饲粮能量水平对白萨福克×湖羊F1羔羊生产性能和脂肪代谢相关基因表达的影响
2021-11-16张小强赵雷云黄晓瑜田林涛张恩平
张小强,赵雷云,黄晓瑜,杨 晨,田林涛,李 博,张恩平
(西北农林科技大学 动物科技学院,陕西 杨凌 712100)
羊肉具有高蛋白、低脂肪和低胆固醇等优点,随着社会经济发展和人民追求健康饮食理念的加强,优质羔羊肉深受市场青睐。2~4月龄的羔羊具有生长发育快、饲料转化率高等特点,因此断奶羔羊快速肥育成为生产优质羔羊肉的主要方式。饲粮能量是影响反刍动物的第一限制性因素[1],早期营养摄入不足会严重阻碍其生长发育。大量研究发现,饲粮能量水平显著影响羔羊的生产性能。吕小康等[2]发现,增加饲粮能量含量能显著提高2~4月龄湖羊羔羊的生长性能。Liu等[3]也发现,2月龄滩羊羔羊的体重、平均日增重和胴体重随着饲粮能量水平的升高而显著改善。血液不仅是动物体液最重要的组成部分,也是连接机体与外界的重要枢纽。Shakeri等[4]认为血清生化指标的变化与动物的能量代谢密切相关。Song等[5]发现,饲粮营养浓度可影响绵羊的血清生化指标。羔羊营养不足时,其血清生化指标也会随之变化[6-7]。经济杂交能够提高肉羊生产性能。有研究表明,萨湖F1代具有良好的育肥、屠宰性能以及上佳的肉质性状[8],但由于生产中肉绵羊杂交组合较多,目前对特定杂交羔羊育肥阶段营养需要的系统研究较少。本文通过对不同饲粮能量水平下萨湖F1代羔羊的生产性能、血液生化指标以及脂肪代谢基因表达的测定,探究饲粮能量水平对萨湖F1代羔羊早期育肥的影响,为杂交羔羊早期育肥的营养需要提供参考依据。
1 材料与方法
1.1 试验时间和地点
本试验于2020年7月24日-2020年10月10日在甘肃省庆阳市环县庆环肉羊制种有限公司进行。试验周期为78 d,其中预试期8 d,正试期70 d。
1.2 试验动物与试验设计
选取2月龄左右、平均体重为20.58±1.47 kg的健康白萨福克羊(♂)×湖羊(♀)F1代杂种羔羊36只,随机分为3组,每组4个重复(公母各2个重复),每个重复3只羊,分别饲喂低能组(LE)、标准组(CON)和高能组(HE)试验饲粮。
1.3 试验饲粮
根据肉羊饲养标准(NY/T816-2004)[9]中体重为20.0 kg,目标日增重为200 g/d的育肥羔羊营养设计标准组(CON)饲粮,消化能为10.38 MJ/kg,粗蛋白质为14.36%;低能组(LE)饲粮设计能量水平为标准组的85%,消化能配方计算值为8.67 MJ/kg;高能组(HE)饲粮设计能量水平为标准组的115%,消化能配方计算值为12.31 MJ/kg,LE组和HE组饲粮蛋白水平与标准组一致。3组饲粮均制成颗粒饲料饲喂。饲粮配方组成及营养水平见表1。

