温度和光周期对蜀柏毒蛾滞育的影响
2021-10-14杨远亮曾全肖银波贾玉珍范成志
杨远亮,曾全,肖银波,贾玉珍,范成志
四川省林业科学研究院森林保护研究所,四川成都 610081
滞育是指昆虫生长、发育和繁殖暂时停滞的一种生命现象[1-2]。昆虫普遍存在滞育现象,在长期的演化过程中,昆虫为了适应周围环境的变化,滞育成为其渡过不良环境的主要对策。同时,滞育能够使昆虫的生长、发育与季节、寄主物候等保持一致,有利于昆虫的活动、取食,确保其种群数量,有利于其种族延续[3]。研究表明,昆虫滞育的主要诱导因素有温度、湿度、光周期、食物质量及种群密度等,滞育的解除除了需要一定的时间之外[4],采取一些特殊的方法如高温、低温、光照、化学刺激和物理刺激等也能有效的解除滞育[4-5]。滞育以及滞育的调节机理研究是昆虫学研究的一个重要领域[1],研究分析滞育因子对于探索有关昆虫滞育理论、昆虫饲养中有效地诱导和解除滞育以及害虫预测防控等具有重要意义。
蜀柏毒蛾(Parocneria orientaChao),属于鳞翅目(Lepidoptera)毒蛾科(Lymantriidae)柏毒蛾属(Parocneria),分布于我国四川、福建、湖北和浙江等省。蜀柏毒蛾是四川省柏木林区最重要的食叶害虫,周期性爆发危害,常造成严重的经济、生态损失,给长江中上游生态屏障建设成果之一的柏木防护林构成威胁,因此其一直是四川省主要的森林害虫防治对象。许多学者先后进行过其生物学[6]、分布规律与抽样技术[7-8]、种群动态[9]、防治时期和防治方法[10]等方面的研究。蜀柏毒蛾在四川地区1年发生2代,越冬代有明显的滞育现象,主要表现为越冬代卵期和幼虫期滞育[11],但针对蜀柏毒蛾滞育还未见更深入的研究报道。蜀柏毒蛾越冬代发生期是其防治的关键时期,研究蜀柏毒蛾的滞育机制对于虫害的预测预报以及防控有重大意义。同时,探明滞育关键因子,将为实现人为调控蜀柏毒蛾滞育,解决蜀柏毒蛾人工饲养中因滞育问题引起的传代难题,建立蜀柏毒蛾人工饲养种群起到关键性作用。因此,本文将从温度和光周期两个方面对蜀柏毒蛾幼虫的滞育影响进行研究,为该虫种群发生动态的预测防控以及人工种群繁育提供理论依据。
1 材料与方法
1.1 试验昆虫
在德阳市中江县、绵阳市盐亭县和南充市南部县等蜀柏毒蛾常发柏木林区,设立林间调查样地,采取林间定点不定期抽样调查的方式观察蜀柏毒蛾越冬代和第1代的生长发育进度。分别于2019年1月和5月以及2020年5月,蜀柏毒蛾越冬代和第1代的卵期、幼虫期(1龄),进行跟踪、采集,以获得试验用虫。
1.2 实验器材
