蜕皮激素对黑化黏虫体液免疫的影响
2021-11-12张乃元张蕾程云霞等
张乃元 张蕾 程云霞等

中图分类号:S 433.4 文献标识码:A DOI:10.16688/j.zwbh. 2020225
外界微生物时刻威胁着昆虫的存活,因此免疫系统在昆虫的生长发育过程中至关重要。先天免疫系统是昆虫的主要免疫系统,主要由细胞免疫和体液免疫组成。体液免疫中酚氧化酶(phenol oxi-dase,PO》和溶菌酶(lysozyme)是免疫关键酶,PO和溶菌酶活性是免疫反应的重要指标。昆虫免疫系统受内分泌系统调节,内分泌激素在昆虫的免疫反应中起重要的调节作用。昆虫体内主要的内分泌激素是保幼激素(juvenile hormone,JH)和蜕皮激素(molting hormone,MH),它们在昆虫的生长过程中不仅可以联合调控昆虫的变态、迁飞和繁殖,而且在昆虫的免疫系统中也有非常重要的调控功能。JH是昆虫咽侧体分泌的一种生长调节剂,能够保持幼虫状态、抑制蜕皮和变态,调控昆虫生命周期中每一个重要发育阶段的转变,在昆虫发育的临界期高浓度JH会阻碍变态提前发生。蜕皮激素的主要成分是20羟基蜕皮酮(20-hydroxyecdys-one,20E),主要作用是激发昆虫的蜕皮活动,调节昆虫所有的幼虫态、蛹以及成虫的转换。在针对黑腹果蝇Drosophila melanogaster的研究中发现,注射20E后,黑腹果蝇的细胞吞噬功能和PO活性得到了增强。棉铃虫Helicoverpa armigera在注射20E后,细胞吞噬功能提高,细胞免疫明显增强。JH和MH对昆虫细胞免疫既有正调控作用,也有负调控作用,并可通过增强或抑制PO活性来调节昆虫体液免疫。
黏虫是严重威胁我国主要粮食作物生产安全的重大农业害虫。由于近年来黏虫在我国局部地区常常暴发,对粮食生产安全带来重大威胁。暴发的黏虫幼虫发生黑化现象已成常态,其免疫能力显著增强。但这种免疫能力是否受到内分泌激素的影响还不清楚,关于黑化黏虫内分泌激素MH如何调控免疫反应未见报道。因此,本研究旨在明确黑化黏虫6龄幼虫被球孢白僵菌Beauveria bas-siana侵染后其内分泌激素MH的含量变化。注射20E稀释液后,明确被白僵菌侵染的黑化黏虫幼虫死亡率的变化以及体液免疫中PO和溶菌酶活性的变化,揭示黑化黏虫体液免疫和内分泌调节机制间的关系。
1材料与方法
1.1材料
供试菌株:球孢白僵菌Beauveria bassiana菌株由中国农业科学院植物保护研究所蔬菜害虫组提供。使用0. 01%吐温80溶液配制105、5×105、106、5×106、107cfu/mL和108cfu/mL的球孢白僵菌孢子悬浮液进行预试验,经SPSS-Probit回归模型分析处理,球孢白僵菌对黑化黏虫的LC50约为106cfu/mL,本试验所有黏虫感染球孢白僵菌的浓度均为106cfu/mL。
