过表达性别决定区Y框蛋白7基因对骨肉瘤细胞生长及免疫逃逸相关因子表达的影响
2021-11-15于云祥龚泰芳刘小涛柯文李彬彬廉凯徐进
于云祥 龚泰芳 刘小涛 柯文 李彬彬 廉凯 徐进△
骨肉瘤是常见的原发性恶性骨肿瘤,恶性程度高,预后差,5年生存率低[1]。近年分子靶向治疗成为肿瘤治疗研究热点。尽管骨肉瘤治疗方法已不断更新,但是治疗效果却不甚理想,寻找有效的治疗方法具有重要的意义。SOX7是含高迁移率组蛋白(HMG)结构域的SOX基因家族SOXF亚组成员之一,在癌症中低表达,对肿瘤发生发展具有重要作用[2-4]。有研究表明miR-664能通过靶向抑制SOX7增强骨肉瘤细胞的侵袭和迁移能力[5]。Wnt/β-catenin信号通路是比较经典的信号转导途径,与肿瘤发展密切相关,如下调RIPK4可通过抑制Wnt/β-catenin信号通路抑制骨肉瘤细胞增殖[6]。但SOX7与骨肉瘤细胞凋亡、免疫反应及其可能相关的Wnt/β-catenin信号通路的关系还未明确。鉴于此,本研究通过建立重组体pcDNA3.1-SOX7,旨在探讨过表达SOX7对骨肉瘤细胞生长、周期、凋亡、免疫抑制因子VEGF和TGF-β1表达及Wnt/β-catenin信号通路的影响。
1 材料与方法
1.1 试剂和仪器
RPMI-1640培养基、CCK-8试剂盒均购自美国Sigma;FBS购自美国Gibco;pcDNA3.1载体及Lipofectamine 2000试剂盒均购自Invitrogen 公司;SOX7、β-catenin、cyclinD1、survivin、VEGF和TGF-β1抗体及HRP标记的二抗均购自美国CST;细胞凋亡试剂盒、流式细胞仪均购自美国BD;酶标仪及凝胶成像系统均购自美国Bio-Rad。
1.2 方法
1.2.1细胞培养 人骨肉瘤MG-63细胞株购自中国科学院上海细胞库。细胞用RPMI-1640培养基(含100 U/mL青霉素、100 U/mL链霉素及10% FBS),在5%CO2、37 ℃恒温、饱和湿度条件下传代培养。细胞生长状态良好时用于实验。每2~3 d换液1次,依据细胞密度传代。
1.2.2细胞转染 转染前24 h调整MG-63细胞密度,以2 mL/孔(5×105/mL)接种于6孔板。分为重组pcDNA3.1-SOX7组、pcDNA3.1组和空白组。无双抗的完全培养基培养MG-63细胞24 h,使转染时细胞达90%~95%的细胞密度。无血清培养基冲洗6孔板内细胞两遍,加2 mL无血清培养基。无血清培养基分别稀释pcDNA3.1-SOX7、pcDNA3.1及Lipofectamine 2000,轻摇混匀。将稀释好的pcDNA3.1-SOX7和pcDNA3.1分别与Lipofectamine 2000混合,室温孵育20 min,加入6孔板相应的孔内,在5%CO2、37 ℃恒温、饱和湿度培养箱中孵育6 h,更换为含血清的完全培养基。48 h后通过Western Blot检测转染效果。
1.2.3Western Blot检测 收集转染pcDNA3.1-SOX7和pcDNA3.1 48 h的MG-63细胞,加入适量RIPA裂解液,在冰上反应30 min,离心,收集上清。BCA试剂盒测定上清蛋白浓度。蛋白与上样缓冲液混匀,煮沸变性5 min。取40 μg变性蛋白上样,经SDS-PAGE、半干法转NC膜及5%脱脂奶粉封闭膜后,洗膜,加稀释后的SOX7、β-catenin、CyclinD1、Survivin、VEGF和TGF-β1抗体(稀释比例1∶1 000),4 ℃孵育过夜,洗膜,加1∶3 000稀释的HRP标记的羊抗鼠二抗,37 ℃摇床震荡1 h,洗膜,ECL显色。Quantity One软件进行灰度值分析。蛋白相对表达量=目的蛋白灰度值/内参GAPDH灰度值。实验重复3次。
1.2.4CCK-8法检测细胞活力 取生长至对数期的MG-63细胞,以5×103/孔接种于96孔板,参照前述方法转染pcDNA3.1-SOX7和pcDNA3.1,并设置空白组,在培养的24,48和72 h收集上述3组细胞,在每孔中加入10 μL CCK-8试剂,继续孵育4 h,450 nm吸光度酶标仪测定各孔的吸光度值(A)。以A值绘制细胞生长曲线。实验重复3次。
1.2.5流式细胞仪检测细胞周期和凋亡率 按照前述方法转染pcDNA3.1-SOX7后48 h,收集细胞,将细胞制备成单细胞悬液,计数1×106个细胞,常规固定处理,PI染色,筛网过滤细胞悬液后,使用流式细胞仪检测各组细胞周期分布。同样方法收集细胞,按照Annexin V-FITC/PI双染细胞凋亡试剂盒说明书染色,避光室温放置15 min,流式细胞仪检测各组细胞的凋亡率。实验重复3次。
1.3 统计学方法
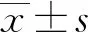
2 结果
2.1 pcDNA3.1-SOX7转染MG-63细胞效率
pcDNA3.1-SOX7转染MG-63细胞48 h,通过Western Blot检测转染效果,结果如图1和表1所示,转染重组体pcDNA3.1-SOX7后,MG-63细胞SOX7表达明显升高,差异有统计学意义(P<0.05),而转染pcDNA3.1空载体组SOX7表达与空白组差异无统计学意义(P>0.05)。

