秦岭南麓中、东段小型兽类多样性及分布特征
2021-11-14赵文超赵依然卜书海
赵文超 李 琦 赵依然 卜书海
(1.陕西牛背梁国家级自然保护区,西安,710100;2.西北农林科技大学生命科学学院,杨凌,712100;3.西北农林科技大学林学院,杨凌,712100)
物种多样性是生物多样性在物种水平上的一种体现形式,包含2个最基本的参数,即物种丰富度和物种多度[1],其分布特征受经纬度、海拔等因素的影响较大[2-3]。不同海拔物种丰富度的分布呈现出不同形式。目前认为,物种丰富度的海拔分布格局主要包括4种形式,即单调递减格局、低海拔平台格局、低海拔峰值的单峰分布格局和中海拔峰值的单峰分布格局,其中单峰分布格局更为普遍[4]。影响物种丰富度海拔梯度格局的因素包括气候变量、空间面积、进化机制、种间作用,以及栖息地异质性等因素[5-6]。
研究物种多样性的分布特征及其维系机制是进一步探索物种多样性下降、分布区域变迁和物种灭绝因素的基础[7-8]。学者们分别对不同类群动物的海拔分布格局进行了研究,发现非飞行类小型哺乳动物和鸟类的物种丰富度随着海拔的升高呈单峰分布格局,而两栖类动物丰富度则呈现低海拔平台格局[9-11]。综合以往研究结果可以看出,物种丰富度的海拔分布格局与不同类群物种具有一定的关系,其中非飞行类小型哺乳动物(啮齿动物等)大部分呈中海拔峰值的单峰分布格局[12-13]。非飞行哺乳动物因其调查方法简单、便于野外实施,易获得可信的物种丰富度,因此常被用来研究物种多样性的分布特征[14]。
小型兽类(以啮齿类、食虫类为主)的物种多样性及分布特征影响着当地森林生态系统的质量和生物多样性,它们不仅可以捕食林中害虫、传播种子,而且还是多种掠食性动物的食物,在森林生态系统中起着重要作用[15-17]。由于体型限制,其物种多样性受海拔、植被类型及地形的影响较大[18],借助小型兽类来研究物种多样性与生态系统的相互关系更具代表性、准确性。
秦岭地区有着丰富的小型兽类资源,是研究物种多样性的理想场所。曾有学者[19]对秦岭地区小型兽类的分类、组成以及分布做过调查,但对其物种多样性方面的研究甚少。为此,本研究借助陷阱法以及铗日法对秦岭小型兽类物种,特别是食虫类多样性进行调查,旨在探索不同地理环境下小型兽类物种多样性及分布特征的差异,对秦岭地区小型兽类的保护与管理提供基础数据和科学参考。
1 研究区概况
南麓中段样地选在佛坪国家级自然保护区(33°33′—33°46′N;107°41′—107°55′E)内,该保护区位于陕西省佛坪县境内,总面积292.4 km2,是目前我国秦岭地区保存原始森林植被最完整、人为干扰最小的自然保护区。地貌特征显著,海拔980—2 904 m,相对高差1 924 m。保护区处于亚热带向暖温带过渡区的北缘,年均温度11.5℃,年降水量940—1 200 mm,雨季集中在6—9月,主要河流为金水河,属汉江支流。区内植被垂直带谱明显,从低到高依次为:海拔2 000 m以下的栎林带,海拔2 000—2 500 m的桦林带,2 500 m以上的针叶林带,山脊或峰顶多强风处,分布有斑块状小面积的亚高山灌丛和草甸。
