枯草芽孢杆菌JY24菌株产β-D-葡萄糖醛酸苷酶发酵工艺优化及催化转化
2021-10-25高书锋孔利华曾发姣王升平周小玲刘惠知
高书锋,孔利华,雷 平,曾发姣,王升平,龚 平,周小玲,刘惠知
(湖南省微生物研究院,长沙 410009)
β-D-葡萄糖醛酸苷酶(β-D-glucuronidase,GUSB),是一种糖苷类水解酶,多属于糖基水解酶家族2和79,能催化各类含β-D-糖苷键的化合物水解。当前优化微生物菌株产该酶文献报道较少,Chouiter等[1]对树舌灵芝产β-D-葡萄糖醛酸苷酶培养基和补料方式进行优化,发酵14 d酶活提高5倍,达1.09 U/mL;张斌等[2]对毕赤酵母高效表达重组β-D-葡萄糖醛酸苷酶诱导方法进行了研究,发酵5 d高效表达GAMG的酶活达1 970 U/mL;郭晓晓等[3]对产紫青霉β-D-葡萄糖醛酸苷酶的诱导表达进行了研究,初始葡萄糖耗尽时,继续诱导3 d和发酵2 d,酶活达2 356 U/mL,提高近3倍;吴文婷[4]对ChaetomiumglobosumS108菌株产β-D-葡萄糖醛酸苷酶工艺进行了研究,发酵4 d酶活达900 U/mL;Ku等[5]对德氏乳杆菌Rh2产β-D-葡萄糖醛酸苷酶的66种培养基优化筛选,厌氧发酵18 h,相对酶活提高8倍。上述优化微生物菌株产该酶文献报道所用菌种多属真菌,代谢产酶周期较长,为4 d至14 d,而德氏乳杆菌Rh2虽为细菌菌株,但产酶发酵方式为厌氧发酵,实际生产有一定困难。当前有关好氧细菌产该酶的发酵工艺优化研究尚未见报道,加之细菌具有代谢产酶周期短的优势,因此,有必要针对本实验室前期试验从健康鸡肠道选育的产β-D-葡萄糖醛酸苷酶的饲用枯草芽孢杆菌JY24菌株,开展产酶发酵工艺优化,为后续工作中提升黄芩苷转化率和转化效率提供借鉴和理论依据。
1 材料与方法
1.1 材料
1.1.1 菌种
枯草芽孢杆菌(Bacillussubtilis)JY24菌株,湖南省微生物研究院动物营养与疫病防控室分离选育保存。
1.1.2 培养基
种子培养基:LB培养基;基础发酵培养基:淀粉8 g、蛋白胨10 g、磷酸二氢钾1 g、七水硫酸镁0.5 g、氯化钠5 g、黄芩苷诱导物1 g、蒸馏水1 L,pH 7.0~7.2。
1.1.3 主要试剂和仪器
标准品:对硝基酚-β-D-葡萄糖醛酸苷,Sigma Aldrich(>99%,质量分数);对硝基苯酚,Dr.Ehrenstorfer GmbH(99.4%,质量分数)。TGL20M-II冷冻离心机、Tu-1810分光光度计、ZQZY-AS9振荡培养箱、LRH400A生化培养箱和Agilent1260高效液相色谱仪。
