棉花MYB转录因子的研究进展*
2021-10-25朱英洁
朱英洁, 赵 航, 包 颖
(曲阜师范大学生命科学学院,273165,山东省曲阜市)
0 引 言
棉属(GossypiumL.),起源于热带和亚热带地区,类型包括灌木到小乔木,共约52种[1].根据染色体数目、核型以及鉴定的时间顺序,在棉属内已经发现并命名8种二倍体基因组(AA至GG以及KK)和1种四倍体基因组(AADD).目前,棉属中共有4个棉种被驯化,它们分别是2个具有AA基因组的二倍体——亚洲棉(G.arboretumL.)和草棉(G.herbaceumL.)以及2个具有AADD基因组的四倍体——陆地棉(G.hirsutumL.)和海岛棉(G.barbadenseL.)[2].栽培棉花作为世界重要的纤维作物,是纺织工业的重要原料支撑,在推动国民经济发展中占有重要地位.因此,进一步提高棉花的纤维品质和产量是促进棉花产业提升的关键.目前,分子育种作为作物改良的主要方法已经得到广泛应用,而要做好分子育种,除了需要识别形成植物农业表型的主效基因之外,筛查调控这些基因表达的转录因子同样重要.
转录因子是指能够结合在基因上游特异核苷酸序列上的蛋白质,这些蛋白质具有DNA结合结构域和转录激活结构域.MYB作为植物和动物中广泛存在的一类转录因子,其特点是在N端存在高度保守的DNA结合结构域(MYB DNA-Binding),该结构域通常包含1~4个半保守基序(R),每个R由大约50~53个氨基酸残基和间隔序列组成.这些氨基酸残基使MYB结构域折叠成一个螺旋-转角-螺旋(HTH)结构[3],可插入到目标DNA的大沟中.
根据结构域中R数目的不同,MYB转录因子可以分为4个亚家族:1R-MYB、R2R3-MYB、3R-MYB和4R-MYB.1R-MYB转录因子只含有一个MYB结构域,是一种重要的端粒结合蛋白,在调节基因转录和维持染色体结构的完整性中起重要作用[4,5].R2R3-MYB亚家族含有2个MYB保守结构域(R2和R3),是植物MYB家族中数目最多、功能最广泛的转录因子,是研究的重点.含有R1、R2、R3共3个MYB结构域的是3R-MYB亚家族,主要在细胞周期调节和细胞分化中起作用[6].4R-MYB转录因子含有4个MYB结构域,这类MYB转录因子数目最少且研究较少,很多功能未知.
本文通过回顾不同棉种MYB蛋白家族的进化特点以及生物学功能,对MYB转录因子在棉花中的研究做出阶段性的总结,为后续研究MYB转录因子在棉花中的功能、调控机制以及进化动态提供参考.
1 棉花MYB转录因子的进化特点
随着全基因组测序的兴起,棉属中部分棉种的全基因组已经公布,目前基于二倍体亚洲棉、草棉和雷蒙德氏棉(G.raimondiiL.,DD基因组),四倍体陆地棉和海岛棉全基因组的MYB转录因子鉴定已经展开[7-9](表1).在这5个基因组中,四倍体陆地棉中鉴定的MYB转录因子数量最多,共有812个;二倍体草棉和雷蒙德氏棉分别鉴定到数目几乎相等的MYB(349,348)(表1).MYB的4个亚家族在棉花中都有分布,其中R2R3-MYB亚家族数量最多.陆地棉、草棉和雷蒙德氏棉基因组内分别有55%、59%和62%的MYB转录因子隶属R2R3-MYB亚家族(表1);与之相反,来自4R亚家族的MYB数量最少,目前仅在二倍体棉中鉴定到1个直系位点,且该位点在四倍体棉中以倍增的形式存在(表1).
