲本参考改良当地大豆品种。聚类分析将供试材料分成3个类群,其中类群Ⅰ株型较高大、分枝性较好、百粒重较大、丰产性好,可作为选育多目标性状综合性状较好的优良亲本。类群Ⅱ百粒重最大,株型矮小,分枝数少,丰产性差,可作为选育大粒品种的优良亲本。类群Ⅲ分枝数最多,百粒重最小,株型中等偏小、丰产性一般,可作为选育多分枝品种的优良亲本。