1.2 方法
1.2.1 β-D-葡萄糖醛酸苷酶酶活检测
(1)标准曲线绘制。称取0.347 8 g对硝基酚,PBS定容至100 mL,摇匀,吸取4 mL,PBS定容至100 mL,即储备液。稀释储备液为10~100 μmol/L系列标准液,检测OD405,选择合适梯度绘标准曲线。(2)酶活检测。参照文献[6]进行。酶活力单位:在pH 7.0、35 ℃条件下,每小时释放1 μmol对硝基苯酚所需酶量为1个活力单位,记为U。酶活单位(U/mL)=对硝基酚释放量(μmol)/保温时间(h)/被检液量(mL)。
1.2.2 发酵条件优化
发酵时间设9个处理:6、12、18、24、30、36、42、48和54 h;转速设4个处理:150、180、210和240 r/min;装液量设5个处理:10%、15%、20%、25%和30%;初始pH值设5个处理:6.0、6.5、7.0、7.5和8.0;温度设6个处理:29 ℃、31 ℃、33 ℃、35 ℃、37 ℃和39 ℃;接种量设5个处理:1%、2%、3%、4%和5%;上述不同处理均设3次重复。初始条件:采用三角瓶(500 mL)摇瓶发酵,接种量3%、初始pH 7.0、装液量20%、温度35 ℃、转速220 r/min。分别测定不同发酵条件下各处理3次重复发酵液的酶活性,优化发酵条件。
1.2.3 培养基成分优化
碳源、氮源和磷源种类筛选:计算基础发酵培养基碳源、氮源和磷源中碳、氮和磷含量,分别用6种碳源(葡萄糖、蔗糖、淀粉、红糖、糖蜜和麦麸)、7种氮源(蛋白胨、酵母粉、酵母膏、黄豆粉、牛肉膏、硝酸钾和硫酸铵)和3种磷源(磷酸二氢钾、1/2磷酸二氢钾+ 1/2磷酸氢二钾、磷酸氢二钾)替换,优化碳、氮和磷源种类。无机盐种类筛选:去除基础发酵培养基所有无机盐,分别添加8种0.002 mol/L的无机盐(硝酸钾、氯化钠、氯化钙、七水硫酸镁、一水硫酸锰、三氯化铁、七水硫酸亚铁和五水硫酸铜),CK不添加,优化无机盐种类。表面活性剂、生长因子种类和诱导物浓度筛选:表面活性剂设5个处理:PEG-400、Tween-20、Tween-80、TritonX-100和SDS,SDS添加浓度为1.0 g/L,其他添加浓度为1.0 mL/L,CK不添加;生长因子设3个处理:玉米浆、糖蜜和麦芽汁,添加浓度为2.0 mL/L,CK不添加;黄芩苷诱导物添加浓度设6个处理:0(CK)、0.5、1.0、1.5、2.0和2.5 g/L,优化表面活性剂、生长因子种类和诱导物浓度。上述各处理均设3次重复,分别测定不同培养基成分各处理3次重复发酵液的酶活性,优化培养基成分。
1.2.4 培养基组成优化