供试昆虫及饲养:供试黏虫采集于吉林省四平市公主岭市田间。在饲养瓶(直径9 cm×高13 cm)中高密度诱导饲养,饲养密度为10头/瓶。饲养方法参照郭栋、吕伟祥等室内人工饲养方式,每日更换30~50cm高的玉米苗直至幼虫停止取食。饲养温度设置为(23±1)℃,湿度设置为(70±5)%,光周期设置为L∥D=14 h∥10h。
1.2球孢白僵菌和外源蜕皮激素注射方法
1.2.1黑化黏虫幼虫白僵菌注射方法
根据预试验的结果,球孢白僵菌浓度106cfu/mL注射后可以激活黑化黏虫6龄幼虫的免疫反应,且有近半数黏虫幼虫可以完成世代发育。将蜕皮24h内的黑化黏虫6龄幼虫置于冰上冷冻麻醉15 min,幼虫静止后,使用微量注射器吸取106cfu/ml_的球孢白僵菌孢子悬浮液1μL,注入幼虫第3与第4腹足之间的腹部。处理对照组注射0. 01%吐溫80溶液。每个处理组30头虫,每个处理重复3次。幼虫伤口闭合并且晾干后,放入装有新鲜玉米幼苗的饲养瓶中继续饲养。
1.2.2黑化黏虫幼虫20E稀释液注射方法
取20 mg 20E加入1mL无水乙醇充分溶解混匀后,得到20μg/μL的20E母液。用无水乙醇对20μg/μL的20E母液2倍梯度稀释,得到浓度为1. 25、2.5、5、10μg/μL的20E稀释液并进行编号。蜕皮24h内的黑化黏虫6龄幼虫注射球孢白僵菌孢子悬浮液后,立刻对其注射20E液。对照组为注射球孢白僵菌孢子悬浮液后立刻注射无水乙醇。每个处理组30头虫,每个处理重复3次。注射方法同1.2.1。
1.3免疫指标测定方法
1.3.1 MH含量测定方法
在注射球孢白僵菌后8、24、48、72h和96h分别取对照组(注射吐温80)以及处理组(被白僵菌侵染)的黏虫血淋巴样品,每次采集3头黏虫,每头幼虫采集血淋巴约50μL,每头黏虫血淋巴样品重复测试3次。使用昆虫蜕皮激素(MH)试剂盒测定MH含量。在96孔酶标板中设空白孔、标准孔和样品孔。标准孔中加入稀释为10、20、40、80、160ng/L的MH标准品,样品孔中加入待测样品及样品稀释液后在预热至37℃的水浴锅中温育30 min。温育后弃去板中的液体,使用洗涤液洗板并拍干。加入酶标试剂温育30 min并使用洗涤液洗涤拍干。加入显色试剂在37℃的水浴锅避光显色15 min。避光显色后加入终止剂终止反应,用酶标仪在450 nm波长下测定吸光度(OD),分别通过标准曲线计算样品中的MH含量。
1.3.2被球孢白僵菌侵染的黑化黏虫注射20E后幼虫死亡率测定
每天观察记录对照组(被白僵菌侵染后仅注射无水乙醇)的黑化黏虫6龄幼虫,以及处理组(被白僵菌侵染后)分别注射1. 25、2.5、5、10、20 μg/μL20E稀释液的黑化黏虫6龄幼虫的存活情况,统计幼虫死亡数量,直至幼虫化蛹。统计分析黑化黏虫幼虫的校正死亡率。每个处理组30头虫,每个处理重复3次。选取对黑化黏虫死亡率影响最显著的20E稀释液进行血淋巴免疫酶活性测定试验。