人工气候箱(宁波江南仪器厂,RXZ型);温湿度记录仪(江苏省精创电气股份有限公司,型号RC-4HA);饲养盒(口径15 cm,底径7.5,高7 cm,纱网孔盒盖);饲养盒遮光罩;培养皿;定性滤纸;毛笔等。
1.3 幼虫滞育判断
通过前期饲养观测发现,蜀柏毒蛾越冬代滞育个体表现为:初孵幼虫静伏不动,不取食,亦不蜕皮,即滞育1龄幼虫。同时,前人研究表明,蜀柏毒蛾第1代非滞育个体卵期约15 d、1龄幼虫期约25 d,而其越冬代在自然条件下,卵滞育个体卵期约110 d、1龄幼虫期28 d;幼虫滞育个体卵期约24 d、1龄幼虫期约144 d[11]。本试验以蜀柏毒蛾1龄幼虫为实验对象,因此,在统计滞育个体数量时,以滞育个体表现结合1龄幼虫发育历期,来判断滞育幼虫个体,确认其数量。
1.4 温度和光周期对蜀柏毒蛾(第1代)滞育诱导的影响
试验运用人工气候箱进行,温度梯度设置为12℃、15℃、18℃、22℃、25℃和28℃,并组合光周期2L:22D(光:暗,下同)、4L:20D、12L:12D、14L10D、18L:6D。光周期的控制通过暗期处理采用不透光黑色卡纸遮光罩进行人工遮光,光期移除遮光罩来实现。光照强度为15 000 lx,湿度75%±5%,共30个处理。以蜀柏毒蛾的第1代初孵幼虫(1龄)作为实验材料,将初孵幼虫放入底部垫有保湿滤纸和新鲜柏木鳞叶的塑料养虫盒内,每盒约30头左右。每个处理重复3次。定期更换保湿滤纸与新鲜柏木鳞叶以保持养虫盒清洁干净。观察和记录滞育个体的数量,统计不同处理中的滞育率。
1.5 温度和光周期对蜀柏毒蛾(越冬代)滞育解除的影响
以蜀柏毒蛾越冬代滞育1龄幼虫作为实验材料,实验于人工气候箱内进行。试验温度梯度设置为18℃、22℃、25℃、28℃和30℃,并组合光周期10L:14D、12L:12D、14L:10D,湿度均为75%±5%,共15个处理。将滞育幼虫放入底部垫有保湿滤纸和新鲜柏木鳞叶的养虫盒内,置于各处理条件下进行滞育解除试验,幼虫开始活动、取食视为滞育解除,统计各处理的滞育解除率。
1.6 数据处理
实验所得数据均用SPSS20.0软件进行统计分析,结果用平均数(Mean)±标准误(SE)来表示。对蜀柏毒蛾幼虫在不同恒温温度和不同光周期条件下的滞育率,以及不同温度和光周期条件下的滞育解除率进行单因素方差分析,采用Ducan’s法进行差异显著性检验;对蜀柏毒蛾幼虫在不同温度和光照下的滞育率进行二因子随机设计方差分析,采用Duncan’s新复极差法进行多重比较检验。
2 结果与分析
2.1 温度对蜀柏毒蛾滞育诱导的影响