据单因素试验结果,选出最佳碳源蔗糖、氮源酵母膏、磷源 (1/2磷酸氢二钾和1/2磷酸二氢钾)、2种主要无机盐(氯化钙和硝酸钾)、表面活性剂Tween-20和生长因子玉米浆,设计7因素3水平(表1)正交试验,选用L18(37)正交表进行试验,重复3次,分别测定各组合3次重复发酵液酶活性,结果进行直观和方差分析,优化培养基组成和酶活性。各正交试验组合均添加1.5 g/L黄芩苷诱导物。
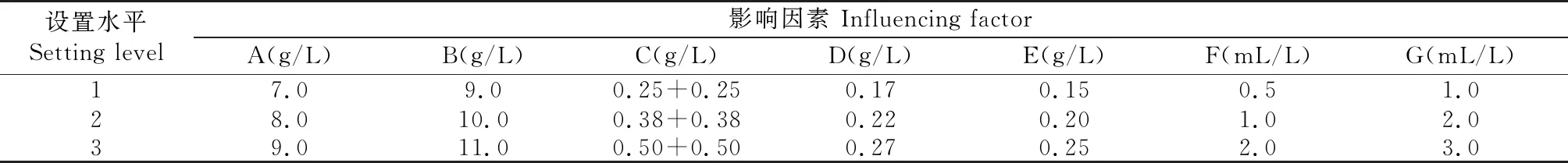
表1 培养基组成正交试验因素和水平设计Table 1 The factors and levels for soforthogonal test of the fermentation medium
1.2.5 酶催化黄芩苷转化初步试验
(1)酶催化黄芩苷转化试验方法。采用试验最优产酶培养基,添加50 g/L 80目黄芩粉,在试验最佳产酶条件下,进行β-D-葡萄糖醛酸苷酶催化黄芩苷转化试验,于发酵0 h和72 h分别检测发酵液黄芩苷和黄芩素含量,计算黄芩苷转化率,重复3次。黄芩素转化率=[(发酵72 h黄芩素含量-发酵0 h黄芩素含量)×1.651 7/发酵0 h黄芩苷含量]×100%(1.651 7为黄芩苷和黄芩素摩尔质量之比)。(2)黄芩苷、黄芩素含量检测方法。采用HPLC法,取适量发酵液,4 ℃、10 000 r/min离心10 min,上清液用色谱级甲醇进行梯度稀释,超声萃取30 min,0.22 μm 聚偏氟乙烯微孔滤膜(F型)过滤,Agilent1260高效液相色谱检测。色谱条件:色谱柱Eclipse XDB-C18 (250 mm×4.6 mm,5 μm);检测波长275 nm;柱温30 ℃;流速1.0 mL/min;进样量10 μL;流动相为甲醇∶水∶磷酸(55∶45∶0.04)。黄芩苷浓度在2~100 μg/mL范围内线性关系良好,黄芩苷保留时间(tR=6.024 min);黄芩素浓度在5~100 μg/mL范围内线性关系良好,黄芩素保留时间(tR=8.317 min)。
1.3 数据处理
采用SPSS16.0统计软件one-way ANOVA程序进行显著性分析,采用Duncan氏法进行处理间多重比较,P<0.05为差异显著性标准,结果以“平均值±标准差”表示。
2 结果与分析
2.1 发酵条件优化
2.1.1 发酵时间优化
发酵时间对发酵液酶活性影响结果(图1)显示:随发酵时间延长,酶活性先升后降,发酵时间为42 h时,酶活性最高,为36.43 U/mL,与其他发酵时间结果相比,均存在显著差异。发酵时间从18 h到42 h,发酵液产酶量逐渐增加,不同发酵时间处理间酶活性均存在显著差异。