表1 饲粮组成及营养水平(风干基础)Table 1 Diet composition and nutritional level (air-dry basis)
1.4 饲养管理
试验开始之前对圈舍、水槽、料槽以及走廊等进行全面清扫和消毒,并对试验羊按程序免疫(三联四防)和体内驱虫(伊维菌素)。试验期各处理组分别饲喂对应能量水平的颗粒饲粮,每日分别于08:30和17:00投料2次,同时观察试验羊的健康状况,试验羊自由采食,自由饮水。
1.5 样品采集和指标测定
1.5.1 生产性能的测定 以试验设置的重复为单位记录各组的饲喂量和剩料量,并测定试验羊的初始体重和终末体重,计算干物质采食量(DMI)、平均日增重(ADG)和料肉比(F/G)。饲养试验结束后,从每个重复中随机选取1只试验羊(共12只)进行屠宰(空腹24 h),记录宰前活重和胴体重,计算屠宰率。采集皮下脂、尾脂和背最长肌组织样品,并做好RNA提取样品前处理。
1.5.2 胴体重 屠宰后去除头、蹄、尾、内脏,保留肾脏,称量胴体重。
1.5.3 屠宰率
屠宰率(%)=胴体重/宰前活重×100%
1.5.4 血液生化指标的测定 试验结束的当天晨饲前,所有试验羊颈静脉采血,静置30 min,用低速离心机以3 500 r/min离心10 min,移取上层血清于2 mL离心管内,于-20 ℃冰箱内保存待测。采用迈瑞BS-420全自动生化仪(深圳迈瑞生物医疗电子股份有限公司)测定总胆固醇(TC)、甘油三酯(TG)、葡萄糖(GLU)、高密度脂蛋白(HDL)、低密度脂蛋白(LDL)、总蛋白(TP)、白蛋白(ALB)、球蛋白(GLB)、尿素(BUN)和生长激素(GH)含量。试剂盒由中生北控股有限公司提供。
1.5.5 相关基因表达检测
1.5.5.1 RNA的提取和cDNA的合成 将皮下脂、尾脂和背最长肌组织从装有保存液的冻存管内取出,按照RNA提取试剂盒的说明书提取组织的总RNA,运用核酸定量仪(K5800,上海DRAWELL科学仪器有限公司)测定总RNA浓度和纯度,用1%琼脂糖凝胶电泳检测RNA的完整性。按照反转录试剂盒说明进行操作将RNA反转录为cDNA,于-20 ℃冰箱中保存。
1.5.5.2 引物设计与合成 依据NCBI(https://www.ncbi.nlm.nih.gov/)公布的绵羊脂肪酸合成酶基因(FASN)、乙酰CoA羧化酶基因(ACC)、过氧化物酶激活受体基因(PPARγ)、脂肪酸结合蛋白4基因(FABP4)、激素敏感脂肪酶基因(HSL)和内参基因β-肌动蛋白(β-actin)的mRNA序列使用软件Primer 5.0 设计特异性引物,交由陕西中科羽瞳生物公司合成,引物序列见表2。

表2 引物序列Table 2 Primer sequences
