压电通道蛋白1调节激素性股骨头坏死大鼠的成骨细胞凋亡与血管生成
2021-11-15何智军何伟洪志楠李子褀魏秋实陈晓俊颜新昊
何智军 何伟 洪志楠 李子褀 魏秋实 陈晓俊 颜新昊
临床上股骨头坏死(Osteonecrosis of Femoral Head,ONFH)与过度使用皮质类固醇密切相关,导致骨细胞凋亡和股骨头结构完整性丧失,并破坏骨血管组织[1-2]。压电通道蛋白1(Piezoelectric channel protein 1,Piezo1)是骨骼发育和成骨细胞分化所需的关键生物力传感器之一,其功能的发挥与Yes相关蛋白1(Yes associated protein 1,Yap1)密切相关[3]。Piezo1缺失可抑制成骨细胞分化,还改善在发育过程中的骨骼损失,因为Piezo1激活可诱导Yap1和转录因子β-连环蛋白(β-catenin)的表达并活化Yap1/β-catenin信号[4]。然而,Piezo1在皮质类固醇诱导ONFH中的表达情况及对骨细胞凋亡和血管生成的调节作用并不明确。
本研究通过类固醇诱导ONFH动物模型,研究沉默Piezo1对ONFH发生的影响并探讨其机制。
1 材料与方法
1.1 实验动物和分组处理
48只成年雄性Wistar大鼠(12周龄)购于广州中医药大学实验动物中心。 每只动物饲养环境为55 cm×35 cm×26 cm的有机玻璃笼子中,光照/黑暗周期12 h/12 h,温度24~25 ℃,湿度50%~55%。动物自由采食饮水。所有实验方案均遵守美国国立卫生研究院出版的《实验动物的护理和使用指南》,并经本院动物研究伦理委员会批准。
经过完全随机法将所有大鼠分为4组,各12只。1)类固醇激素+siP1组(STE+siP1组):在第1天用1.5 mL注射器于大鼠股骨远端松质骨富集处垂直骨面进针,髓腔注射siP1的体内转染复合物 50 μL,注射完2 h后和24 h后静脉注射1.8 mg/kg的LPS,并于第3,4,5,6,7天肌肉注射25 mg/kg甲基强的松龙,以促进股骨头坏死的发展,在第15天再次髓腔注射50 μL siP1体内转染复合物。2)类固醇激素+siNC组(STE+siNC组):将髓腔注射的siP1替换为等量的siNC的体内转染复合物,其余步骤和1)一致。3)类固醇激素组(STE组):在第1天用1.5 mL注射器于大鼠股骨远端松质骨富集处垂直骨面进针,髓腔注射生理盐水50 μL,注射完2 h后和24 h后静脉注射1.8 mg/kg的LPS,并于第3,4,5,6,7天肌肉注射25 mg/kg甲基强的松龙,以促进股骨头坏死的发展,在第15天再次髓腔注射50 μL生理盐水。4)对照组,与1)相同模式相同时间点,用0.9%生理盐水替代siPiezo1、LPS、甲泼尼龙。最后一次注射甲基强的松龙21 d后,断颈法处死动物。每组随机收集6只股骨头骨新鲜组织用于表达检测及制作切片。
在4%低聚甲醛-0.1 mol/L磷酸盐缓冲液(pH7.4)中固定24 h后,用10%乙二胺四乙酸-0.1 mol/L磷酸盐缓冲液(pH7.4)将股骨脱钙5周。脱钙后,将组织在梯度乙醇中脱水,包埋在石蜡中,沿冠状平面切成4 μm厚的切片。
1.2 试剂与耗材