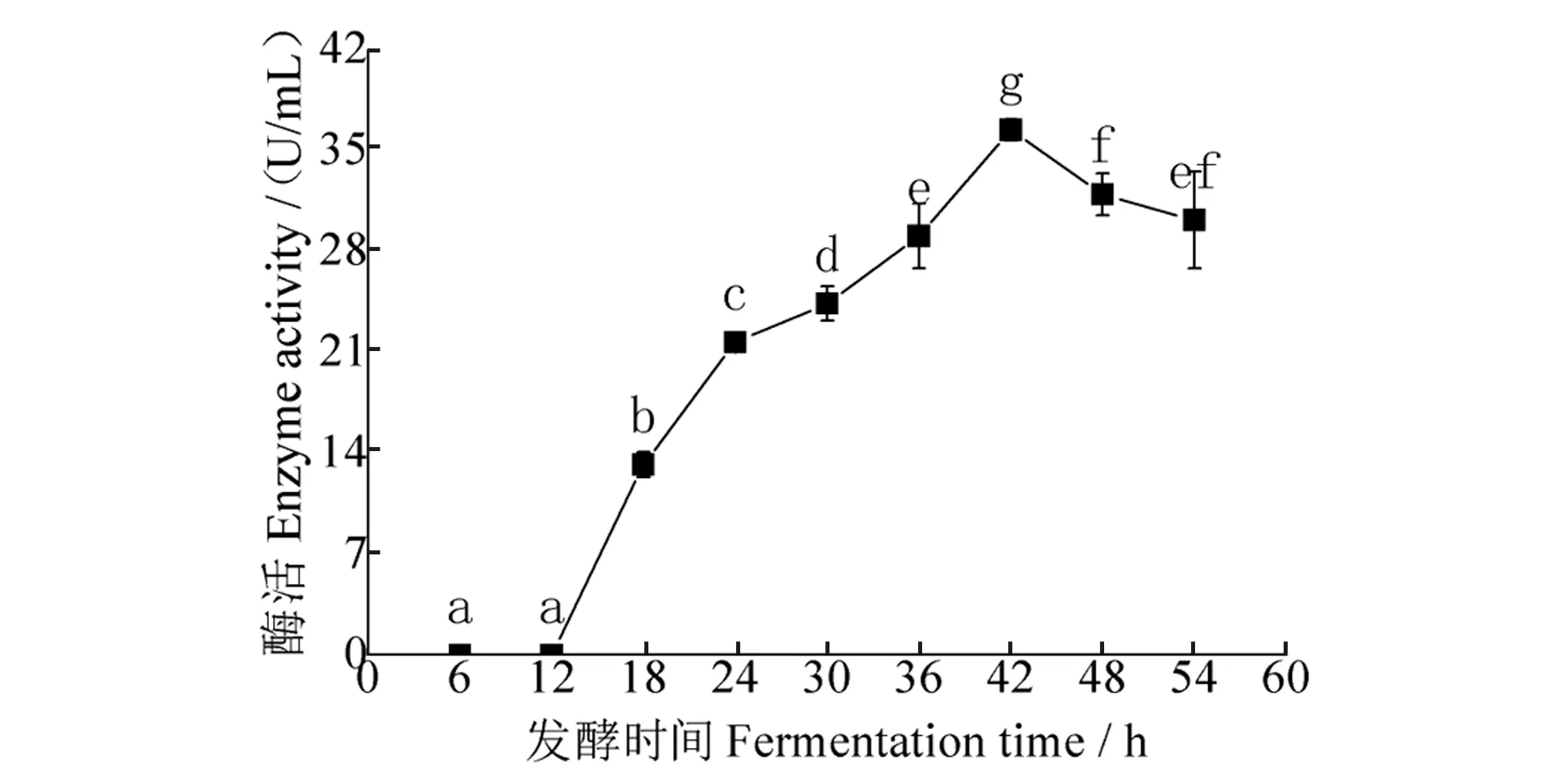
图1 发酵时间对酶活性的影响Figure 1 Effect of fermentation time on enzyme activity
2.1.2 培养转速优化
转速对发酵液酶活性影响结果(图2)显示:随转速增加,酶活性不断增加,转速为240 r/min时,酶活性最高,为40.90 U/mL,与其他处理酶活性相比,均存在显著差异。
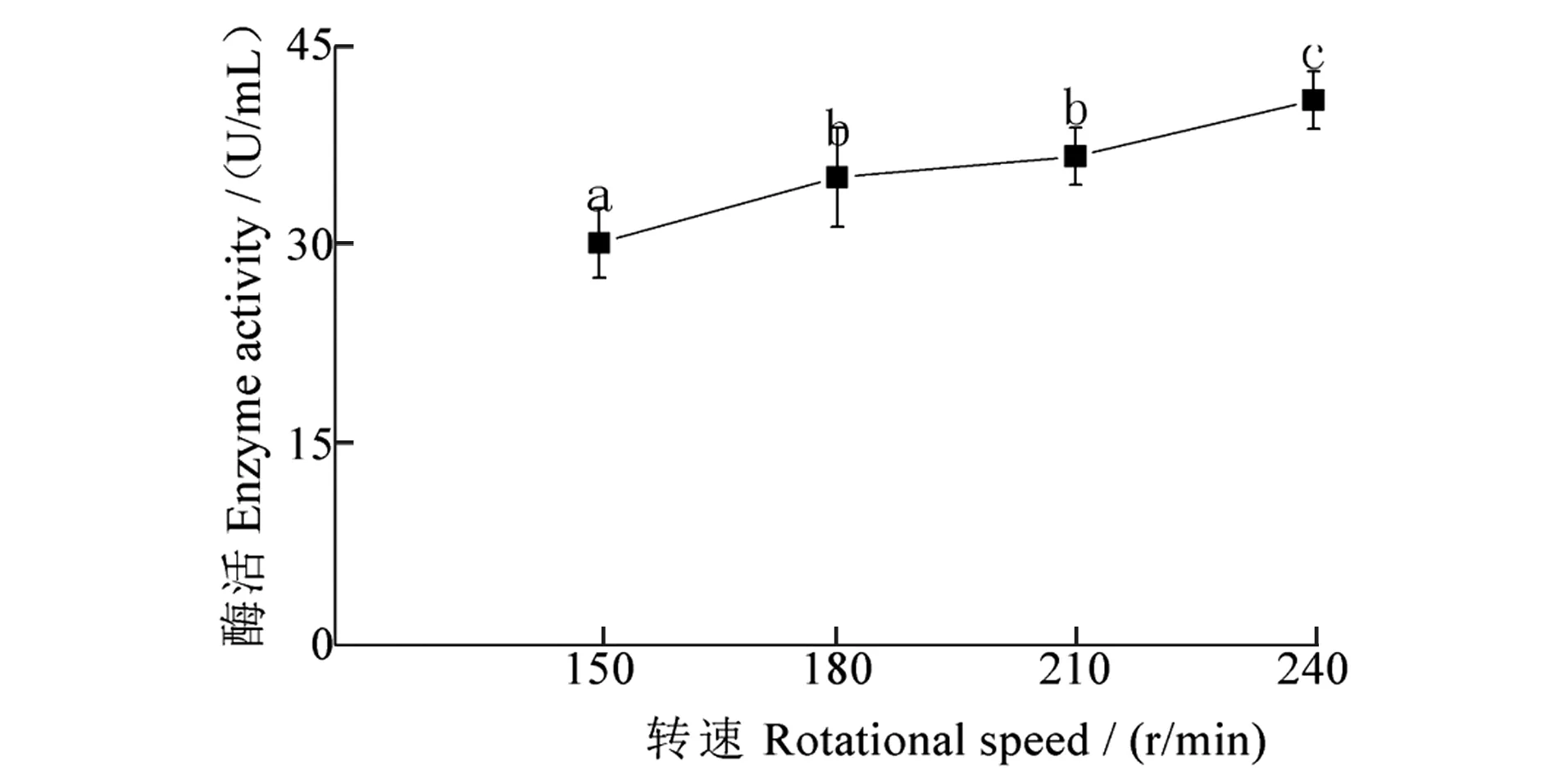
图2 转速对酶活性的影响Figure 2 Effect of rotational speed on enzyme activity
2.1.3 装液量优化
装液量对发酵液酶活性影响结果(图3)显示:随装液量增加,酶活性不断减小,装液量为10%时,酶活性最高,为47.59 U/mL,装液量为30%时,酶活性最低,为42.24 U/mL,10%装液量酶活性与20%、25%和30%装液量酶活性相比,均存在显著差异。
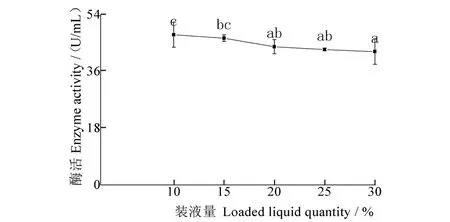
图3 装液量对酶活性的影响Figure 3 Effect of liquid culture volume on enzyme activity
2.1.4 培养基初始pH优化
初始pH对发酵液酶活性影响结果(图4)显示:随初始pH值升高,酶活性相对稳定而后急降,pH值为7.0时,酶活性最高,为48.71 U/mL,pH值为8.0时,酶活性显著下降,为12.10 U/mL。
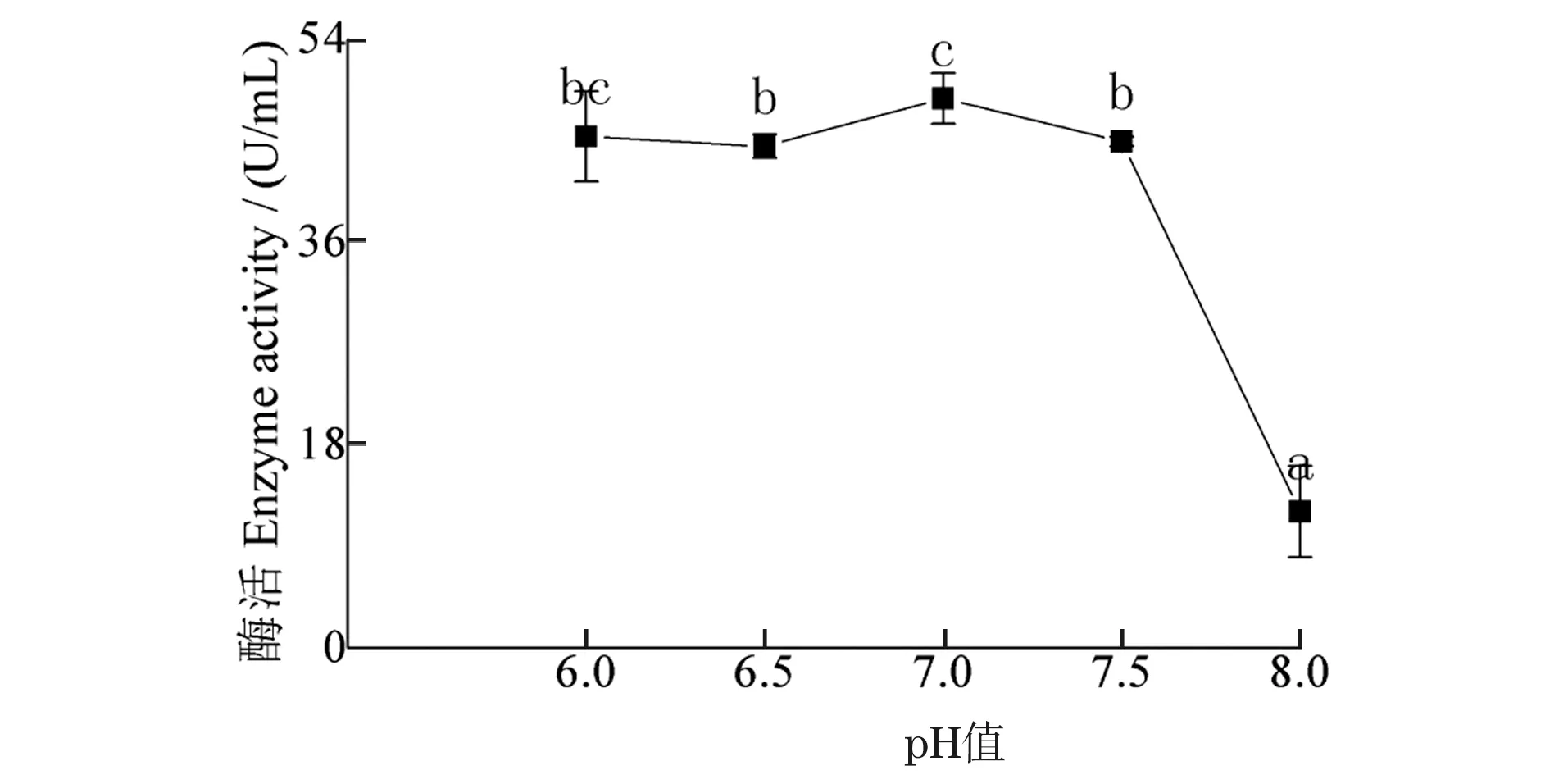
图4 初始pH值对酶活性的影响Figure 4 Effect of initial pH on enzyme activity
2.1.5 发酵温度优化
温度对发酵液酶活性影响结果(图5)显示:随温度升高,酶活性先增后减,温度为35 ℃时,酶活性最高,为79.07 U/mL,与33 ℃处理酶活性相比无显著差异,与其他处理相比均存在显著差异;温度为39 ℃时,酶活性为6.52 U/mL,与其他各处理相比均显著下降。

图5 温度对酶活性的影响Figure 5 Effect of temperature on enzyme activity
2.1.6 接种量优化
接种量对发酵液酶活性影响结果(图6)显示:随接种量增加,酶活性先升后降,接种量为4%时,酶活性最高,为79.51 U/mL,接种量为3%和5%时,酶活性较高,分别为75.72 U/mL和77.06 U/mL,且3%~5%接种量之间无显著差异。4%接种量处理与1%和2%接种量处理相比,酶活性显著增加;3%和5%接种量处理,与1%接种量处理相比,酶活性显著增加。