1.5.5.3 荧光定量PCR 反应体系为20 μL:2× SYBR qPCR Master Mix 10 μL,cDNA 1 μL,上下游引物各0.4 μL,ddH2O 8.2 μL。PCR扩增条件:95 ℃预变性30 s,95 ℃变性10 s,60 ℃延伸退火30 s,40个循环。融解条件:95 ℃下15 s,60 ℃下60 s,95 ℃下15 s,循环1次,检测结果由LightCyler 96定量仪给出,以β-actin为内参基因,各目的基因反转录强度以基因绝对拷贝数/β-actin绝对拷贝数表示,通过熔解曲线判断是否有非特异性扩增产物或引物二聚体出现,采用2-ΔΔCT法进行相对定量。
1.6 统计分析
采用Excel 2010对试验数据进行初步整理,使用GraphPad Primer 8.0 软件做图;使用SPSS 25.0统计软件中的单因素方差分析(One-Way ANOVA)和Duncan氏法进行多重比较,数据用平均值±标准差表示,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果与分析
2.1 试验羔羊的生产性能
饲粮能量水平对羔羊生产性能的影响数据见表3。由表3可知,初始体重各处理组间无差异。试验期羔羊末重和平均日增重HE组和CON组显著高于LE组(P<0.05);日均干物质采食量CON组显著高于HE组(P<0.05);胴体重HE组生产成绩最好,显著高于LE组(P<0.05),与CON组无显著差异。屠宰率HE组较LE组和CON组升高了9.95%和7.09%。该结果表明,中国肉羊饲养标准(NY/T 816-2004)[9]适用于萨湖F1代,适当提高饲粮能量水平能够提高饲料转化效率和肉羊屠宰率。
2.2 试验羔羊的血液生化指标
由表4可知,饲粮能量水平对血清TC、TG、LDL、HDL和TP含量有显著影响, HE组的血清TC含量较CON和LE组升高了35.53%和37.56%(P<0.05);HE组的血清TG含量较CON组和LE组均升高了33.33%(P<0.05);HE组的LDL含量较CON组和LE组升高了35.53%和37.56% (P<0.05);HE组的HDL较CON组和LE组升高了27.78%和23.15%(P<0.05);HE组的TP较CON组和LE组升高了8.12%和8.35%(P<0.05);血清GLOB含量随着能量水平的上升有升高趋势(P=0.054)。
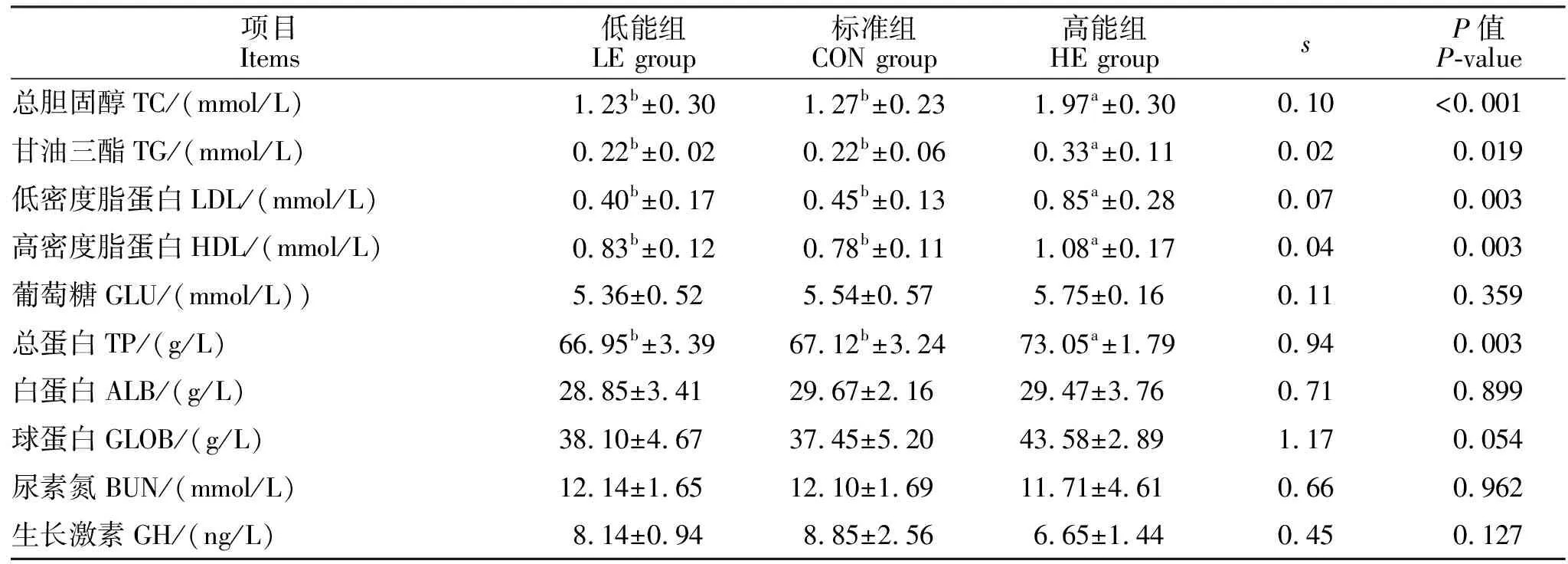
表4 饲粮能量水平对血清生化指标的影响Table 4 Effect of dietary energy level on serum biochemical indexes
2.3 试验羔羊脂肪代谢相关基因mRNA的表达