表1 pcDNA3.1-SOX7转染MG-63细胞后SOX7的蛋白相对表达量

图1 pcDNA3.1-SOX7转染MG-63细胞后SOX7蛋白表达
2.2 过表达SOX7抑制MG-63细胞活力
如表2所示,pcDNA3.1-SOX7转染MG-63细胞24,48和72 h,细胞活力均明显降低,差异有统计学意义(P<0.05),而pcDNA3.1组在3个时间点的细胞活力与空白组差异无统计学意义(P>0.05)。
表2 转染pcDNA3.1-SOX7的MG-63细胞活力
2.3 过表达SOX7阻滞MG-63细胞周期
如表3所示,与空白组比较,pcDNA3.1-SOX7组G0/G1期细胞百分比升高,G2/M期和S期细胞百分比降低,差异均有统计学意义(P<0.05),不同组间、不同时间点间、组间×时间交互作用下的过表达SOX7阻滞MG-63细胞周期差异均有统计学意义(P<0.05),说明细胞发生G0/G1期阻滞。

表3 过表达SOX7对MG-63细胞周期的影响
2.4 过表达SOX7诱导MG-63细胞凋亡
如图2和表4所示,pcDNA3.1-SOX7转染MG-63细胞48 h,细胞凋亡率明显高于空白组(P<0.05)。

图2 过表达SOX7对MG-63细胞凋亡的影响
表4 过表达SOX7后的各组MG-63细胞凋亡率
2.5 过表达SOX7抑制MG-63细胞VEGF和TGF-β1表达
通过Western Blot检测转染pcDNA3.1-SOX7的MG-63细胞免疫抑制相关因子VEGF和TGF-β1表达,结果如图3和表5所示,pcDNA3.1-SOX7组VEGF和TGF-β1蛋白表达均明显低于空白组(P<0.05)。
表5 过表达SOX7的MG-63细胞VEGF和TGF-β1蛋白相对表达量

图3 过表达SOX7对MG-63细胞VEGF和TGF-β1表达的影响
2.6 过表达SOX7抑制MG-63细胞Wnt/β-catenin信号通路
通过Western Blot检测转染pcDNA3.1-SOX7的MG-63细胞Wnt/β-catenin信号通路相关蛋白β-catenin、CyclinD1和Survivin表达,结果如图4和表6所示,pcDNA3.1-SOX7组β-catenin、CyclinD1和Survivin蛋白表达均明显低于空白组,差异有统计学意义(P<0.05)。