南麓东段样地选在牛背梁自然保护区(33°47′—33°56′N,108°45′—109°4′E)内,该保护区地跨南北坡,总面积164.18 km2,海拔1 000—2 802 m,相对高差1 802 m,属暖温带半湿润气候区,年均气温9℃,年降水量850—950 mm,雨季集中在7—9月,水源丰富,有乾佑河、沣峪河、大东沟及西沟峡等溪流。区内植被垂直带谱较为明显:2 100 m以下为中低山典型落叶阔叶林,特征种为栓皮栎(Quercusvariabilis)和锐齿槲栎(Q.alienvar.acuteserrata);2 100—2 600 m为中山落叶阔叶小叶林带,优势林带为红桦(Betulaalbosinensis)林和牛皮桦(B.albosinensisvar.septentrionalis)林;2 600 m以上为亚高山寒温性针叶林带;海拔2 300—2 802 m为亚高山灌丛草甸或开阔裸露的山梁[20]。
2 研究方法
2.1 调查方法
基于秦岭南麓地形、气候特点及植被群落特征,将研究区域划分为4个垂直带:农耕带、栎林落叶阔叶林带、针阔混交林带以及针叶林带。在各林带内沿海拔梯度布设样地,秦岭中段佛坪保护区设置6个调查样地:农耕带(三官庙保护站,1 374—1 561 m)、栎林落叶阔叶林(蒸笼厂,1 787—1 838 m)、针阔混交林带(凉风垭河谷,2 063—2 218 m;凉风垭瞭望塔,2 204—2 339 m)和针叶林带(凉风垭山腰,2 395—2 528 m;凉风垭顶,2 544—2 643 m);秦岭东段牛背梁保护区设置6个调查样地:农耕带(红庙河河谷,1 113—1 358 m)、栎林落叶阔叶林带(老黑山乱石滩,1 479—1 602 m;娘娘庙,1 746—1 843 m)、针阔混交林带(八音台,1 987—2 143 m)和针叶林带(南天门,2 264—2 337 m;百步梯,2 474—2 747 m)。
每个样地中,随机选取3个小样方(200 m×100 m),间隔500 m。小样方内以10 m间隔布设鼠夹(12.0 cm×6.5 cm)及陷阱(直径15 cm,高度25 cm),放入花生米、昆虫作为诱饵。每个样方内连续放置2 d,次日早晨进行检查,收取捕获的标本、擦去鼠夹上沾有的血液并重新安置陷阱。第3日收取标本后将3个样方内的铗日及陷阱转移至下一样地。对捕获到的标本,记录捕获样地类型、陷阱号、标本体重及性别,并按照时间顺序进行编号。
2.2 数据处理及分析
依据体尺、体重、毛色及形体特征进行初步鉴定,外部特征难以鉴定的物种,剥离头骨,利用头骨特征及测量数据进行鉴定。利用多样性指数和均匀性指数来反映物种丰富度,以相对捕获率这一指标来体现物种多度,由于中、东段陷阱数有差异,所以对捕获率进行标准化。计算公式如下:
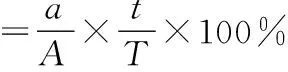
式中:a为单个区域内某物种个数;A为单个区域内所有物种个体数;t为单个区域内工作日;T为调查总工作日。
利用香农-维纳(Shannon-Wiener)多样性指数和Pielou均匀度指数来指示物种多样性和均匀度。
各指数计算公式如下:
香农维纳多样性指数(H′)计算公式为:
式中:S为样方内所有物种数;Pi为物种i的个体数量占群落中总个体数量的比例。
Pielou均匀度指数(J)计算公式为:
J=H′/log2S
式中:H′为香农-维纳多样性指数值;S为样方内所有物种数。
3 结果与分析
3.1 物种组成与区系特征