饲粮能量水平对羔羊脂肪代谢相关基因mRNA表达的影响见图1。由图1A可知,饲粮能量水平对皮下脂中基因ACC、FABP4和HSLmRNA的表达有显著影响。ACCmRNA的表达随能量水平的升高而升高,HE组显著高于LE组(P<0.05)。HSLmRNA的表达量在3组之间差异显著,HE组高于CON组高于LE组。CON组FABP4的表达水平最高,较LE和HE组分别提高了56.90%和58.62%(P<0.05);饲粮能量水平对皮下脂肪中FASN和PPARγmRNA的表达无影响(P>0.05)。
由图1B可知,饲粮营养水平对尾脂中基因FASN、ACC、FABP4和HSLmRNA的表达有显著影响。FASNmRNA的表达量HE组最高,显著高于CON组和LE组(P<0.05);尾脂中ACCmRNA的表达模式与皮下脂中的相同,随着能量水平的升高而逐渐升高,HE组显著高于LE组(P<0.05);FABP4 mRNA的表达量CON组最高,较LE组和HE组升高了31.97%和25.85%(P<0.05);HSLmRNA的表达量LE组最高,较CON和HE组升高了18.00%和16.00%(P<0.05)。
由图1C可知,饲粮能量水平对背最长肌中脂肪沉积相关基因mRNA的表达均有显著影响。FASNmRNA的表达量CON组最高,分别较LE组和HE组提高了40.83%和42.01%(P<0.05);ACCmRNA的表达量CON组最低,分别较LE和HE组下降了57.44%和30.55%(P<0.05);PPARγmRNA的表达量HE组最低,分别较LE和CON组降低了16.00%和19.00%(P<0.05);HSLmRNA的表达量LE组最高,分别较CON和HE组增加了83.00%、82.00%(P<0.05)。FABP4 mRNA的表达量HE组显著高于CON组(P<0.05),CON组显著高于LE组(P<0.05)。
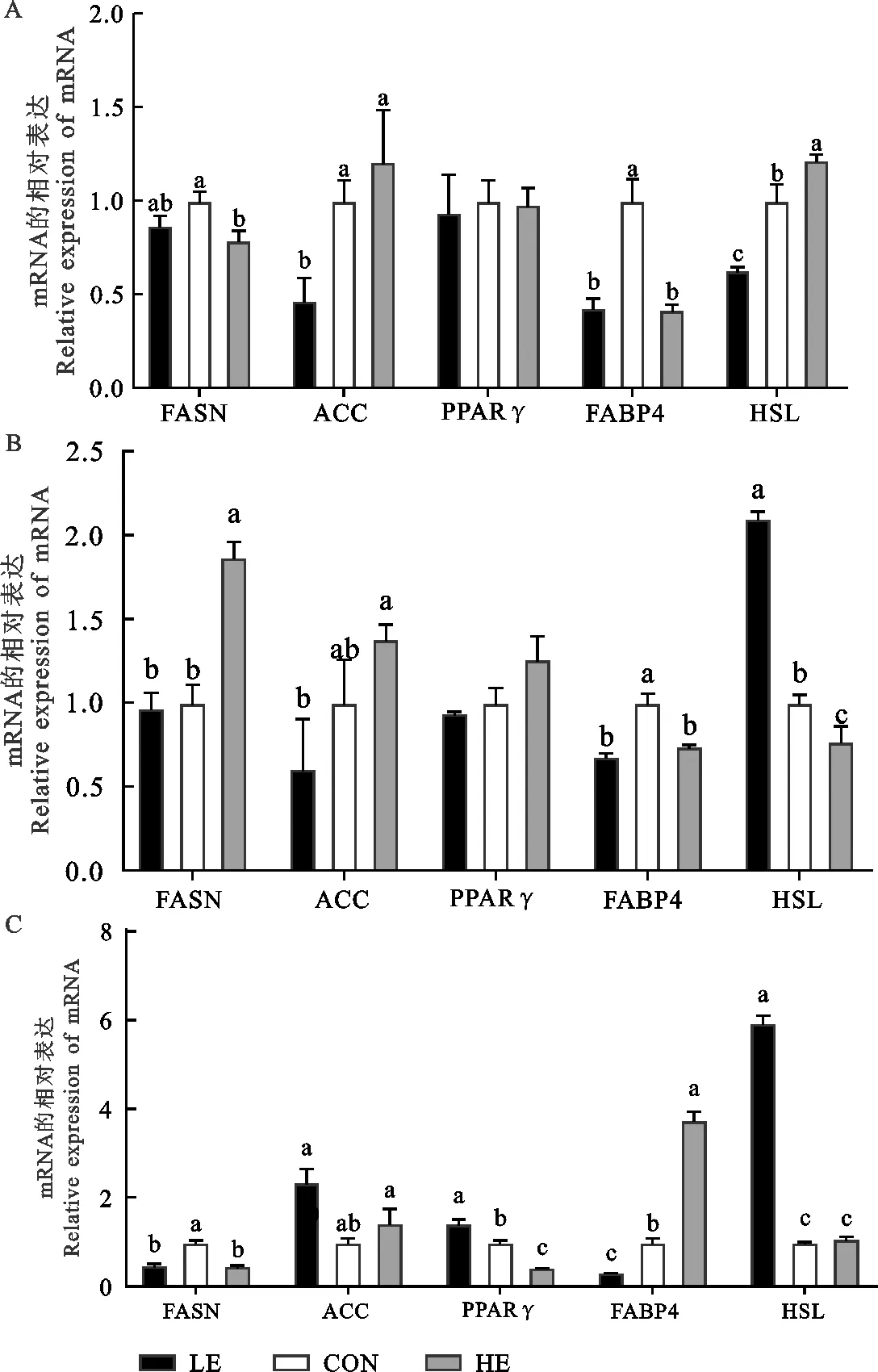
图1 萨湖F1代羔羊脂肪组织脂肪代谢相关基因mRNA的表达A.皮下脂肪;B.尾部脂肪;C.背最长肌脂肪同一基因标相同字母或无字母表示差异不显著(P>0.05),不同小写字母表示差异显著(P<0.05)Fig. 1 mRNA expression of genes related to fat metabolism inadipose tissue of White Suffolk×Hu F1 Lambs A. subcutaneous adipose; B. tail fat; C. fat in longissimus dorsiIn the same gene, the same or no letter superscripts indicateinsignificant difference(P>0.05), while different lowercasesuperscripts mean significant difference (P<0.05)