1.3.3血淋巴PO和溶菌酶活性的測定
处理组在注射球孢白僵菌孢子悬浮液后注射5μg/μL20E稀释液。处理对照组在注射球孢白僵菌后注射无水乙醇。另设空白组不作处理以验证注射球孢白僵菌的黑化黏虫6龄幼虫被侵染成功。在被白僵菌侵染后8、24、48、72 h和96 h取黏虫血淋巴样品,每次采集3头黏虫,每头采集约50μL,每头黏虫血淋巴重复测试3次。使用酶联免疫试剂盒测定溶菌酶和PO活性。试剂盒操作方法参照1.3.1。
1.4数据统计与分析
数据采用Excel进行统计,所有数据采用平均值±标准误(mean±SE)表示,应用统计软件SPSS24.0对数据进行分方差分析。用单因素方差分析(ANOVA)Tukeys HSD和独立样本t测验进行差异显著性分析。
2结果与分析
2.1白僵菌侵染对黑化黏虫幼虫体内MH含量的影响
从图1可以看出,黑化黏虫幼虫的MH含量随侵染后时间的增加呈先降低再升高的趋势,总体上呈现侵染组高于对照组(侵染后24h除外)。对照组(注射吐温-80) MH含量在处理后8~72 h呈下降趋势。在处理后96 h时MH含量显著升高(F4,40=25. 131,P<0. 05),达到120. 00 ng/l_。与对照组相比,注射球孢白僵菌后,在侵染初期8h时黑化黏虫幼虫的MH含量达到136. 96 ng/L,显著高于对照组的95. 25 ng/L(t=-9.028,fg=16,P<0. 05),是对照组的1. 44倍。在侵染中期24 h时,MH含量低于对照组,仅为54. 54 ng/L。48h时MH含量高于对照但无显著差异(t=-1.112,df=16,P>0. 05)。而在侵染后期72 h(t=-4.510,df=16,P<0. 05)、96h(t=-6.889,df=16,P<0. 05)时,黑化黏虫幼虫的MH含量显著增加,并且显著高于对照组。在侵染后96h时达到最大值197. 79 ng/L,是对照组的1. 65倍。
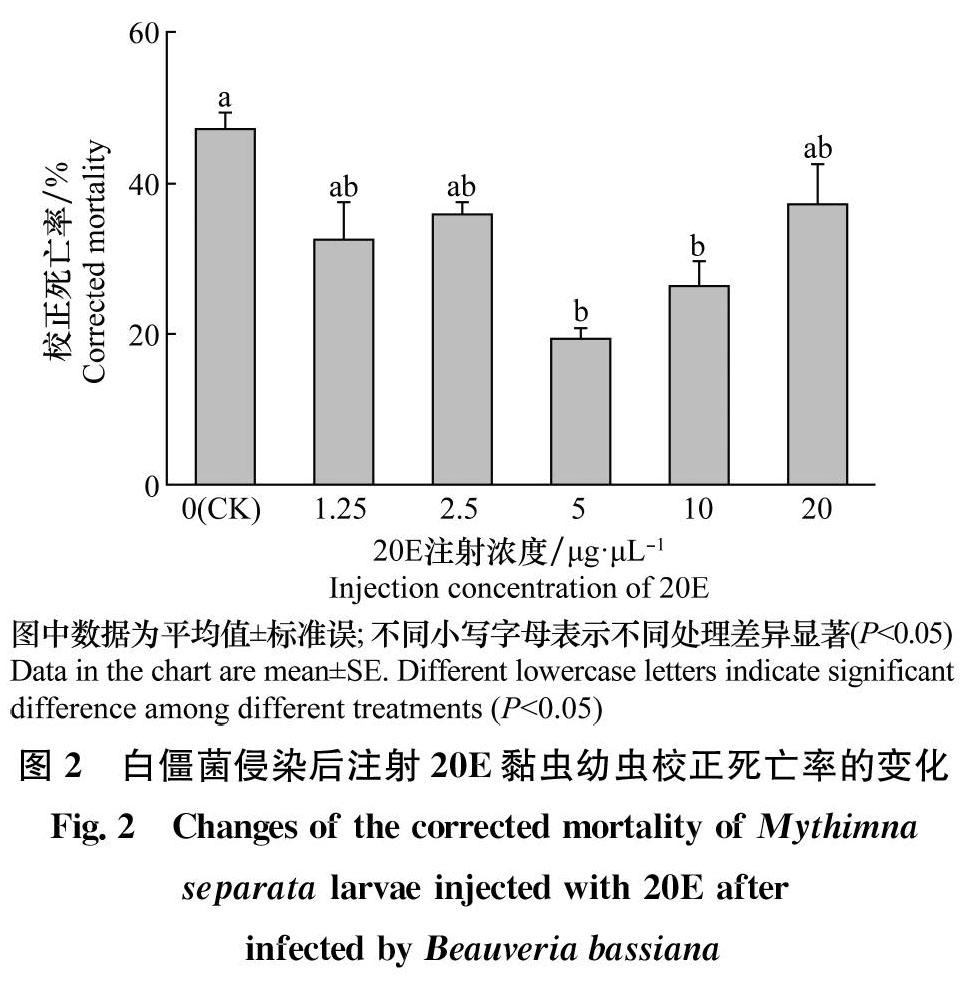
2.2注射20E对被菌侵染的黑化黏虫死亡率的影响
由图2可以看出,注射球孢白僵菌的黑化黏虫幼虫在注射不同浓度20E后,其校正死亡率有不同程度的下降。注射1. 25、2.5μg/μL和20μg/μL的20E后黑化黏虫的校正死亡率有所降低,但与注射无水乙醇的对照组相比,差异不显著(F5,12=2.909,P>0.05)。注射5μg/μL和10μg/μL的20E后黑化黏虫的校正死亡率显著降低(F5,12=2.909,P<0. 05),分别为19. 28%和26. 51%。供试的5个浓度中,5μg/μL的20E处理后被侵染的黑化黏虫的死亡率显著低于对照组(F5,12=2.909,P<0.05)。这说明注射20E有利于黏虫免疫能力增强。