去除各种原因造成捕捉器械的损坏及丢失,共放置2 968个工作日,捕获小型兽类21种266只,平均捕获率7.11%,包括鼩鼱目(Soricomorpha)2科7属7种,啮齿目(Rodentia)4科8属14种(表1)。数量占比较高的种类有高山姬鼠(Apodemuschevrieri)、四川短尾鼩(Anourosorexsquamipes)、小纹背鼩鼱(Sorexbedfordiae)和中华姬鼠(Apodemusdraco);数量最少的有鼩鼹(Uropsilussoricipes)、灰麝鼩(Crociduraattenuata)、黄胸鼠(Rattustanezumi)和安氏白腹鼠(Niviventerandersoni),均为1只;有5只喜马拉雅水麝鼩(Chimarrogalehimalayica)、2只秦岭鼢鼠(Myospalaxrufescens)和1只隐纹花松鼠(Tamiopsswinhoei)系其他方法捕获。
捕获的小型兽类以东洋界物种为主(表1)。秦岭中段采集到13种,其中东洋种9种(69.23%),广布种3种(23.08%),古北种1种(7.69%);东段采集到18种,其中东洋种9种(50.00%),广布种7种(38.89%),古北种2种(11.11%)。捕获种类中,中国特有种有9种(42.90%),包括食虫类3种、啮齿类6种。
3.2 各植被带的物种分布特征
小型兽类物种在两区段同一植被带中的分布存在显著差异(表1)。
农耕带:中段捕获3种,以中华姬鼠为主,占该带捕获数的55.56%;东段捕获9种,高山姬鼠捕捉数最多,占该带捕获数的27.66%。黄胸鼠仅见于中段,麝鼹(Scaptochirusmoschatus)、喜马拉雅水麝鼩、岩松鼠(Sciurotamiasdavidianus)、甘肃仓鼠(Cansumyscanus)、高山姬鼠、褐家鼠(Rattusnorvegicus)和北社鼠(Niviventerconfucianus)仅于东段捕获。
栎林落叶阔叶林:捕获物种数最多。中段捕获8种,中华姬鼠捕获个体数最多,占该林带捕捉数的34.48%;东段捕获10种,大林姬鼠(Apodemuspeninsulae)捕捉数最多,占该林带捕捉个体总数的30.43%。鼩鼹、小纹背鼩鼱和针毛鼠(Niviventerfulvescens)仅于中段捕获,灰麝鼩、隐纹花松鼠、岩松鼠、甘肃仓鼠和大林姬鼠仅于东段捕获。
针阔混交林:中段捕获7种,以四川短尾鼩为主,占该带捕捉数的51.43%;东段捕获6种,以大林姬鼠为主,占该林带捕捉数的37.50%。小纹背鼩鼱、川西长尾鼩(Chodsigoahypsibia)和安氏白腹鼠仅于中段捕获,大林姬鼠、北社鼠和针毛鼠仅于东段捕获。
针叶林:中段捕获5种,四川短尾鼩捕获个体数最多,占该林带捕捉的50%;东段捕获9种,小纹背鼩鼱捕捉数最多,占该林带捕捉数的47.83%,占据优势地位。川西长尾鼩和四川短尾鼩仅见于中段,小纹背鼩鼱、隐纹花松鼠、甘肃仓鼠、高山姬鼠、大林姬鼠和大足鼠(Rattusnitidus)仅见于东段。
可以看出,中华姬鼠和四川短尾鼩为秦岭中段的常见物种;大林姬鼠、高山姬鼠和小纹背鼩鼱为秦岭东段的常见物种。
3.3 中、东段的物种分布差异
中段放置1 356个工作日,共捕获13种小型兽类,捕获个体87只,其中,针阔混交林捕获率最高(18.39%),农耕带捕获率最低(4.73%);栎林落叶阔叶林捕获物种数最多(8种),农耕带捕获物种数最少(3种)(表1)。捕获到啮齿类9种,鼠科(Muridae)占大部分(6种);食虫类4种,包括鼹科(Talpidae)的鼩鼹和鼩鼱科(Soricidae)的小纹背鼩鼱、川西长尾鼩和四川短尾鼩。