在相同光周期条件下,不同恒定温度处理的蜀柏毒蛾幼虫滞育率存在极显著差异(P<0.01),见表1。在各光周期处理下,蜀柏毒蛾幼虫滞育率均随着温度的降低而升高。在中性光周期(12L:12D)下,25℃和28℃处理下,无滞育个体,滞育率为0%;22℃处理,滞育率为15.31%;当温度降低到18℃时,滞育率上升为30.23%;在12℃和15℃条件下,滞育率高达80%以上.。短光照和长光照处理下,温度对滞育率的影响也表现出相同的规律。此结果表明,蜀柏毒蛾幼虫滞育率与温度呈负相关,低温会抑制蜀柏毒蛾生长,诱导其进入滞育。
2.2 光周期对蜀柏毒蛾滞育诱导的影响
光周期对蜀柏毒蛾幼虫滞育率的影响如表1所示,当温度在12℃、15℃、18℃、22℃时,光周期对蜀柏毒蛾幼虫的滞育率有显著作用(P<0.05),在这些温度条件下,随着暗期的增长,蜀柏毒蛾滞育率显著上升,由此可知蜀柏毒蛾属于长日照反应型昆虫。18℃时,在长暗期条件下,蜀柏毒蛾幼虫有超过50%的幼虫进入滞育,中性光照处理滞育率30%,长光照处理滞育率约25%。25℃时,在短光照条件下极少数蜀柏毒蛾幼虫表现出滞育状态(该处理下死亡率较高),中性和长光照光周期下滞育率均为0%;28℃时,蜀柏毒蛾幼虫在短光照、中性和长光照处理下都没有发生滞育。说明在环境温度较高时,短光照对蜀柏毒蛾无滞育诱导作用。
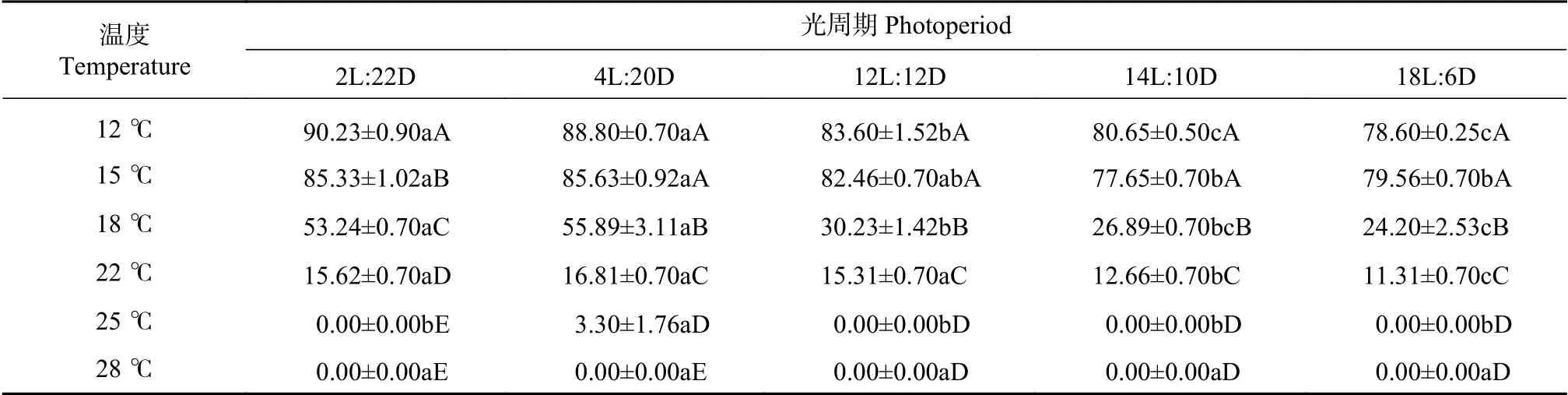
表1 温度和光周期对蜀柏毒蛾滞育的影响Tab.1 Effectsof temperature and photoperiod on diapauseof Parocneria orienta Chao
2.3 温度和光周期对蜀柏毒蛾滞育的交互作用
通过对温度和光周期诱导蜀柏毒蛾滞育的双因素方差分析可知(见表2),温度(F(4,50)=1476.370,P<0.01)和光周期(F(4,50)=140.536,P<0.01)对蜀柏毒蛾滞育率的诱导有着显著影响,温度与光周期对蜀柏毒蛾滞育诱导的交互作用显著(F(4,50)=6.780,P<0.01)。说明,在相同光周期条件下,蜀柏毒蛾的滞育率随着温度的降低而升高,温度越低滞育率的变化越明显。在温度和光周期共同作用的诱导效应中,光周期只在一定范围内的温度内起作用,说明温度是主要因子,起决定性作用,光周期是次要因子,起辅助作用。
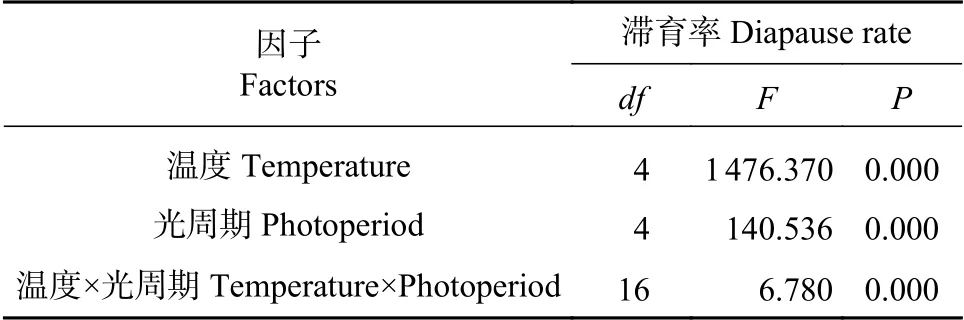
表2 光周期和温度对蜀柏毒蛾滞育率的方差分析Tab.2 Variance analysisof temperature and photoperiod on diapause rate of Parocneria orienta Chao