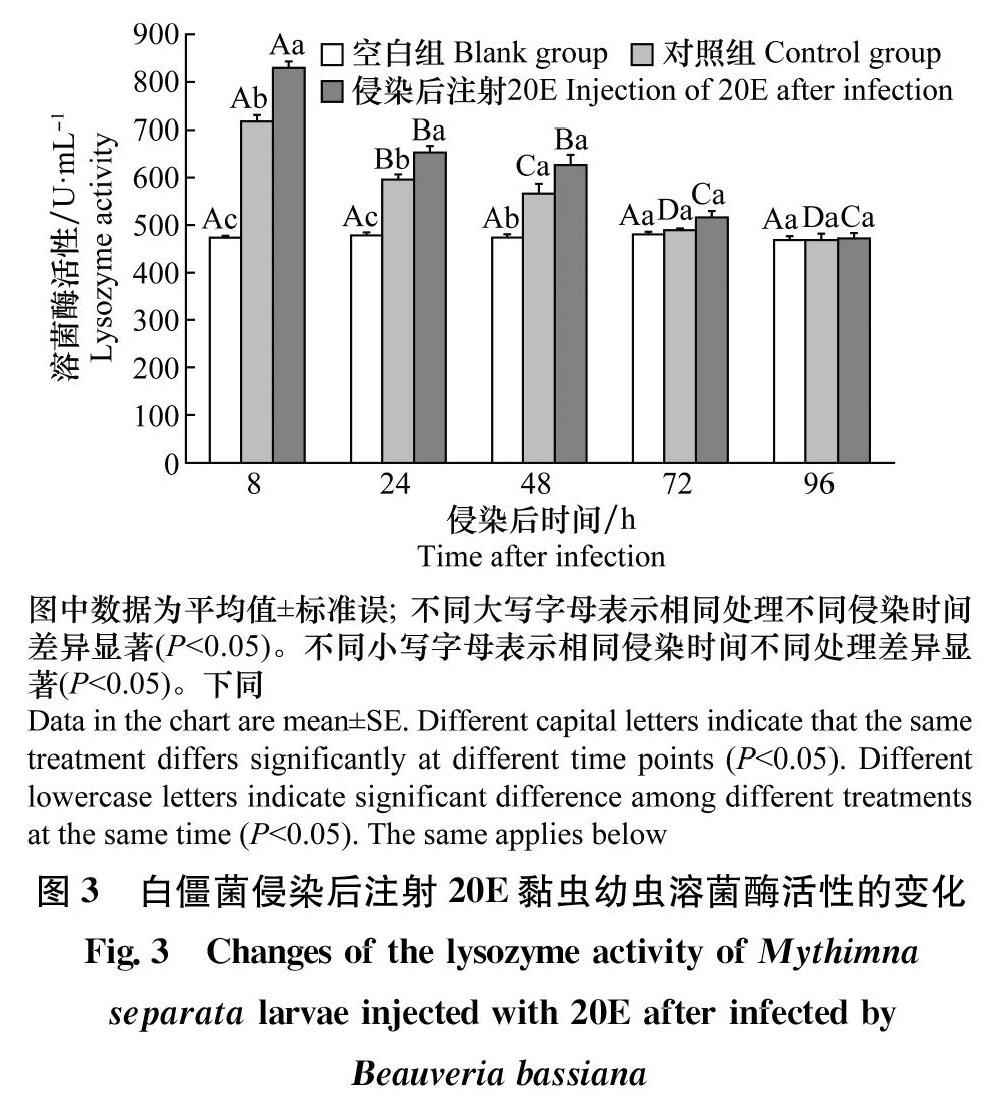
2.3注射20E对被茵侵染的黑化黏虫幼虫溶菌酶活性的影响
从图3中可以看出,空白对照组(不作处理)溶菌酶活性较低,在不同时间其活性无显著差异(F4,40=0. 366,P>0. 05)。对照组(被球孢白僵菌侵染后仅注射无水乙醇)被球孢白僵菌侵染后,体液免疫激活,溶菌酶活性增加。被球孢白僵菌侵染后注射20E的处理组免疫反应增强,溶菌酶活性高于对照组。注射20E的处理组的溶菌酶活性与对照组变化趋势相同,在侵染后随时间增加溶菌酶活性降低。对照组溶菌酶活性在被侵染后8h最高,达到最大值717. 61 U/mL,随后溶菌酶活性开始下降,随侵染后时间的增加,溶菌酶活性逐渐趋于稳定。经20E诱导的黑化黏虫幼虫的溶菌酶活性在8h时最高,并显著高于其他时间(F4,40=97. 262,P<0. 05)。被白僵菌侵染后8h(F2,24=386. 35,P<0. 05)和24 h(F2,24=128. 323,P<0. 05)时20E处理组溶菌酶活性显著高于对照组,在侵染48h之后,注射20E的处理组的溶菌酶活性也逐渐回到空白组基线水平。
2.4注射20E对被菌侵染的黑化黏虫幼虫PO活性的影响
从图4中可以看出,空白组PO活性较低,在不同时间无显著差异(F4,40=2. 885,P>0. 05)。处理对照组在注射球孢白僵菌后,球孢白僵菌激活了黑化黏虫的体液免疫,其PO活性增加。24h时PO活性达到最大值132. 25 U/mL。随被侵染时间的增加,其PO活性显著降低(F4,40-225. 340,P<0. 05)。注射球孢白僵菌后又注射20E的处理组,黑化黏虫经20E诱导后免疫反应增强,PO活性高于对照组。在侵染后8 h(F2,24=250. 345,P<0. 05)、24 h(F2,24=235. 849,P<0. 05)和48 h(F2,24=121. 385,P<0. 05)20E处理组PO活性均显著高于对照组,并在侵染后8h达到最大值156. 83 U/mL。
3讨论
昆虫种类繁多、形态各异,在长期的进化过程中,作为地球上数量最多、分布最广的动物群体,逐渐形成了独特的免疫机制。神经和内分泌系统对昆虫的免疫功能起到主要的调节作用。昆虫体内的蜕皮激素(MH)已被证明在发育、变态、生殖等方面具有重要作用,并且对昆虫的先天免疫反应有重要调控作用。蜕皮激素的主要成分20羟基蜕皮酮(20E)可以激活细胞免疫和体液免疫来对抗外源入侵微生物,而外源微生物的刺激也会通过3脱氢蜕皮激素-3β-还原酶促进20E滴度升高。本试验通过对黑化黏虫6龄幼虫注射球孢白僵菌来测定黑化黏虫幼虫体内的MH含量,发现被侵染后的黑化黏虫体内的MH含量出现显著变化。MH含量在侵染后高于对照组,侵染后8、72 h和96 h尤为明显。其原因是被球孢白僵菌侵染后,20E可以迅速激活免疫反应来对抗外源入侵微生物。这说明MH在黑化黏虫体液免疫反应中起到重要的调节作用。