3 讨 论
3.1 饲粮能量水平对羔羊生产性能的影响
饲粮能量水平是影响反刍动物生长发育的第一限制性因素[1]。前人研究发现饲粮能量水平影响不同品种羔羊的体重、ADG和饲料转化率,但却存在一定差异。Yagoub等[10]研究发现,饲粮能量水平的升高(ME:8.5~11.5 MJ/kg)可极显著提高苏丹羊羔羊的末重、ADG和饲料转化率。Ebrahimi等[11]也发现,低能量饲粮可显著降低梅赫拉班公羔的ADG和体重。本试验结果与前人研究一致,羔羊末重和ADG随营养水平的升高而显著升高。研究发现,动物为能而食,可依据饲粮能量水平自主调整采食量,进而提高饲料报酬和体重[12]。Brand等[13]发现,羔羊采食量随着饲粮能量的提高而降低。在本试验中,高能组的干物质采食量最低,且显著低于标准组。可能因为高能饲粮更容易满足羔羊的营养需要,动物根据能量水平自主调整采食量,进而造成采食量的下降。料肉比(F/G)反映饲料利用率的高低。吕小康等[2]也发现,提高饲粮能量水平显著降低90~120日龄湖羊羔羊的料肉比。这与本试验的结果一致,料肉比随着能量的升高而极显著下降,表明高能量饲粮能显著促进羔羊的饲料转化率,提高经济效益。
研究发现,饲料营养水平与屠宰性能也有一定联系。Wang等[14]发现不同能量水平的饲粮(ME为9.17~10.82 MJ/kg)可显著提高20.0 kg肥育湖羊公羔的宰前活重和胴体重。Wang等[15]发现能量水平(ME为9.17~10.41 MJ/kg)的升高可有效增加4月龄湖羊公羔的宰前活重和胴体重。本试验结果与前人的一致,HE组(12.31 MJ/kg)的宰前活重和胴体重显著高于LE组(8.67 MJ/kg),屠宰率也得到显著提高。在本试验中,LE组饲粮精粗比为40:60,CON组和HE组的饲粮精粗比一致为50:50,卢盛勇等[16]研究发现,精粗比为50:50与40:60的日粮对贵州白山羊(15.96±2.94 kg)的生长性能和屠宰性能无显著影响,这与高林青等[17]在湖羊羔羊(15.62±0.01 kg)上的研究结果相同,这表明可将因饲粮的精粗比差异对羔羊育肥效果的影响排除在外。本试验结果表明,饲粮能量水平的提高可显著影响白萨湖羔羊的肥育期生产性能,在中国肉羊饲养标准(NY/T 816-2004)[9]的基础上增加15%可取得良好饲喂效果。
3.2 饲粮能量水平对羔羊血清生化指标的影响