Piezo1 siRNA(siPiezo1,siP1)和siRNA阴性对照(siNC)购于上海Genechem公司。甲基强的松龙购于大连辉瑞制药。杜氏改良培养液(Dulbecco’s Modified Eagle Medium,DMEM)/F-12、青霉素、链霉素均购于上海碧云天生物科技有限公司。人成骨细胞系(hFOB1.19)和脐静脉内皮细胞系(HUVEC)购于武汉普诺赛生物科技有限公司。重组人YAP1蛋白(Rh-YAP1)购于美国Abcam公司。β-catenin激活剂(ICG-001)购于美国MedChemExpress公司。PrimeScript RT试剂盒购于大连TaKaRa公司。PCR仪购于北京Eastwin生命科学公司。SYBR Green酶购于深圳富酶泰斯有限公司。Eco Real-Time PCR 系统购于上海Illumina公司。裂解缓冲液购于上海碧云天生物技术研究所。兔抗VEGFR2、CD3、Yap1、β-catenin抗体购于美国Abcam公司。末端标记(Terminal-deoxynucleoitidyl Transferase Mediated Nick End Labeling,TUNEL)试剂购于南京凯基生物技术公司。
1.3 细胞培养和处理
人成骨细胞系hFOB1.19和脐静脉内皮细胞系(HUVEC)均培养于DMEM/F-12培养基(20%的BSA、50 U/mL青霉素、50 μg/mL链霉素)。分别使用1.0 μmol/L重组人YAP1蛋白(Rh-YAP1)及2.5 μmol/L的β-catenin激活剂(ICG-001)处理hFOB1.19细胞和HUVEC细胞 24 h。收集细胞进行后续实验。
1.4 方法
1.4.1TUNEL分析 通过TUNEL检测试剂盒检测股骨头组织及所培养hFOB 1.19细胞的凋亡。骨组织中具有棕色核的细胞被评估为阳性。对hFOB 1.19细胞则统计TUNEL阳性细胞核的数量和总细胞核数。每组随机选择五个高倍视野(×200)来评估凋亡细胞的百分比(凋亡率)。
1.4.2实时定量PCR(qPCR) 用Trizol试剂从大鼠的股骨头中提取总RNA,检测260 nm(A260)的吸光度来定量RNA的浓度,通过确定A260/A280比例评估RNA的纯度。提取的总RNA(0.84 μg)用PrimeScript RT试剂盒在PCR仪上反转录合成cDNA(反转录PCR反应条件:16 ℃ 0.5 h,41 ℃ 0.5 h,85 ℃ 5 min,4 ℃循环),cDNA在4 ℃下保存。在PCR扩增之前,将cDNA保持在-20 ℃。qPCR反应在20 μL含有引物和SYBR Green酶反应体系的48孔光学PCR板中进行,条件为在94 ℃下预变性4 min,然后在94 ℃下再次变性45个循环,在60 ℃下退火2 min,并在75 ℃下延伸1 min。目标基因和内参基因GAPDH同板扩增,计算CT值,并使用2-ΔΔCt计算相对表达量。实验设置6个复孔,并且重复3次。实时定量PCR引物序列见表1。

表1 实时定量PCR引物序列