近年来的研究发现,昆虫体液免疫可以受20E调控,酚氧化酶(PO)和溶菌酶等保护昆虫免受伤害。酚氧化酶是昆虫体液免疫中非常重要的免疫因子。在昆虫表皮的硬化和黑化的过程中酚氧化酶具有重要作用。在昆虫受到外源病原物的刺激时酚氧化酶同样具有重要作用。酚氧化酶活性是衡量昆虫体液免疫的重要指标之一。黑腹果蝇注射MH后,其体内的PO活性增强。针对埃及伊蚊Aedes aegypti的研究发现,鉴定到的一条PO基因序列中有2个可以结合20E受体复合物EcR/USP的顺式作用元件,20E的刺激能上调PO的转录表达。本试验结果与上述试验结果一致。通过对球孢白僵菌侵染后的黑化黏虫6龄幼虫注射20E稀释液来测定黑化黏虫血淋巴中的酚氧化酶活性。黑化黏虫6龄幼虫被球孢白僵菌侵染后,经20E稀释液诱导其PO活性显著增强,幼虫的免疫水平因外源20E诱导而提高。未经20E稀释液诱导的对照组黑化黏虫幼虫的酚氧化酶活性显著低于经20E稀释液诱导的试验组。这表明20E处理组的黑化黏虫幼虫的体液免疫功能明显增强。
在昆虫的非特异性免疫中,溶菌酶同样是非常重要的免疫因子。溶菌酶通过昆虫的脂肪体分泌释放到血淋巴中。昆虫在受到外界环境刺激时,溶菌酶的活性急剧增加。当外界的病原物进入昆虫体内后,病原体细胞壁会在溶菌酶的作用下破裂并使内容物溢出,最终病原细胞破裂死亡。针对棉铃虫的研究发现,棉铃虫20E滴度增高,脂肪体中溶菌酶基因的转录表达上调。针对东亚飞蝗Lo-custa migratoria的研究发现,溶菌酶基因在转录水平上可被20E显著上调。本试验结果与上述试验结果一致。本试验通过对白僵菌侵染后的黑化黏虫6龄幼虫注射20E稀释液来测定黑化黏虫血淋巴中的溶菌酶活性。试验中被侵染的黑化黏虫幼虫在注射20E稀释液后,溶菌酶活性的变化与酚氧化酶活性的变化趋势大致相同。这同样表明20E对黑化黏虫幼虫的体液免疫功能有增强作用。
综上所述,对黑化黏虫内分泌激素含量的分析表明,黑化黏虫6龄幼虫被球孢白僵菌侵染后,MH的分泌显著提高。对酚氧化酶和溶菌酶活性的分析表明,在注射20E稀釋液后,黑化黏虫幼虫的酚氧化酶活性和溶菌酶活性显著提高。本试验证明,黑化黏虫6龄幼虫在受到外源病原微生物侵染后,虫体通过内分泌激素对免疫系统进行调控从而增强免疫反应。本文试验结果对黑化黏虫免疫机制和内分泌调节机制间的关系提供了一定的理论依据,对揭示黑化黏虫、防御能力和免疫应答三者间联系具有重要意义。