能量贮存于饲粮的各种养分中,饲粮能量的消化、转化和利用总是伴随着物质代谢同步进行,血液是体内循环流通的代谢池,血液中相关生化指标可间接反映机体的物质代谢情况[18]。霍俊宏等[19]研究发现,精粗比为40:60与50:50的日粮对努比亚山羊(20.07±0.57 kg)的血清生化指标无显著影响。本试验中CON组和HE组的饲粮精粗比相同,这说明在本试验中饲粮精粗比的差异对白萨湖羔羊的育肥效果无影响。血清TC和TG指标均与能量摄入有关,反映脂类的代谢和吸收[20]。江赵宁等[21]研究表明,随着能量水平的升高,妊娠期母猪血清中的TG和TC含量呈显著上升趋势。本试验的研究结果与前人一致,血清TC和TG含量随着饲粮能量水平的升高呈升高趋势,血清TC和TG含量HE组显著高于CON组和LE组,说明高能量饲粮可有效促进动物的脂代谢过程。
HDL和LDL都是胆固醇的运载介质,其含量高低与体脂沉积密切相关。HDL可将血液中的TC和TG运输到肝内和脂肪组织中进行降解,进而减少TG的沉积。LDL因能量摄入其水平显著降低,可以有效防止脂代谢的紊乱[22]。刘爽[23]研究表明,架子牛血清中的HDL和LDL水平均随能量水平的增加而升高。孙国虎[24]发现绵羊血清LDL水平随能量(ME为7.85~11.67 MJ/kg)的升高而显著增加。在本试验中,血清HDL和LDL含量HE组极显著高于CON组和LE组,可能因为高能量水平的饲粮先造成机体TC和TG含量的升高,进而促进机体中HDL和LDL的合成与分泌,用于转运TC和TG到肝脏组织和脂肪组织中进行降解,最终促进机体脂肪沉积和内环境的稳态平衡。
血清葡萄糖水平反映机体能量代谢水平的高低,动物GLU浓度应在6.1 mmol/L之下,否则影响机体的糖代谢运转[25]。Chelikani等[26]发现血清中的GLU随着饮食能量的增加而增加。在本试验中,血清GLU含量随着能量水平的升高而呈升高趋势,且均在6.1 mmol/L以下,无显著差异。这表明在本试验中,较高能量水平可促进机体的糖代谢增强,但并未影响糖代谢的正常运转。
TP包括ALB和GLOB。TP含量的升高可促进动物组织的蛋白质沉积,ALB能直接反映机体营养状态的高低,GLOB可参与机体的免疫活动。钟港等[27]发现高能饲粮(16.42 MJ/kg)组湘南黄牛的血清TP水平高于低能组(14.93 MJ/kg),但低能组的ALB水平要优于高能组。田春丽[28]发现滩羊(22.0~25.0kg)的血清ALB和GLOB在不同饲粮能量水平下(7.45~11.32 MJ/kg)无显著影响。本试验结果为血清TP水平随着能量水平的升高而显著升高,HE组的血清TP水平显著高于CON组和LE组。这表明高能饲粮可提高机体的蛋白质代谢,可能是因为高能饲粮为瘤胃微生物提供丰富的营养物质,促进微生物的繁殖和生长,进而产生更多的菌体蛋白,最终促进机体的蛋白质代谢。而各组的血清ALB和GLOB含量无显著差异,这与田春丽的研究结果一致。本试验结果提示,适当提高饲粮能量水平可提高机体的蛋白质利用率。