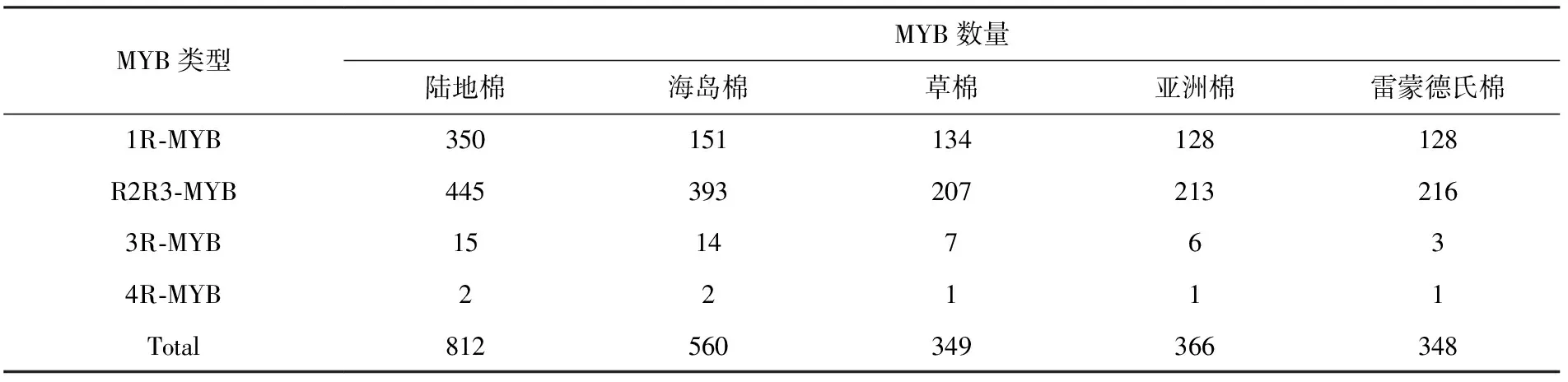
表1 棉花中全基因组鉴定的MYB转录因子
棉花MYB转录因子根据进化时间不同可分为3类:在祖先棉花分化之前就已存在的MYB转录因子,广泛存在于各种植物中;在AA基因组和DD基因组二倍体棉花分化后,由MYB转录因子经过不断复制产生,属于棉花特有的MYB转录因子;进入AADD基因组,二倍体亲本MYB在全基因组加倍后,经过丢失、重排、以及再扩增等出现的MYB.系统进化研究发现,四倍体棉花中D基因组可能的祖先二倍体和雷蒙德氏棉最接近;而A基因组的供体则被认为是已经灭绝的A0[10].理论上,如果基因没有发生丢失,四倍体内的MYB总数应该等于2个二倍体内MYB之和,但是,目前四倍体陆地棉中MYB数量(812)却远大于AA和DD基因组二倍体棉的MYB转录因子总数之和(697)(表1),表明MYB在四倍体棉花内可能存在大幅度的扩增现象,或者是在二倍体中有基因丢失(图1).由此可以看出,四倍体棉中MYB蛋白家族的进化除了由全基因组加倍导致的直系同源倍增基因之外,还有另外的复制方式存在.最近研究证实了这种推测,串联复制和转座复制在棉花MYB家族扩增中发挥重要作用[11].

图1 MYB在3种棉花不同染色体上的共线性分布
研究证明,大部分MYB同源基因在二倍体之间不但以相同的速率进化,而且基因时空表达的模式在棉种间也相对保守[8].但是,当这些同源基因被异源整合到四倍体棉中,倍增基因组中冗余重复的MYB基因则可能经历不同的命运.近期有研究发现陆地棉中来自D基因组的MYB转录因子显示出比来自A基因组的同源拷贝具有更快的进化速率,并且两种同源拷贝在所有器官中也不再等量表达,表明这些同源拷贝间可能已经发生了功能上的亚分化[7].
2 棉花中MYB转录因子的生物学功能
棉花作为重要的经济作物,因此如何提高其产量和纤维质量受到广泛关注.通过汇总棉花MYB转录因子研究的相关文献,我们发现已有功能报道的MYB转录因子共有56个,其中45个MYB转录因子属于陆地棉,海岛棉和亚洲棉报道的分别有6个和5个.根据已有报道,可以看出MYB转录因子广泛参与棉花生长发育阶段的多种生物学过程,例如参与纤维发育、花青素生物合成、应对生物和非生物胁迫等(表2).