1.4.3蛋白质免疫印迹 组织样本:将股骨头磨成粉状,在冷的含苯基甲基磺酰氟的RIPA裂解缓冲液中匀浆。细胞样本:收集2组的HUVEC细胞,经过超声裂解。以上样品经过与上样缓冲液混合,沸水浴中煮5 min,然后在-20 ℃下保存直至用于电泳。用12%十二烷基硫酸钠聚丙烯酰胺凝胶分离蛋白质,并转移至聚偏二氟乙烯膜(Millipore Corporation,Bedford,马萨诸塞州,美国)上。用5%脱脂奶粉封闭2 h后,将膜与兔抗Piezo1(1∶500)、血管生成标志物VEGFR2(1∶600)、CD3(1∶800)和Yap1(1∶800)、β-catenin(1∶600)蛋白第一抗体在4 ℃下孵育过夜,而在25 ℃下与辣根过氧化物酶偶联的二抗(1∶50 000,武汉博斯特生物技术有限公司) 孵育2 h。使用Super Signal West Pico化学发光底物(Thermo Fisher Scientific Inc.,马萨诸塞州,美国)对膜上的蛋白进行显色,通过Geliance 200 Imaging System进行拍照。使用Image J软件分析条带灰度值。GAPDH作为的内参蛋白。实验重复3次。
1.4.4管状结构形成实验 将HUVEC细胞使用1.0 μmol/L Rh-YAP1及2.5 μmol/L的ICG-001处理24 h,并以5×103/孔的密度接种在Matrigel包被的12 孔板中,在37 ℃条件下孵育。16 h后,每孔拍摄5张照片。使用Image J软件测量细胞形成的总小管长度,每组实验3个复孔。
1.5 统计学方法
2 结果
2.1 股骨头坏死大鼠模型
Micro-CT和HE染色可见STE组软骨较薄,软骨表面粗糙不平,部分脱落,骨细胞和骨基质大量减少或丢失,皮质骨和骨小梁内出现坏死的细胞集落和大量空腔,对照(Control)组无上述特征(见图1)。
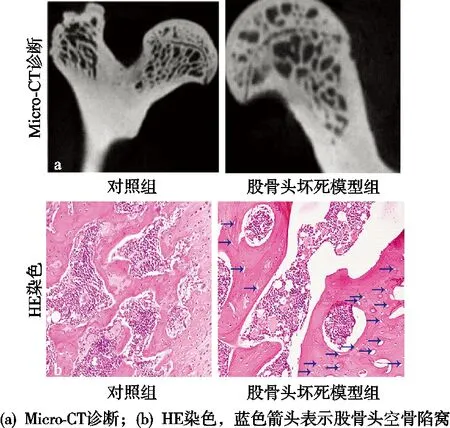
图1 STE诱导大鼠ONFH模型的建立
2.2 沉默股骨头坏死大鼠体内的Piezo1
与对照(Control)组比,STE组股骨头骨组织中的Piezo1的mRNA水平以及蛋白水平在3,6,12,24 h都上调,差异有统计学意义(P<0.01)。另外,与STE+siNC组比,STE+siP1组Piezo1的mRNA水平以及蛋白水平在3,6,12,24 h下调,差异有统计学意义(P<0.01),见图2。
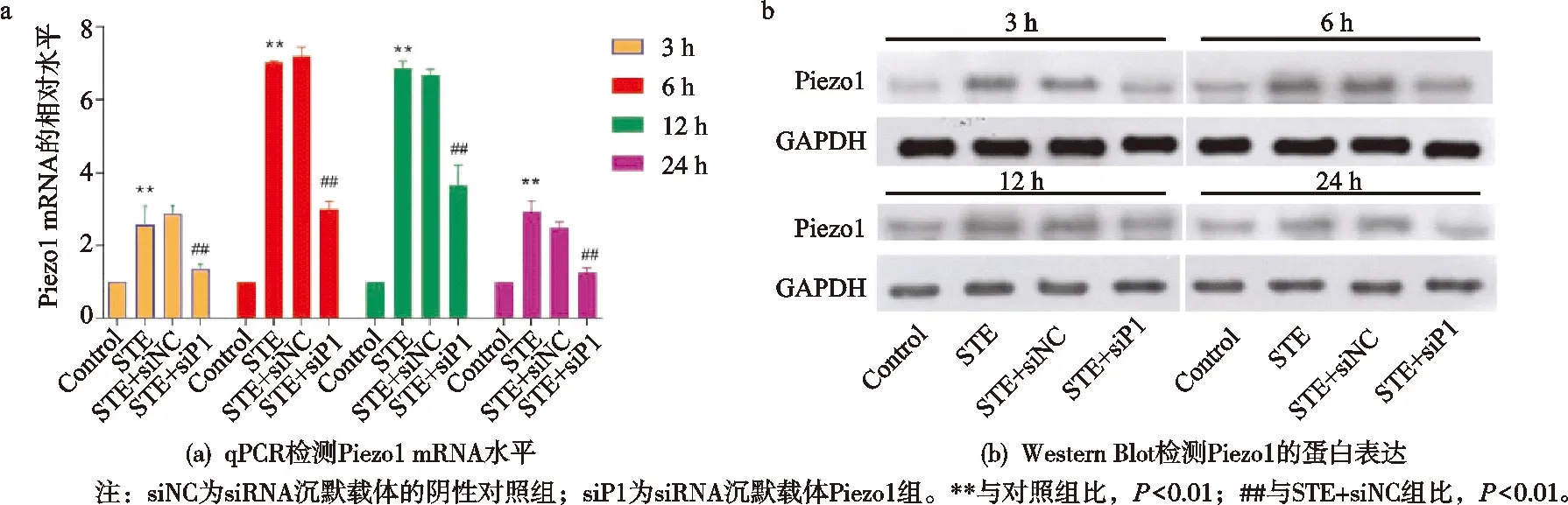
图2 Piezo1在股骨头坏死大鼠中的表达
2.3 Piezo1对股骨头坏死大鼠成骨细胞凋亡的影响
最后一次注射甲基强的松龙21 d后,对照组中TUNEL凋亡率为5.2%,而STE组凋亡率为90.3%(P<0.01)。与STE+siNC组的凋亡率(89.5%)比,STE+siP1组的凋亡率(37.9%)降低,差异有统计学意义(P<0.01),见图3。

图3 Piezo1沉默对 ONFH大鼠股骨头组织凋亡的影响 (×200)
与对照组比,STE组Bax和Caspase-3的mRNA表达水平升高,差异有统计学意义(P<0.01),而Bcl-2 mRNA的表达水平降低,差异有统计学意义(P<0.01)。与STE+siNC组比,STE+siP1组Bax和Caspase-3的mRNA表达水平降低,差异有统计学意义(P<0.01);而Bcl-2 mRNA的表达水平升高,差异有统计学意义(P<0.01),见图4。