血清BUN含量反映动物机体的氮代谢情况,与蛋白合成率和氨基酸平衡呈负相关[29]。吕小康等[2]指出,提高能量水平可显著降低4月龄湖羊羔羊的血清BUN含量,这与本试验结果相似。在本试验中,血清BUN水平与能量呈负相关,随着能量水平的升高而下降,当能量水平提高到12.31 MJ/kg 时,可促进机体的氮沉积和氨基酸利用效率。
GH的主要作用是促进脂肪分解与蛋白质合成,进而控制动物的生长和营养物质代谢。崔原年等[30]发现,高营养水平可降低反刍动物的GH浓度,而限饲时血浆GH浓度显著高于自由采食的。在本研究中,HE组血清GH水平分别较CON组和LE组下降了24.86%和18.30%,与前人研究一致。因此提高饲粮能量水平会促进羔羊的脂代谢和蛋白质代谢。
3.3 饲粮能量水平对脂肪组织中脂肪代谢相关基因表达的影响
体脂沉积与动物自身的营养代谢密切相关,不仅可以缓解因能量过度摄入而造成的器官损伤,在必要时还可以作为机体的主要能量来源。脂肪组织的生长发育主要通过细胞的增殖与分化,并由多种脂肪代谢基因共同调控,其中包括FASN、ACC、PPARγ、FABP4和HSL等基因。大量研究表明,饲粮能量水平与体脂沉积有一定相关性。高良霜[31]发现,高能饲粮(0.75 MJ/W0.75×d-1)显著促进阿勒泰羊皮下脂和尾脂中基因ACC和PPARγ的表达。刘培峰等[32]发现,随着能量水平(NE:9.84~10.67 MJ/kg)的增加,38.00 kg丹系三元猪的背膘厚度呈线性增加。
Liu等[3]发现,4月龄滩羊的皮下脂肪和尾脂中FASN随着能量水平的升高而显著升高,而HSL表达量则显著下降。Zhang等[33]在杂交牛皮下脂肪中发现,FASN、ACC、PPARγ随着能量水平的增加而显著升高,而HSL的表达量则显著下降。在本试验中,低饲粮营养水平对脂肪组织中ACC和FABP4mRNA表达影响相同,表达量均降低,与前人研究一致。但HE组基因FABP4的表达有降低趋势,可能是因为饲粮营养水平对其的影响已达到阈值,再提高能量水平,反而会降低其基因的表达;能量水平对皮下脂和尾脂中PPARγ表达的影响差异不显著,但随着营养水平的升高而呈升高趋势。在本试验中,饲粮能量对脂肪组织中基因PPARγ和FABP4表达的影响趋势相同,这与王小芳[34]提出的饲粮营养对FABP4基因的调控可能由PPARγ来实现的观点一致。饲粮能量对皮下脂和尾脂中FASN基因表达的影响不同,在尾脂中显著升高,而在皮下脂肪中无显著影响,可能是因为羔羊皮下脂和尾脂沉积存在空间上的差异。