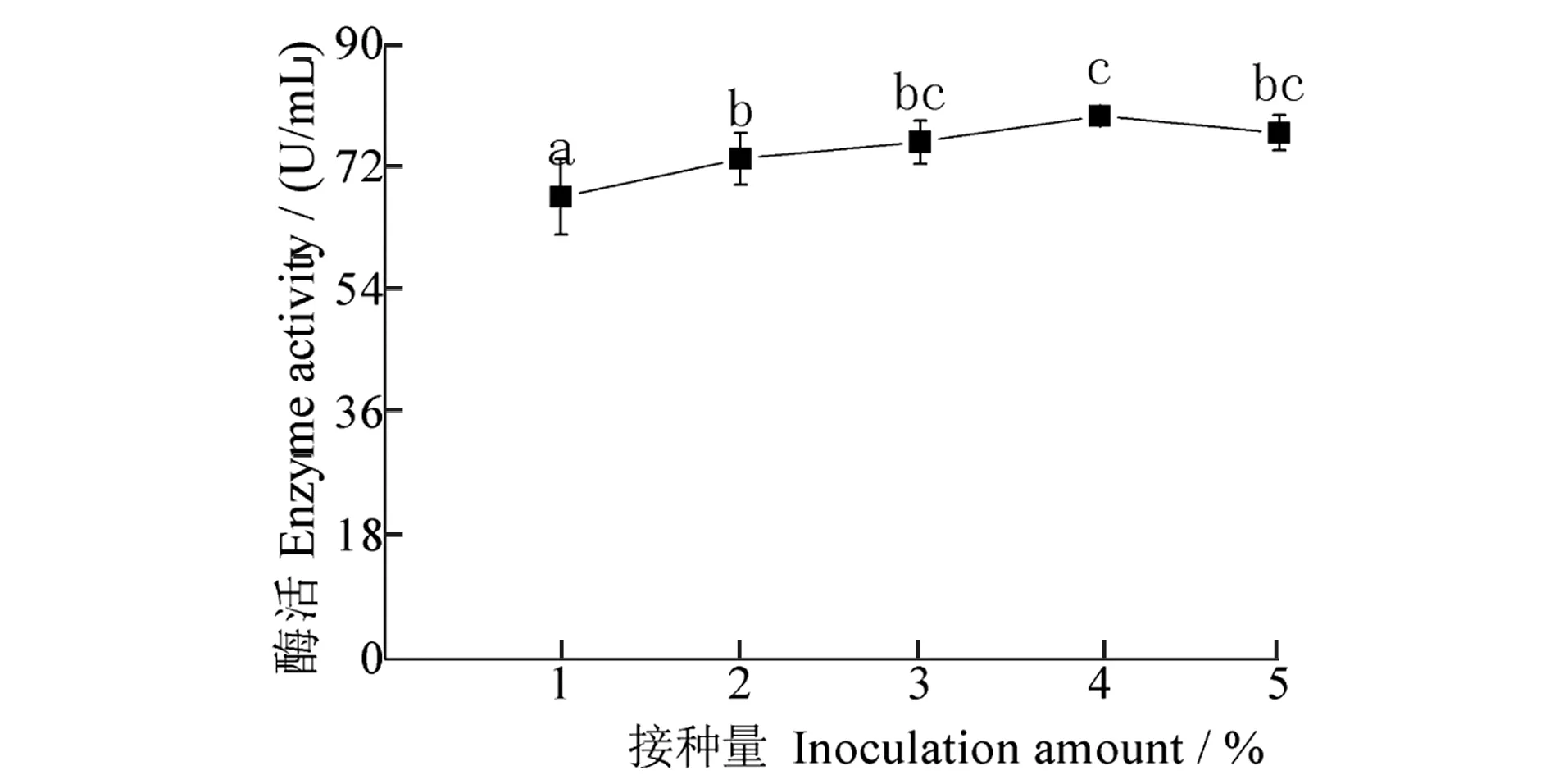
图6 接种量对酶活性的影响Figure 6 Effect of inoculation amount on enzyme activity
2.2 培养基成分优化
2.2.1 碳源种类优化
碳源对发酵液酶活性影响结果(图7)表明:蔗糖为碳源时,酶活性最高,为142.14 U/mL;淀粉为碳源时,酶活性较高,为105.71 U/mL;而红糖、糖蜜为碳源时,酶活性差。各种碳源处理酶活性间均存在显著差异,表明碳源对发酵产酶具有重要影响作用,选择最佳碳源种类至关重要。

图7 碳源对酶活性的影响Figure 7 Effect of carbon source on enzyme activity
2.2.2 氮源种类优化
氮源对发酵液酶活性影响结果(图8)表明:酵母膏为氮源时,酶活性最高,为229.64 U/mL,与其他氮源处理相比,酶活性均显著增加;蛋白胨、黄豆粉为碳源时,酶活性较高,分别为142.28 U/mL和87.61 U/mL,与其他氮源处理酶活性相比,均存在显著差异,二者间亦存在显著差异。其他氮源酶活性较差,且无显著差异。
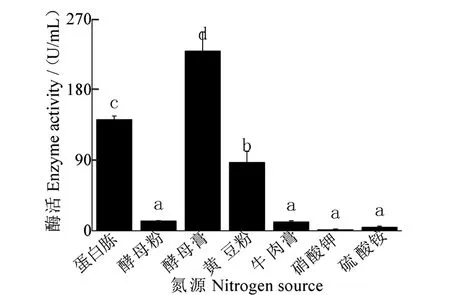
图8 氮源对酶活性的影响Figure 8 Effect of nitrogen source on enzyme activity
2.2.3 磷源种类优化
磷源对发酵液酶活性影响见图9:1/2磷酸二氢钾+1/2磷酸氢二钾为磷源时,酶活性最高,为189.43 U/mL;磷酸氢二钾为磷源时,酶活性最低,为168.09 U/mL,与其他两种磷源相比,存在显著差异。
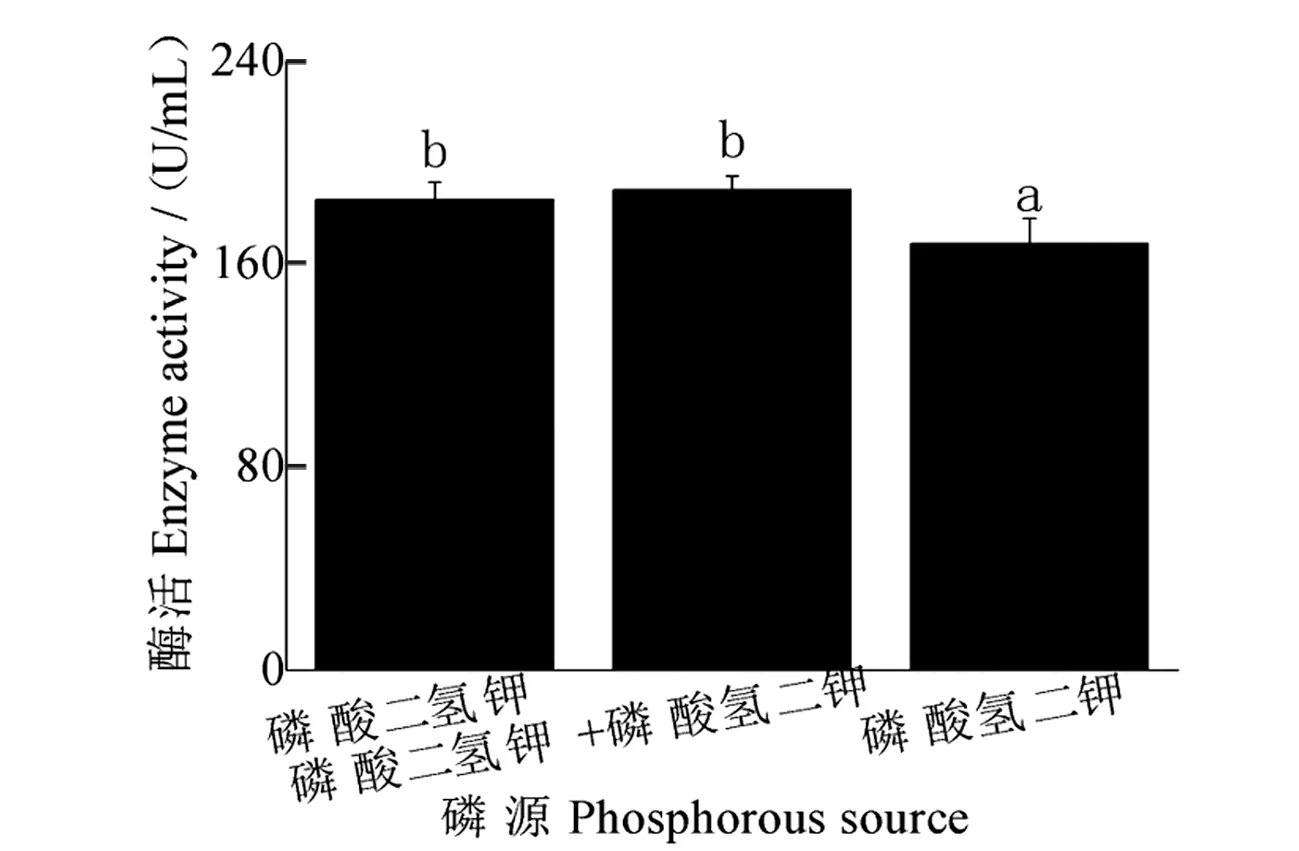
图9 磷源对酶活性的影响Figure 9 Effect of phosphate source on enzyme activity
2.2.4 无机盐种类优化
无机盐对发酵液酶活性影响结果(图10)表明:氯化钙、硝酸钾、硫酸锰、三氯化铁、硫酸铜和硫酸亚铁为无机盐时,酶活性较高,分别为368.36、359.65、332.24、332.24、327.56和326.17 U/mL,均优于CK,且存在显著差异。其他无机盐种类处理酶活性低于CK。
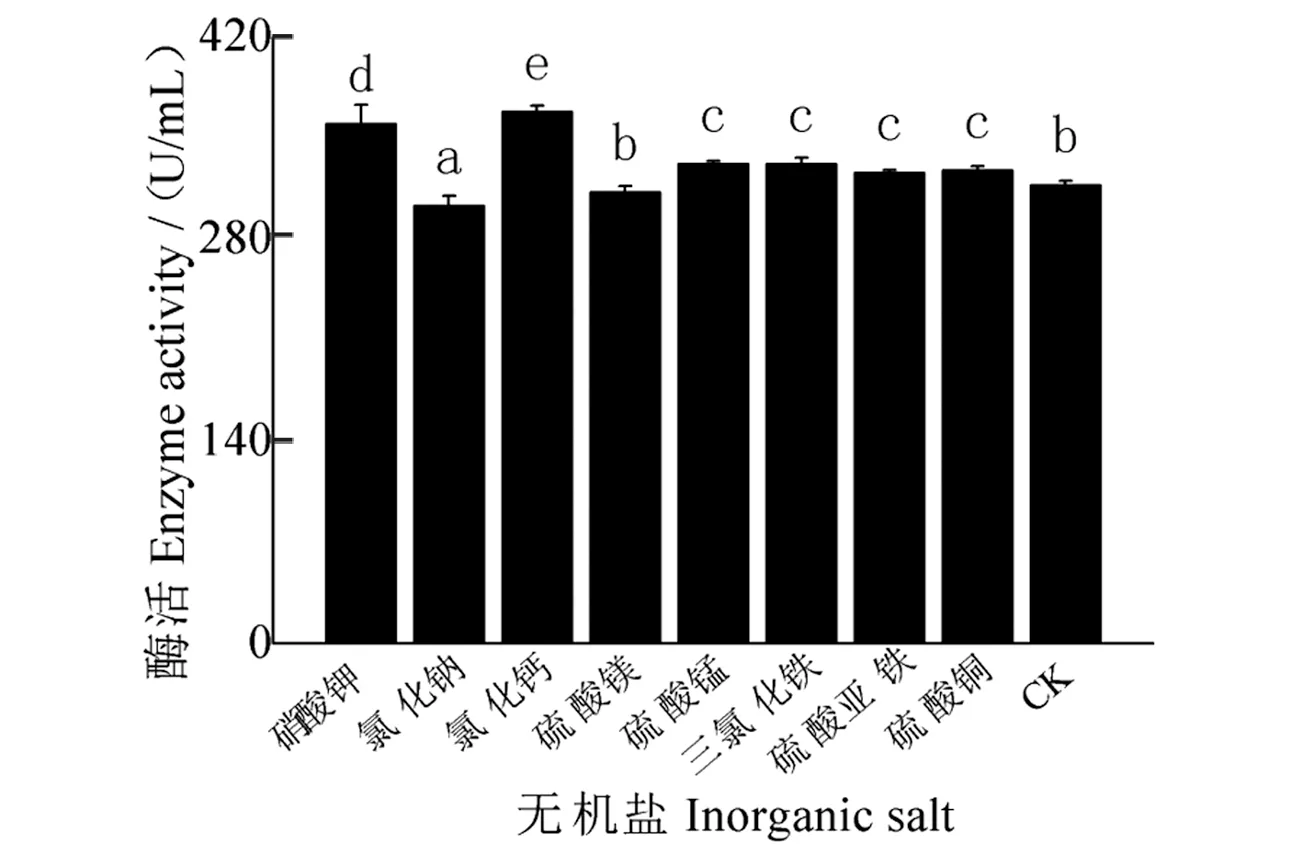