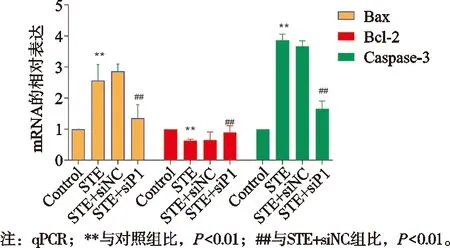
图4 Piezo1沉默对ONFH大鼠中凋亡标志基因的mRNA表达量的影响
2.4 微血管形态变化结果分析
与对照组比,STE组的股骨头内血管体积、血管表面积、血管厚度均降低(P<0.01);与STE+siNC组比,STE+siP1组股骨头内血管体积、血管表面积、血管厚度均增多(P<0.01),见图5。
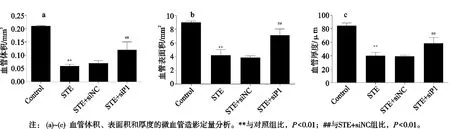
图5 Piezo1沉默对 ONFH大鼠股骨头组织血管量的影响
与对照组比,STE组VEGFR2和CD31表达水平降低,差异有统计学意义(P<0.01);与STE+siNC组比,STE+siP1组VEGFR2和CD31表达水平增高,差异有统计学意义(P<0.01),见图6。
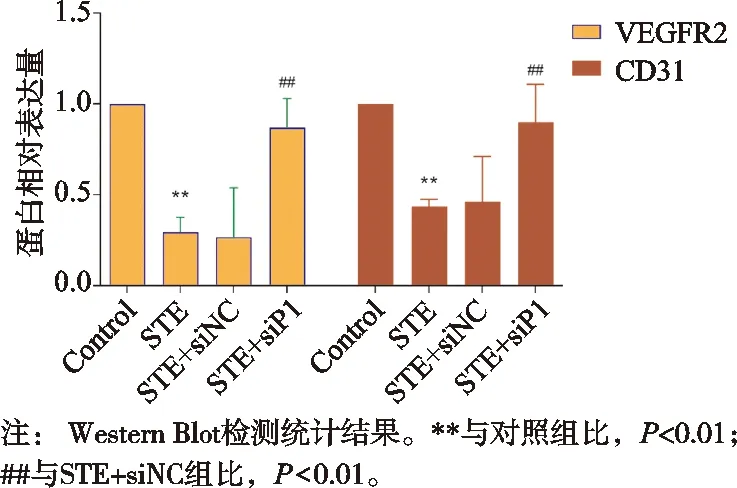
图6 Piezo1沉默对VEGFR2和CD31的蛋白表达量的影响
2.5 Piezo1沉默对股骨头坏死大鼠Yap1/β-catenin信号通路的影响
与对照组的股骨头骨组织中Yap1和β-catenin的蛋白(1.00±0.03,1.00±0.04)比,STE组(3.43±0.23,3.57±0.44)表达上调,差异有统计学意义(P<0.01);与STE+siNC组Yap1和β-catenin的表达(3.42±0.55,3.61±0.64)比,STE+siP1组(1.85±0.25,2.71±0.34)降低,差异有统计学意义(P<0.01),见图7。
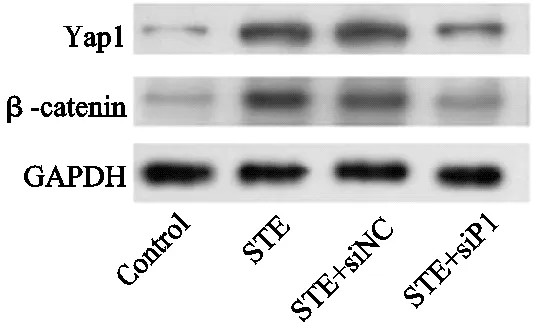
图7 Western Blot检测Piezo1沉默对Yap1和β-catenin的蛋白表达量的影响
2.6 激活Yap1/β-catenin信号通路促进成骨细胞的凋亡并抑制血管生成能力
与Ctrl(1.00±0.06,1.00±0.05)比,Rh-YAP1组中Yap1和β-catenin的蛋白水平(4.21±0.28,2.23±0.25)都增高,差异有统计学意义(P<0.01)。与对照(Ctrl)组(1.00±0.05)比,ICG-001组β-catenin(4.52±0.45)的蛋白表达上调,差异有统计学意义(P<0.01),见图8a。
与Ctrl组比,Rh-YAP1组和ICG-001组的细胞凋亡率增加,差异有统计学意义(P<0.01)。与Ctrl组比,Rh-YAP1组及和ICG-001组的HUVEC细胞管状结构形成长度均下调,差异有统计学意义(P<0.01),见图8(b,c)。

图8 激活Yap1/β-catenin信号通路促进成骨细胞的凋亡并抑制血管生成能力
3 讨论