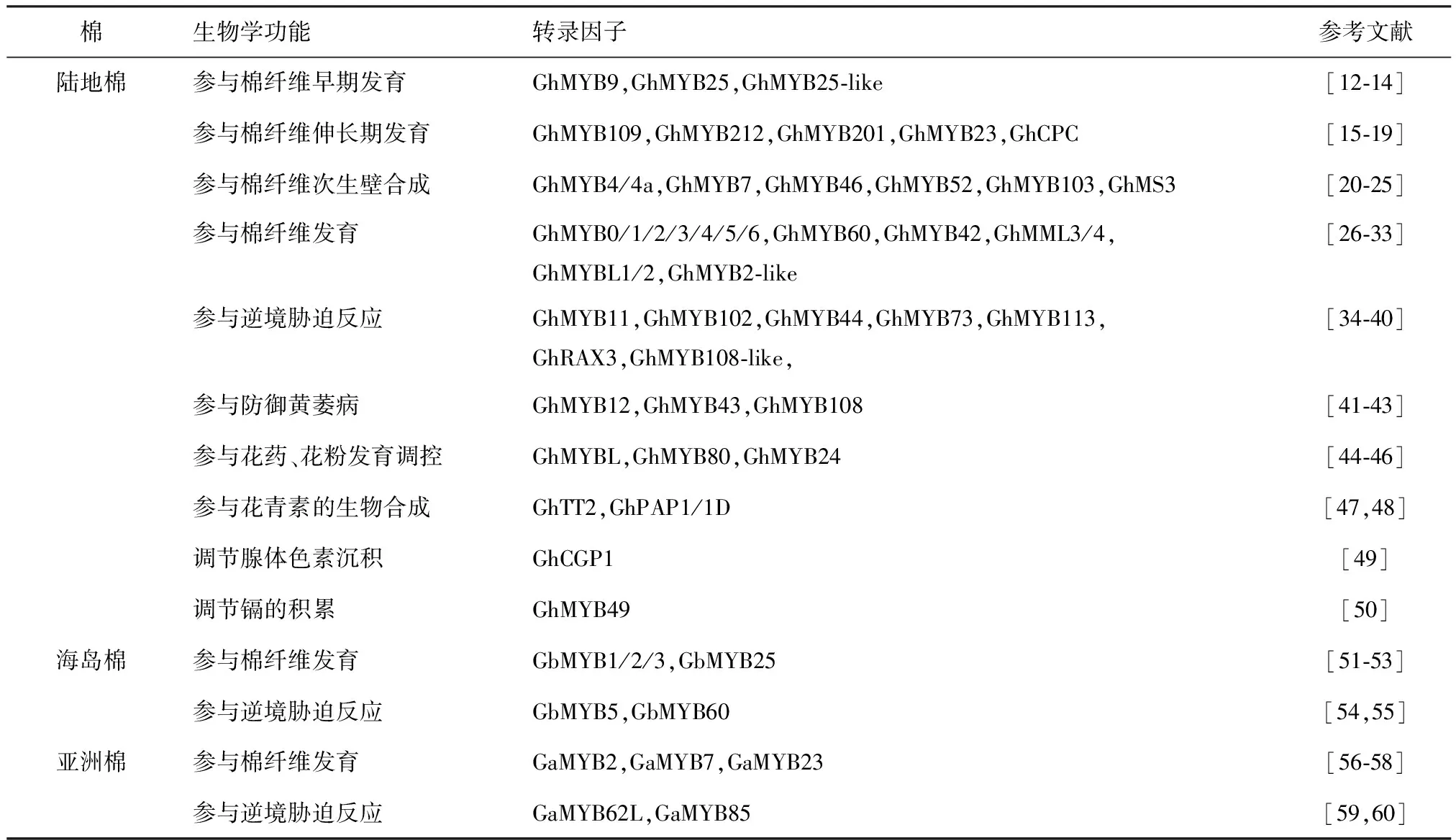
表2 棉花中已报道的MYB转录因子的功能
2.1 MYB转录因子在纤维发育中的作用
棉纤维是棉种皮表皮层的单细胞毛状体.作为世界上重要的天然纤维材料,棉纤维的发育可分为4个阶段:起始阶段(Initiation)、伸长阶段(Elongation)、次生细胞壁合成阶段(Secondary cell wall synthesis)和成熟阶段(Maturation).棉纤维在4个不同发育阶段发生的变化均会对棉花的品质和产量产生影响.MYB作为植物重要的转录因子家族之一,已被证明在棉纤维发育的各个时期均有不同的调控作用(图 2).在棉纤维发育起始阶段发挥作用的MYB基因主要有GhCPC、GhMYB25和GhMYB25-like.拟南芥CAPRICE(CPC)基因被证实是毛状体起始的负调节剂.在棉花中发现GhCPC与GhTTG1/4和GhMYC1组成CPC-MYC1-TTG1/4复合物,进而调节GhHOX3(GL2)和GhRDL1 的表达来负调节棉纤维的起始和早期伸长[15].