图10 无机盐对酶活性的影响Figure 10 Effect of inorganic salts on enzyme activity
2.2.5 表面活性剂种类优化
表面活性剂对发酵液酶活性影响结果(图11)表明:添加Tween-20、Tween-80、TritonX-100和PEG-400时,酶活性分别为407.66、404.07、403.56和389.06 U/mL,与CK相比均显著增加;而添加SDS时,酶活性有所下降,与CK相比无显著差异。
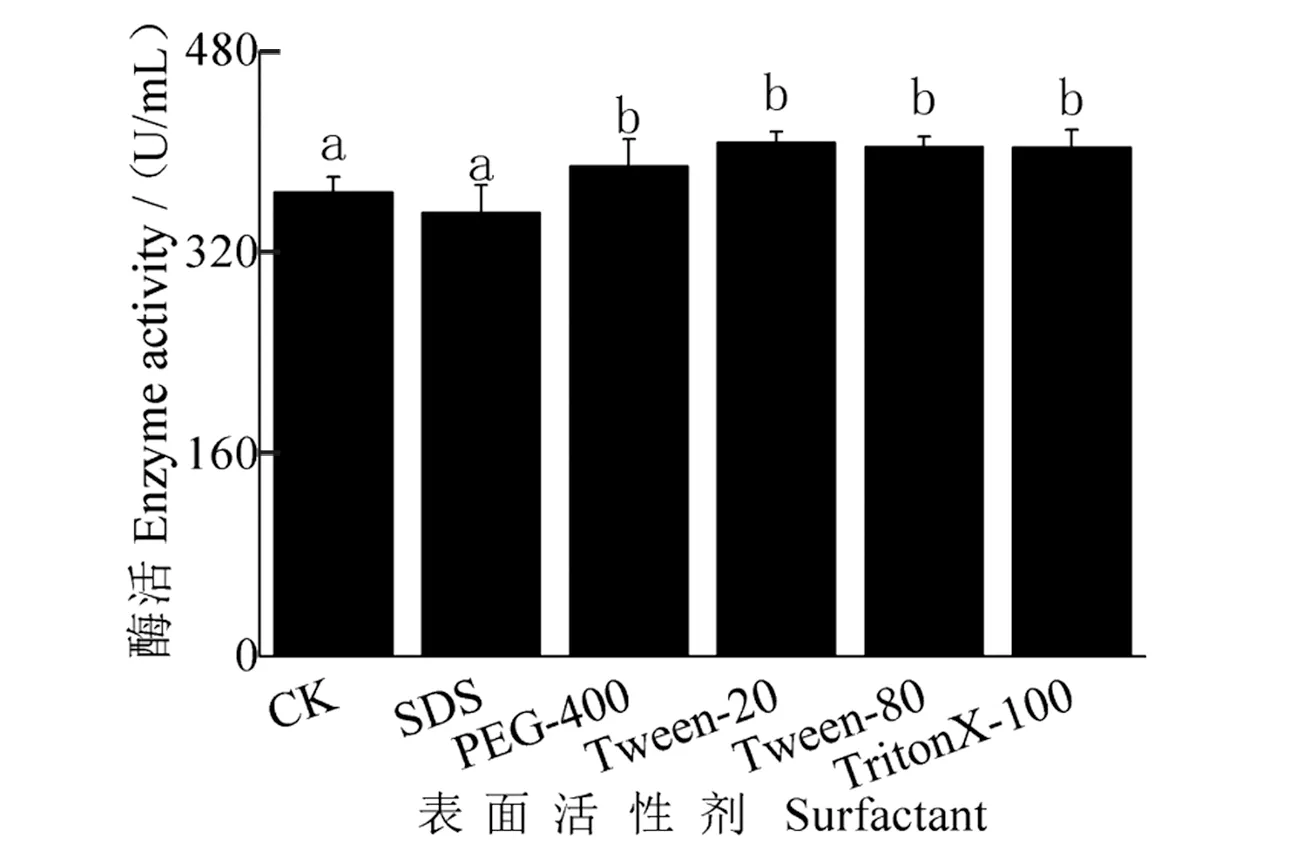
图11 表面活性剂对酶活性的影响Figure 11 Effect of surfactant on enzyme activity
2.2.6 生长因子种类优化
生长因子对发酵液酶活性影响结果(图12)表明:添加玉米浆、糖蜜和麦芽汁时,酶活性分别为448.65、412.32和432.54 U/mL,与CK相比,添加玉米浆和麦芽汁处理酶活性均显著增加,而添加糖蜜处理无显著差异。

图12 生长因子对酶活性的影响Figure 12 Effect of growth factor on enzyme activity
2.2.7 诱导物浓度优化
诱导物浓度对发酵液酶活性影响(图13)表明:诱导物浓度为0时,未检测到酶活性;随添加浓度升高,酶活性先升后稍降,添加浓度为1.5 g/L时,酶活性最高,为488.65 U/mL,与诱导物添加浓度为2.0 g/L处理相比无显著差异,与其他诱导物浓度处理相比均显著增加。

图13 诱导物浓度对酶活性的影响Figure 13 Effect of induces concentration on enzyme activity
2.3 培养基组成优化
正交试验结果见表2。极差分析表明:影响发酵液酶活性主次因素关系:A(蔗糖)>B(酵母膏)>G(玉米浆)>D(氯化钙)>C(1/2磷酸氢二钾和1/2磷酸二氢钾)>E(硝酸钾)>F(Tween-20);因素最佳水平组合:A2B3C2D3E2F3G1;最佳产酶培养基组成:蔗糖8.0 g/L,酵母膏11.0 g/L,磷酸二氢钾0.38 g/L,磷酸氢二钾0.38 g/L,氯化钙0.27 g/L,硝酸钾0.20 g/L,Tween-20 2.0 mL/L,玉米浆1.0 mL/L。最佳培养基组合不在正交试验组合内,3次重复验证试验结果表明:发酵液酶活性达(935.50±14.01)U/mL。方差分析结果(表3)显示:F蔗糖=48.723>F0.05(2,2)=9.28,蔗糖作用达显著水平,应严格控制其用量,其他因素未达显著水平。