类固醇可诱导ONFH[5],其诱导的细胞凋亡是关键原因之一[6]。本研究大鼠接受类固醇激素甲基强的松龙的连续注射后,ONFH模型成功建立。文献中类固醇所诱导ONFH的股骨头会出现明显的骨坏死,其特征在于STE组的骨小梁中有大量的空骨细胞陷落,没有发现明显的骨髓坏死,但发现ONFH的骨细胞凋亡率增加[6],本研究股骨部位骨细胞凋亡率也明显增加。Huang等[7]在类固醇诱导的ONFH患者被切除的股骨头部分也观察到骨细胞凋亡,认为糖皮质激素对股骨头的松质骨具有直接的细胞毒性作用,导致凋亡而不是单纯的坏死。Zheng等[6]在动物实验中也证明糖皮质激素可以促进成骨细胞和骨细胞的凋亡。凋亡性骨细胞的积累可能导致骨坏死,表明具有抑制骨细胞凋亡潜能的药物可能具有预防类固醇诱导的ONFH发生的能力。
Piezo1对于骨形成和调节骨吸收发挥关键作用[8-10]。因此,本研究对Piezo1在ONFH模型中的表达进行了检测,观察到Piezo1的mRNA水平以及蛋白水平在造模后3 h至24 h都上调,说明Piezo1很可能对激素性股骨头坏死有调节作用,而沉默Piezo1可以抑制股骨头坏死组织中骨细胞的凋亡。
Bax激活ONFH的骨细胞的凋亡,而Bcl-2选择性结合Bax的活性构象从而抑制其凋亡[11]。在本研究中,ONFH模型中沉默Piezo1,导致Bax mRNA明显降低,但Bcl-2 mRNA显著升高,表明沉默Piezo1可以通过抑制Bax提高Bcl-2从而抑制骨细胞凋亡。Caspase-3是另一种重要的凋亡相关蛋白,属于Caspase家族,在细胞内凋亡信号的诱导、转导和扩增中起着至关重要的作用。Caspase-3可被切割形成活性分子促进骨细胞凋亡[11]。在本研究中,ONFH模型中Caspase-3的mRNA升高,意味着由糖皮质激素诱导的骨细胞凋亡是由Caspase-3执行的,而沉默Piezo1可以明显抑制Caspase-3的表达,进一步表明Piezo1沉默后可抑制骨细胞凋亡。
局部缺血和骨坏死是股骨头骨坏死的核心病理机制。本研究结果发现,ONFH出现血管减少,且血管新生关键蛋白VEGFR2和CD3的表达量下调。而沉默Piezo1可促进大鼠ONFH模型中的股骨头局部血管的生成,包括血管的体积、表面积和血管厚度在内可观察到明显的增加,而且骨组织中VEGFR2和CD3蛋白水平明显上调。研究证明,Piezo1在胎盘血流敏感性中扮演决定作用,可调节胎盘血流[12],表明沉默Piezo1可能对ONFH的血管再生有帮助作用。
Yap1和β-catenin信号通路的协同作用已被证实[13-14],Yap1已被证实可以调节结直肠癌细胞的血管生成[15],而β-catenin信号在多种肿瘤模型和疾病中参与血管生成的调控,包括股骨头坏死动物[16-17]。在本研究实验结果中观察到,沉默Piezo1明显抑制Yap1和β-catenin的蛋白表达,从而抑制Yap1/β-catenin信号通路的活性。当使用Yap1的重组蛋白和β-catenin信号激活剂时,成骨细胞的凋亡率明显上调,而且HUVEC细胞的成管能力部分减弱。说明在成骨细胞中Yap1/β-catenin可能是Piezo1行使功能的关键信号路径,且沉默Piezo1通过抑制Yap1/β-catenin信号改善股骨头坏死导致的骨细胞凋亡和股骨头血管生成能力的缺失。
综上所述,沉默Piezo1通过抗成骨细胞凋亡和促血管生成预防类固醇诱导的ONFH,其潜在机制涉及Bax/Bcl-2、Caspase-3和Yap1/β-catenin信号通路,说明沉默Piezo1是治疗ONFH的一种潜在治疗方法。