目前已鉴定在棉纤维伸长期起调控作用的MYB转录因子数量最多(图2),其中GhMYB109可抑制乙烯合成酶(GhACO1和GhACO2)的表达,且在棉花中沉默GhMYB109会导致转基因棉花纤维长度变短,表明其有可能参与纤维细胞伸长的调控[61].有研究者分别从亚洲棉和海岛棉中克隆出了MYB2基因,在过表达拟南芥中GbMYB2可激活拟南芥发育相关基因(GL2)的表达,促进叶片毛状体和根的发育;GaMYB2和GbMYB2功能相似,在植物表皮毛细胞分化和发育过程中起重要的调控作用[51,57].这些研究表明在不同植物中MYB转录因子的功能有一定的保守性.
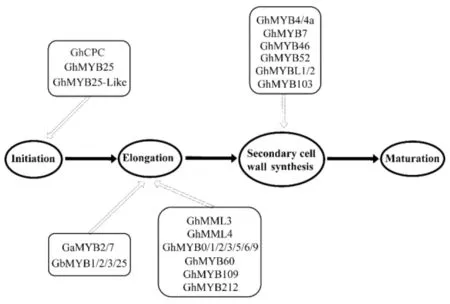
图2 棉纤维形成过程中已知的MYB转录因子
棉花纤维发育的第三阶段(次生壁合成阶段)决定着棉花纤维的强度和韧度.一些 R2R3-MYB转录因子在该阶段通过调控纤维素合酶(celA1)的转录来调节次生壁的纤维素合成.例如, GhMYBL1可识别并结合在celA1启动子的SMRE元件促进次生壁中纤维素的合成[62].同样,GhMYB46也是通过结合celA1的启动子,在棉纤维的次生壁合成阶段起作用[21].
2.2 MYB转录因子在胁迫中的作用
棉花在生长发育过程中受到各种生物和非生物胁迫,如干旱、低温、高盐以及病虫害侵染等.在长期的进化过程中,棉花形成了抵抗胁迫的多种防御机制.有研究表明,MYB转录因子可通过直接或间接调控多个抗逆相关基因的表达,以应对外界不良环境.
2.2.1非生物胁迫
极端环境(干旱、高盐和低温等)会对植物的生长和发育产生许多不利的影响,严重危害作物的品质和产量.植物激素脱落酸(ABA)是植物体内一种重要的生物活性物质,参与植物发育和非生物胁迫反应等过程[63].棉花对ABA信号的感知和转导有ABA依赖和非ABA依赖性两条途径,其中已报道的MYB转录因子是通过ABA依赖途径参与非生物胁迫.