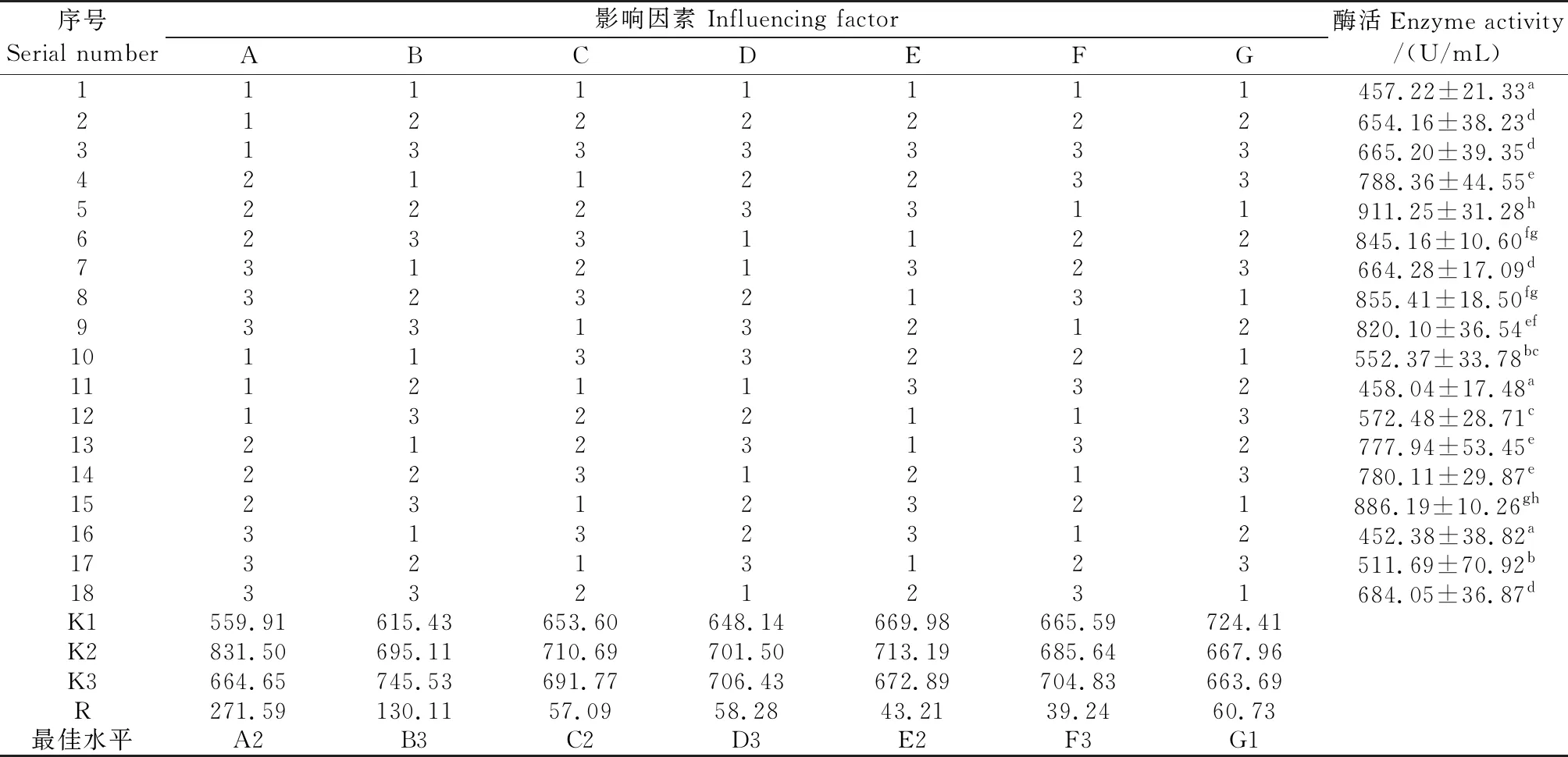
表2 正交试验设计及直观分析结果Table 2 Arrangements of orthogonal experiments and results of visual analysis

表3 正交试验方差分析Table 3 Variance analysis of orthogonal test
2.4 酶催化黄芩苷转化初步试验
采用试验最优发酵产酶工艺,在未对酶催化转化体系优化条件下,发酵产酶催化转化72 h,黄芩苷转化率为31.20%±4.15%。下一步工作将对该酶催化黄芩苷转化最适温度、pH等酶学特性及料液比等转化体系影响因素进行深入研究,优化和提升黄芩苷转化率。
3 讨论
发酵时间优化试验中,发酵12 h之前发酵液未检测到β-D-葡萄糖醛酸苷酶酶活性,12 h之后酶活性逐渐增加,初步判断该酶合成类型为滞后合成型,推测存在分解代谢物阻遏作用,在解除分解代谢物阻遏方面,有待进一步深入研究。发酵12 h至42 h,发酵液酶活性显著增加,之后发酵液酶活性有所下降,原因可能与发酵液营养物质减少、菌体出现衰退衰亡、发酵液pH变化影响酶稳定性有关[7]。培养基初始pH值在6.0~7.5,发酵液产酶量较高且稳定;pH值为8.0时产酶量显著下降,表明碱性环境不利于JY24菌株产酶及酶活性发挥。培养基初始pH值与细胞生长繁殖、发酵产酶关系密切,对菌体细胞膜通透性有直接影响,进而影响细胞膜稳定性及产酶活力[8]。发酵温度35 ℃时,酶活性最高,进一步提高温度,酶活性则显著下降。通常较低温度可提高酶所对应mRNA稳定性,增加酶生物合成延续时间,提高产酶量;但温度太低易影响细胞生长繁殖,减缓代谢速度,降低产酶量和延长发酵周期[9]。
碳源优化试验中,不同碳源发酵液酶活性间均存在显著差异,蔗糖为碳源时,酶活性最高,表明蔗糖是影响发酵液产酶的关键因素。Ku等[5]对德氏乳杆菌Rh2产β-D-葡萄糖醛酸苷酶的66种培养基进行筛选,基础培养基添加4%半乳糖产酶最佳。碳源优化时一般应考虑:菌株营养偏好选择;碳源对酶合成的代谢调节功能(诱导作用和分解代谢物阻遏作用)[10];原料来源是否充裕、价格是否低廉、有否影响发酵工艺等因素[11]。氮源优化试验发现有机氮源产酶效果显著优于无机氮源,这与异养型微生物细胞生长、繁殖和产酶一般要求有机氮源一致[12],可能无机氮源易使培养基pH降低致酶稳定性受影响有关。Chouiter等[1]报道树舌灵芝产β-D-葡萄糖醛酸苷酶最佳碳、氮源分别是阿拉伯胶和酵母膏,最佳氮源酵母膏与试验结果一致,最佳碳源则不同,可能与产酶菌株差异有关。