东段放置1 612个工作日,共捕获18种小型兽类,捕获个体179只。其中,农耕带捕获率最高(28.52%),针阔混交林捕获率最低(4.85%);栎林落叶阔叶林捕获物种数最多(10种),针阔混交林捕获物种数最少(6种)(表1)。捕获到啮齿类12种,鼠科最多(7种);食虫类6种,分别为鼹科的麝鼹、鼩鼱科的小纹背鼩鼱、川西长尾鼩、四川短尾鼩、喜马拉雅水麝鼩和灰麝鼩,其中麝鼹和大足鼠系牛背梁保护区首次记录。
中段相对捕获率最高的是四川短尾鼩(16.28%),其次为中华姬鼠(9.98%),鼩鼹、黄胸鼠、安氏白腹鼠、隐纹花松鼠和洮州绒鼠(Caryomyseva)的相对捕获率最低,均为0.53%;东段相对捕获率最高的是高山姬鼠(9.71%),其次为小纹背鼩鼱(6.67%),灰麝鼩相对捕获率最低(0.30%)。
3.4 物种多样性
中、东两区段内物种多样性存在明显的地理区域和林带差异。中段各群落小型兽类物种多样性指数由高至低依次是:栎林落叶阔叶林(2.600)、针阔混交林(1.940)、针叶林(1.573)和农耕带(1.392),均匀性指数依次是:农耕带(0.878)、栎林落叶阔叶林(0.867)、针叶林(0.787)和针阔混交林(0.691)。栎林落叶阔叶林的物种多样性最高,农耕带物种多样性最低而均匀性最高。
东段各群落小型兽类多样性指数由高至低依次是:栎林落叶阔叶林(2.995)、针叶林(2.292)、针阔混交林(2.234)和农耕带(0.937),均匀性指数依次是:栎林落叶阔叶林(0.902)、针阔混交林(0.864)、针叶林(0.723)和农耕带(0.296)。栎林落叶阔叶林的物种多样性和均匀度最高,而农耕带的物种多样性和均匀度皆为最低。
3.5 秦岭南麓小型兽类的海拔分布特征
绘制沿不同海拔高度小型兽类的多样性指数和均匀度指数分布曲线(图1),可以看出秦岭南麓中段小型兽类多样性的海拔分布格局呈现出较为明显的中海拔峰值,中海拔带的多样性比低海拔和高海拔带都高;而东段的海拔分布格局不明显,高海拔带的多样性指数也较高。
4 讨论
4.1 物种多样性
物种多样性沿山体海拔存在多种分布模式,包括随海拔上升单调递减的格局、低海拔稳定中高海拔逐渐降低的驼峰格局、低海拔平台中海拔峰值格局、中海拔峰值格局[4]。在全球范围内,非飞行性小型兽类的多样性在小尺度多样性格局中基本呈现中海拔峰值分布[21]。本研究探索了秦岭中、东段佛坪保护区以及牛背梁保护区内小尺度格局下的小型兽类物种多样性分布,结果表明,两保护区内小型兽类物种多样性的垂直分布均具有差异性,尤其是佛坪保护区内的物种多样性分布呈现明显的中海拔峰值格局,与秦岭平河梁[19]、武陵山[22]、夹金山[23]以及贡嘎山[24]等地区的小尺度小型兽类物种-海拔分布研究结果相一致,支持McCain等[4]提出的小型兽类海拔分布格局;相较而言牛背梁保护区的中海拔峰值格局显著性不强,海拔最高的针叶林带物种多样性达到第2峰值。究其原因,McCain等[5]认为当受到气候因子及山体效应(mountain mass effect)的影响时,非飞行性小型兽类中海拔峰值格局的峰值海拔会偏高于山体的海拔中值,大约在山体海拔的61%。调查结果显示,针叶林带出现的物种有90%左右在南天门捕获(2 264—2 337 m),其所在海拔位于山体海拔的60%,因此可能是由于该区域内山体效应以及各种气候因子导致物种多样性峰值向高海拔延伸,出现第2峰值。