图4 过表达SOX7对MG-63细胞Wnt/β-catenin信号通路的影响
表6 过表达SOX7的MG-63细胞β-catenin、CyclinD1和Survivin的蛋白相对表达量
3 讨论
SOX7是SOX基因家族SOXF亚组成员之一,近年来的研究表明SOX7在结直肠癌、肺癌、肝癌等多种肿瘤中呈现低表达或不表达,其表达降低可促进肿瘤细胞生长[7-9],这提示在肿瘤发生发展中SOX7可能充当抑癌基因样作用。骨肉瘤中关于SOX7的研究较少,有研究显示SOX7是miR-664的一个靶基因,miR-664可通过靶向抑制SOX7增强骨肉瘤细胞的侵袭和迁移能力[5]。SOX7是否可影响骨肉瘤细胞凋亡、周期等生物学特性还未明确。本研究结果显示过表达SOX7的人骨肉瘤MG-63细胞SOX7表达明显升高,细胞活力明显降低,细胞发生G0/G1期阻滞,细胞凋亡率明显升高,提示SOX7同样可影响骨肉瘤细胞生长,可能是骨肉瘤诊疗的一个分子标记。
Wnt/β-catenin信号通路是Wnt信号的经典信号途径,人类多种肿瘤可发现Wnt/β-catenin信号通路的异常表达,如肝癌、乳腺癌、骨肉瘤等[10-13]。多项研究表明SOX7可通过调控胶质瘤、子宫内膜癌等肿瘤Wnt/β-catenin信号而抑制肿瘤细胞生长[14-15]。已有研究证实SOX7是Wnt/β-catenin/TCF信号的抑制因子,可通过与TCF竞争与β-catenin结合,从而发挥肿瘤抑制基因功能[6]。SOX7对骨肉瘤细胞生长影响是否与Wnt/β-catenin信号通路有关还未明确。本研究将过表达SOX7载体转染MG-63细胞,通过Western Blot检测Wnt/β-catenin信号通路关键分子β-catenin及下游与肿瘤发生密切相对的靶基因cyclinD1和Survivin表达,结果显示过表达SOX7可明显下调β-catenin、cyclinD1和Survivin表达,提示SOX7对骨肉瘤细胞活力、周期及凋亡影响机制可能与Wnt/β-catenin信号通路有关,这也与前人在其他肿瘤中的研究结果一致。
转化生长因子β1(TGF-β1)是一种多功能的蛋白多肽,是目前已发现的肿瘤诱导产生的最强的免疫抑制因子,TGF-β1过表达的肿瘤细胞侵袭能力更强,且预后不良[16-17]。已有研究表明TGF-β1可诱导多种肿瘤细胞生长[18],血管内皮生长因子(VEGF)可由大多数肿瘤细胞产生,对肿瘤血管生成有促进作用。多种肿瘤中VEGF呈现高表达,其表达与患者预后不良及无瘤生存期密切相关[19]。有研究表明抑制Wnt/β-catenin信号通路可明显降低VEGF表达[20],SOX7是否可影响TGF-β1和VEGF表达还未明确。本研究结果显示过表达SOX7可明显下调TGF-β1和VEGF表达,提示SOX7抑制骨肉瘤细胞生长机制之一可能是降低免疫逃逸。
综上所述,本研究将过表达SOX7的载体转染骨肉瘤MG-63细胞,通过CCK-8法、流式细胞仪、Western Blot分别检测过表达SOX7后的MG-63细胞活力、周期、凋亡及免疫抑制因子VEGF和TGF-β1表达及Wnt/β-catenin信号通路β-catenin、cyclinD1和Survivin表达,发现过表达SOX7可明显抑制MG-63细胞活力,阻滞细胞于G0/G1期,并诱导细胞凋亡,下调TGF-β1和VEGF及Wnt/β-catenin信号通路β-catenin、cyclinD1和Survivin表达,提示SOX7可能是骨肉瘤分子诊疗靶点之一,但目前SOX7对骨肉瘤的影响及其机制研究较少,还需进一步深入研究。