而营养水平对皮下脂和尾脂中HSL的表达影响相反,在皮下脂中显著提高,但在尾脂中则显著下降。这与Zhang等[33]和Liu等[3]的研究结果不同,分析其原因可能有二:一是本试验采样时羔羊皮下脂可能已经沉积完全, FASN是脂肪从头合成的关键酶,当皮下脂的脂肪酸从头合成已经达到饱和时,过多的能量对皮下脂而言成为负担,因此脂解基因HSL的表达显著升高用于维持皮下脂中的正常代谢;二是因为高能量饲粮的摄入先引起机体血液中的TC和TG含量的显著升高,进而促使机体合成与分泌大量的LDL和HDL,用于转运TC和TG到肝脏和脂肪组织中进行降解,以维持自身的健康稳态和营养代谢平衡。皮下脂肪是最先沉积的[35],因此很大可能是将过多的TC和TG转运到皮下脂肪中进行降解,最终促进皮下脂肪中HSL基因的表达。本试验表明,提高饲粮能量水平可促进萨湖F1代羔羊育肥期的皮下脂肪和尾脂的沉积。
3.4 饲粮能量水平对背最长肌中脂肪代谢基因表达的影响
肌内脂肪的沉积可以改善羊肉的品质和风味特征,提高肌内脂的沉积也是高档羊肉生产过程中的关键技术难点和突破点。Zhang等[36]报道称,西门塔尔牛×黄种牛肌肉中的FASN和FABP4的表达随着能量的升高而显著升高。Kang等[37]也发现基因FASN表达随能量的升高而显著增加。这与本试验的结果一致,FABP4随着饲粮能量水平的升高而极显著升高,FASN随着能量的升高而先升高后下降,可能是能量水平对其的影响已达到最大,能量水平的再升高,其表达水平反而会降低。郝称莉[38]发现4月龄后的湖羊PPARγ与IMF含量呈负相关(r=-0.441)。Huang等[39]在雄性萨福克羊上也证明了PPARγ的表达对IMF含量有负面作用。这与前人的结果一致,可能是因为PPARγ主要表达在脂肪组织中,是脂肪胞质内特定性转录因子[38]。而肌内脂肪是最后沉积的,故PPARγ在肌肉组织中其表达水平较低,且饲粮营养水平对其的影响与肌内脂肪沉积呈负相关。
在本试验中,LE组的ACC表达水平最高,这与Zhang等[36]的研究结果不同。有研究发现,ACC有两种亚型(ACC1和ACC2),ACC1主要存在于肝脏和脂肪组织中,促进脂肪酸的合成,ACC2主要存在于骨骼肌和心肌中,促进脂解供能[40]。在本试验中,LE组ACC表达水平较高,可能是因为LE组的饲粮能量供给难以满足动物自身的生长需要,总体表现为脂解功能,进而维持机体的正常发育。Kim等[41]研究表明,随着饲粮能量的升高,HSL表达显著降低。Zhang等[36]研究发现,西门塔尔牛×黄种牛的背肌中HSL的表达随着能量水平的升高而显著下降。这与本试验的研究结果一致,脂解基因HSL的表达随着能量摄入的增加而显著降低,说明饲粮营养水平通过下调脂解基因HSL的表达促进背最长肌中的脂肪沉积。
4 结 论
在本研究中,综合生产性能、血清生化指标和脂肪沉积相关基因的表达,饲粮能量水平为12.31 MJ/kg时更适用于白萨福克×湖羊F1肥育期羔羊的生长发育和体脂沉积。