在棉花中,干旱和盐胁迫会使脱落酸(ABA)含量上升,进而引起棉花的适应性反应.当ABA含量较低时,2C型蛋白磷酸酶(PP2C)通过去磷酸化与SNF1相关蛋白激酶2(SnRK2)相互作用并使其失活;当植物遭受干旱、盐等胁迫刺激时,ABA含量增加,并与ABA受体蛋白PYL相互作用并激活PP2C,形成PYL-ABA-PP2C三元复合体,释放SnRK2s[64].释放的SnRK2s通过自身磷酸化激活,并磷酸化许多下游的效应蛋白从而调节胁迫诱导基因的表达,气孔的开闭和离子转运等[63].在棉花中,MYB转录因子主要通过两条途径调控ABA介导的胁迫响应:一类是可与ABA受体蛋白PYL相互作用,影响后续途径;另一类是通过影响下游胁迫基因的表达来应对胁迫.研究发现GhMYB44可与ABA受体蛋白PYL相互作用以结合PP2Cs,通过影响ABA途径增强植株的抗旱能力[34].Chen等[54]研究显示在干旱胁迫下,GbMYB5通过激活ABA依赖性信号传导途径中的脱水反应基因的表达来减少植物失水,并通过调节渗透压和LEA蛋白质的生物合成来维持植物细胞和蛋白质的稳定.不仅如此,GbMYB5还通过有效的排毒系统清除活性氧(ROS).陆地棉中GhMYB11、GhMYB108-like和GhMYB113,在多种非生物胁迫处理后,表达量明显变化,也被认为在棉花抵御非生物胁迫的过程中具有一定作用[35,38,39],但其生物学功能及分子机制有待于更进一步研究.
2.2.2 生物胁迫
自然条件下棉花易受到黄萎病菌和棉铃虫的侵害.越来越多的研究表明,植物MYB转录因子参与抗病原体攻击的防御,在棉花中MYB转录因子抵御黄萎病菌攻击主要集中在防御信号分子途径.Cheng等[41]发现过表达GhMYB108可以增强拟南芥对黄萎病的耐受性,GhMYB108与钙调蛋白GhCML11相互作用形成正反馈环,并以Ca2+依赖性方式增强GhMYB108和GhCML11转录.GhCML11和GhMYB108沉默的植物中可减少Ca2+流入棉花根细胞的胞质溶胶中并下调钙信号相关基因的表达,这也印证了Ca2+/CaM和TFs协同作用调节防御相关反应.最近Shen等[43]克隆得到GhMYB43,发现超表达GhMYB43的棉花可增强棉花对黄萎病菌的敏感性,而且GhMYB43负调控木质素合成和JA信号通路相关基因的转录.在应对棉铃虫侵害的研究中发现,MYB转录因子可通过调控类黄酮次生代谢产物——花青素来增强棉花抵御棉铃虫的能力.Li等[47,65,66]研究发现棉花R2R3-MYB转录因子家族中GhPAP(1A、1D、2A和2D)可结合GhUGFT和GhGST启动子并激活其转录,进而调控棉花花青素的积累.
3 展 望
MYB作为植物体内庞大的蛋白家族,因其功能的重要性以及调控网络的复杂性,受到国内外研究人员的广泛关注.近年来,MYB蛋白家族参与植物生长发育和胁迫响应方面的研究取得突破性进展.然而,目前该家族成员的研究主要集中在拟南芥等少数模式植物中,许多作物特别是棉花中MYB蛋白的功能和调控机制尚不清晰.究其原因主要是以下几点:首先,尽管棉花中已初步报道了部分MYB转录因子在纤维发育和胁迫响应方面的功能,然而这些大多是依赖异源转化或者瞬时表达的实验结果,如何利用这些MYB转录因子提高棉花抗逆性或者培育优质的棉花品种尚有待深入研究.其次,MYB转录因子的研究主要集中在陆地棉,棉属其他种的研究相对较少,且研究内容多集中于生物信息学数据的推测.因此,进一步阐明棉花MYB家族的生物学功能以及其参与的转录调控机制是非常必要的.
现代分子生物学手段在植物学领域的广泛应用以及组学技术的快速发展,极大推动了棉花MYB家族功能及调节机制的研究.今后我们可以从以下几方面深入探究棉花MYB蛋白家族生物学功能;第一,借助基因编辑及棉花转基因技术,获得MYB的过表达及突变体材料,在稳定表达的棉花株系中探究MYB家族的生物学功能及调控机制.第二,通过蛋白互作技术,筛选并鉴定棉花体内MYB家族的相互作用蛋白,进而完善棉花MYB蛋白的转录调控网络.第三,不同种类的MYB转录因子在棉花胁迫响应以及纤维发育过程中发挥重要作用,这两种性状直接关乎棉花的经济效益.后续需要通过对MYB转基因棉花性状分析,筛选抗逆性强且纤维发育正常的MYB蛋白成员用于棉花分子育种,继而为培育抗逆、高产、优质的棉花品种提供理论依据和种质资源.