无机盐优化试验中,与对照相比,分别添加0.002 mol/L氯化钙、硝酸钾、硫酸锰、三氯化铁、硫酸铜和硫酸亚铁对发酵液酶活性均有显著促进作用。无机盐对细胞产酶有以下作用:构成细胞主要组成元素,构成酶分子组成元素,作为激活剂调节酶活性,起稳定氧化还原电位和渗透压作用。杨光等[13]研究表明Ca2+在0.5~10 mmol/L浓度范围内对β-葡萄糖醛酸苷酶具有激活作用;司磊[14]研究表明Ca2+、Mg2+和Cu2+对酶促反应具有促进作用,而Fe2+有抑制作用,Zn2+低与高浓度作用不一。综上,Ca2+作用比较一致,对β-葡萄糖醛酸苷酶数量和活性表现促进作用,其它金属离子作用存在出入。表面活性剂优化试验表明Tween 20产酶效果最佳。表面活性剂可增加细胞膜通透性,利于胞外酶分泌,提高产酶量。杨耀刚等[15]]研究报道Tween能够增加底物与酶有效结合,提高纤维素酶热稳定性,且Tween 20强于Tween 80;β-D-葡萄糖醛酸苷酶主要来源于细菌和真菌,通常分2种:胞内酶和胞外酶。JY24菌株产β-D-葡萄糖醛酸苷酶酶活检测前期试验中,发现发酵液上清酶活性较高,而菌体破碎后粗酶液几乎检测不到酶活力,判断所产该酶为胞外酶,进而在培养基成分优化中,选择添加表面活性剂进行研究,以期增加细胞膜通透性和产酶量。目前,优化微生物菌株产β-D-葡萄糖醛酸苷酶的研究报道较少,已有树舌灵芝、毕赤酵母、产紫青霉和球毛壳菌产β-D-葡萄糖醛酸苷酶的工艺优化报道[3-4],该酶活性范围1.09~2 356 U/mL,发酵产酶时间4~14 d,其结果与产酶菌株、酶促反应底物及菌株重组与否等差异有关。综上可知,试验菌株发酵1.75 d(42 h),酶活性达935.50 U/mL,较优化前提高25.68倍,与上述研究相比,酶活性处于中等水平,发酵产酶时间缩短2.25~12.25 d。
4 结论
鸡源饲用枯草芽孢杆菌JY24菌株产β-D-葡萄糖醛酸苷酶最佳发酵工艺:产酶条件:发酵时间42 h,转速240 r/min,装液量10%,培养基初始pH 7.0,发酵温度35 ℃,接种量4%;产酶培养基组成:蔗糖8.0 g/L,酵母膏11.0 g/L,磷酸二氢钾0.38 g/L,磷酸氢二钾0.38 g/L,氯化钙0.27 g/L,硝酸钾0.20 g/L,Tween-20 2.0 mL/L,玉米浆1.0 mL/L,黄芩苷1.5 g/L。在最佳发酵工艺条件下,鸡源饲用枯草芽孢杆菌JY24菌株产β-D-葡萄糖醛酸苷酶酶活性达935.50 U/mL,较优化前提高25.68倍,发酵产酶时间缩短2.25~12.25 d,酶催化初步研究表明黄芩苷转化率为31.20%。