2.4 温度和光周期对蜀柏毒蛾滞育解除的影响
在光周期为10L:14D、12L:12D、14L:10D的长光照条件下,将滞育幼虫分别饲养在18℃、22℃、25℃、28℃和30℃的设置温度进行滞育解除试验。结果表明,除在30℃时,14L:10D光周期下的滞育解除率显著高于10L:14D和12L:12D处理外,其他相同温度不同光周期条件处理下,蜀柏毒蛾1龄幼虫的滞育解除率没有显著差异(P<0.05)。相同光周期不同温度条件下,蜀柏毒蛾幼虫的滞育解除率有极显著差异(P<0.01),18~25℃下,蜀柏毒蛾1龄幼虫的滞育解除率随温度上升而逐渐增加,温度上升至28℃后,滞育率开始下降(见表3)。由此可知,25℃条件下,最有利于蜀柏毒蛾幼虫滞育解除,光周期对蜀柏毒蛾滞育解除作用不显著。
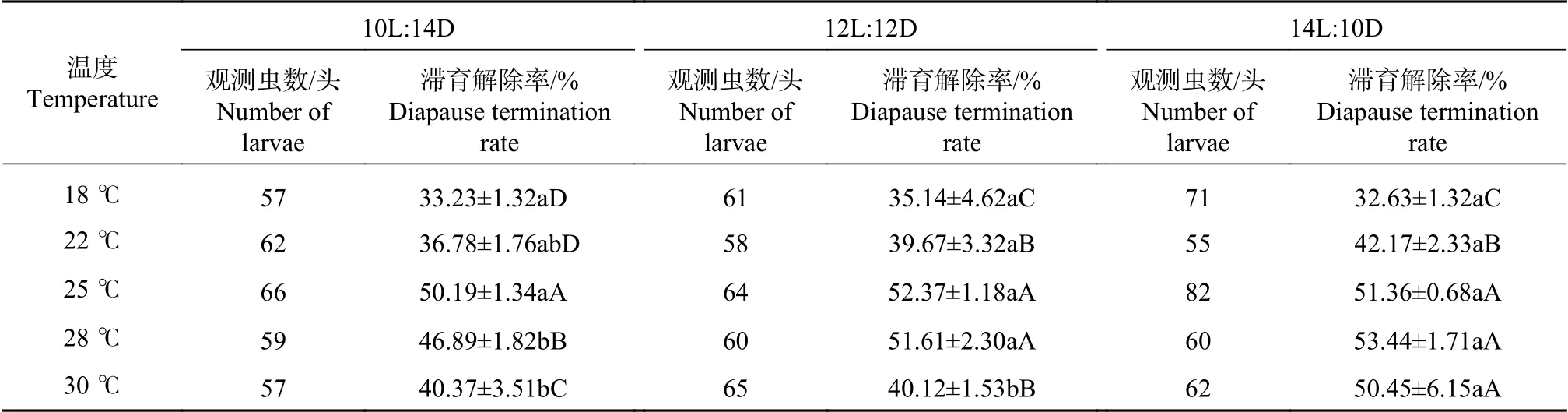
表3 不同温度和光周期对蜀柏毒蛾滞育解除的影响Tab.3 Effectsof temperature and photoperiod on diapausetermination of Parocneria orienta Chao
3 讨论
根据昆虫滞育特性,滞育可分为专性滞育和兼性滞育两类。专性滞育多为一化性昆虫,其滞育的出现有确定的诱导期和特定的虫龄,滞育启动独立于环境。而多化性昆虫多为兼性滞育,其滞育的诱导和解除的环境因素主要有光周期、温度、食料、密度以及湿度等,其中光周期、温度或两者的变化是主要因子[12-13]。蜀柏毒蛾是多化性昆虫,本文以蜀柏毒蛾1龄幼虫作为实验对象,研究了光周期和温度对其滞育的影响。结果表明:在一定范围内蜀柏毒蛾滞育率与温度呈明显的负相关;在环境温度较低条件下(20℃以下),光周期对滞育率作用明显,短光照条件下滞育率较长光照高;温度对滞育解除作用显著,25℃条件下,蜀柏毒蛾幼虫滞育解除率最高;光周期对滞育解除率影响不显著。
温度是影响昆虫滞育的重要因素,主要是在昆虫的夏滞育和冬滞育起到诱导和维持的作用,并且能够决定一些幼虫发育历期跨度较大的昆虫种类进入滞育的发育阶段[5]。同时,有研究表明,特定的低温并维持一定时间的冷刺激是解除某些种类昆虫冬滞育的必要条件[1][4]。但也有大量研究表明,较多冬滞育昆虫的滞育解除不需要冷刺激,其滞育在中性温度条件下就能迅速解除,某些种类滞育解除则需要高温刺激[14]。蜀柏毒蛾属于幼虫发育历期较长的类型,滞育个体(越冬代)幼虫期为208 d,非滞育个体(第1代)幼虫期为85 d,其滞育属于冬滞育。本研究中发现温度对蜀柏毒蛾滞育的影响显著,通过调整环境温度能引发或解除蜀柏毒蛾滞育,其滞育的解除不需要低温冷刺激。这一结果与上述规律一致,即幼虫发育历期长,冬滞育型的昆虫温度因子是影响其滞育的关键因子。