在针阔混交林和针叶林类型中,中段捕获个体数最多的物种是四川短尾鼩,而东段捕获最多的是小纹背鼩鼱,说明在秦岭南麓不同区域内常见种存在差异,可以看出,不同动物在水平分布区域上存在差异。调查结果显示,不同生境中的物种多样性存在差异,秦岭南麓中段低海拔区植被类型栖息地的物种多样性指数处于低水平,中海拔区植被类型栖息地内的物种多样性指数较高。影响不同生境类型中的物种多样性的因素很多,这些因素综合作用于群落的物种多样性。植被类型的分布与海拔高度具有一定的协同关系,同时栖息地类型对哺乳动物的多样性也存在着一定影响,如栖息地内的食物资源[25],因而在不同植被类型和海拔高度上物种多样性会存在差异。海拔高度对植被的垂直分布影响与对哺乳动物物种组成的垂直分布的影响是类似的,这种哺乳动物的物种组成和植被组成的垂直分布特征的一致性反映出了动植物的协同进化特征[26]。
4.2 分布特征
通过对本次捕获的中、东段小型兽类海拔分布进行分析,可见两地段食虫类分布下限普遍较啮齿类高,中段捕获4种食虫类,除四川短尾鼩外其余3种分布下限均在1 500 m左右,显著高于同段分布啮齿类;东段捕获6种食虫类,除麝鼹和喜马拉雅水麝鼩外,其余4种分布下限均在1 300 m以上,亦显著高于同段分布的大多数啮齿类。食虫类多数种类喜栖息于山地森林中,耕种及砍伐活动导致低海拔地区森林大面积破坏,生境异质性降低,觅食、繁殖和避敌活动受到影响,此外,食虫类主要以地面昆虫为食,低海拔较强的人为干扰导致地面昆虫多样性降低[27]。因此,可能导致食虫类被迫向人为干扰小的高海拔林地迁移[28]。
对比中、东段物种分布海拔界限可以看出,四川短尾鼩、岩松鼠、洮州绒鼠、中华姬鼠、大林姬鼠和安氏白腹鼠等物种在秦岭中段分布下限显著较低;川西长尾鼩、四川短尾鼩、中华姬鼠、针毛鼠和秦岭鼢鼠等物种在秦岭中段分布上限显著较高,而鼩鼹、小纹背鼩鼱、岩松鼠、甘肃仓鼠和褐家鼠等物种的分布上限则在秦岭东段较高。秦岭中段佛坪保护区建立时间较早,此前基本未进行森林砍伐,人为干扰较小;秦岭东段的牛背梁保护区人为活动多,干扰大,植被破坏严重,导致多数小型兽类的分布下限上移[29]。
山体效应理论认为,由于山体基面高度不同,内部的山体基面高度往往高于外围。这种差异使得同一海拔上山体内部温度比外部高,进而导致山地垂直带谱的结构和分布产生差异,随之也影响到山体植被带的上限分布[30]。受山体效应影响,中段高海拔地区的气温相对较高,可能导致一些物种的分布上限上移,如川西长尾鼩、四川短尾鼩等物种在中段分布海拔上移较为显著,但是温度并非物种分布的唯一限制因素,地形、植被以及水源等环境因子对物种的分布亦有较强的影响,诸如鼩鼹、小纹背鼩鼱等在东段的分布上限较高。气温、地形、植被和水源对小型兽类分布上限的相对影响程度仍有待进一步研究。
4.3 捕获手段
相比之前利用铗日法在中段佛坪保护区的调查结果[31],本次中段调查利用陷阱辅以铗日法捕获的食虫类种类和数量明显较多。研究发现,单一的捕捉手段应用于小型兽类的调查具有一定片面性,无法全面地采集到所有小型兽类群体[32]。铗日法对于体型较大的小型兽类如安氏白腹鼠、社鼠等具有较为优越的捕获效率,而绝大多数食虫类由于体型小、体重轻,难以触发铗日开关,导致这类物种的捕获效率欠佳;陷阱法恰好弥补了这一缺点,小型食虫类进入陷阱后难以爬出,从而提高了捕获率[33-34]。因此利用铗日法与陷阱法相结合的捕捉手段可以更全面地捕捉小型兽类,降低物种多样性的偏差。