光周期诱导的滞育分为长日照反应型、短日照反应型、短日照—长日照反应型和中间型日照反应型四种类型[15][16]。鳞翅目昆虫中,长日照反应型有马尾松毛虫(Dendrolimus punctatus)[17]、水稻二化螟(Chilo suppressalis)[18]、菜粉蝶(Pieris rapae)[19]和苹果蠹蛾(Cydia pomonella)[20]等,其滞育发生在短日照条件下;短日照反应型有家蚕(Bombyx mori)和梨小食心虫(Grapholita molesta)[21]等;欧洲玉米螟(Ostrinia nubilalis)属于短日照—长日照反应型;桃小食心虫(Carposina niponensis)[16]和黑纹粉蝶(Helicoverpa armigera)[22]属于中间型日照反应型,在中间性光照条件下发育,其他光周期下滞育。通过对蜀柏毒蛾的林间观察和室内光周期实验可看出,低温短日照促进其进入滞育,属于长日照反应型昆虫。
光周期与温度共同作用于昆虫滞育诱导时,光周期反应可以分为温度补偿和温度敏感两种反应类型,光周期决定了温度补偿型反应的滞育诱导效果,温度决定了温度敏感型光周期反应的滞育诱导效果[16]。本次试验结果显示,温度对蜀柏毒蛾滞育诱导起主导作用,光周期需要在一定的温度条件下,才能起到加强滞育诱导的作用。28℃时,无论光周期如何变化都无法诱导蜀柏毒蛾进入滞育。由此可知,蜀柏毒蛾的滞育需要低温和短光照共同作用,其中温度起主要作用,属于温度敏感光周期反应型。
多化性昆虫不同世代间滞育率往往存在差异。如对北京地区玉米螟的研究发现,不同世代间的滞育率存在明显的差异,表现为第3代>第2代>第1代[23];对樟叶蜂(Mesoneura rufonota)人工饲养种群滞育研究发现,其第2代的滞育率显著高于第1代[24]。本次试验研究由于受诸多条件限制,没有进行蜀柏毒蛾不同世代之间滞育的对比研究。但前期研究发现,蜀柏毒蛾越冬代多以1龄幼虫滞育的方式越冬,而第1代卵期、幼虫期及蛹期几乎没有滞育的现象。分析认为这可能是蜀柏毒蛾在进化过程中为了应对环境变化形成了冬滞育的遗传特征,其滞育的主要诱导因子除了温度和光周期等环境因子外,可能有其他更为重要的因子。而这些因子诱导的滞育机制尚不清楚。因此,对于蜀柏毒蛾的滞育机制仍有待进一步研究,如世代、食料、生理因子等对其滞育的影响,以期能明确各类因子诱导和解除滞育的内在机理。
