锰渣堆场优势植物对Mn的富集特点及作用机制
2021-10-11罗绪强
罗 洋, 刘 方, 任 军, 朱 健, 罗绪强
(1.贵州大学 资源与环境工程学院, 贵州 贵阳 550025; 2.贵州师范学院 地理与资源学院, 贵州 贵阳 550018)
由于技术和设备落后、矿石品位低和管理不善等原因,在锰矿的开采和加工过程中产生了大量的废渣[1]。锰渣大多采取露天堆积方式,其中仍残留一定数量的Mn,在雨水淋溶、风力搬运等作用下进入周围环境介质中造成污染[2-3]。黄小娟等[4]的研究结果表明,重庆溶溪锰矿尾渣堆积区土壤受到Mn的严重污染,其平均含量达48 382.5 mg/kg;杨胜香等[5]以花垣矿区4个锰矿点为现场,采用污染指数法对矿区土壤污染状况进行了评价,发现矿区土壤Mn含量较高,是湖南省土壤背景值的8.7倍;陈红亮[6]的研究结果也证实电解锰渣在堆存过程中,Mn会不断从渣中溶出进入周围环境,对动植物生长和人类健康造成危害。因此,采取经济有效的措施对锰渣堆场中的Mn进行治理是生态环境保护的一项基本任务,对当地的可持续发展具有重要意义。
植物在生长过程中既可以通过吸收作用将重金属元素转运累积至地上部分,也可以通过根际效应改变土壤或沉积物中重金属的化学形态,在重金属元素的地球化学循环过程中扮演着重要角色[7]。优势植物是指在一定的地区,植物群落的各个层或层片中数量最多,盖度最大,在群落中作用最为明显的植物种[8]。锰渣堆场虽然存在着基质结构性差,养分缺乏,Mn含量仍然较高等限制条件,但由于长期的物理、化学和生物风化过程,加上自然选择的作用,总有一些优势植物能够适应这种特殊的环境而在堆场上生长[9]。
因此,对锰渣堆场内定居的优势植物Mn富集特征及作用机制进行调查研究,是在该区域开展Mn污染治理工作的前提。优势植物的类型以及对Mn的吸收特点因地理分布的不同而不同。目前,国内对于锰矿废弃地植物修复的研究多集中于湖南、重庆和广西等区域,研究内容主要是Mn胁迫下植物的生理响应、矿区植物种类调查和Mn超富集植物筛选等[10-12],缺乏优势植物生长条件下其根际和非根际基质中Mn含量状况的对比研究。
对矿区基质中Mn形态的探讨也多以无机酸、中性盐和DTPA等单一提取或传统的连续提取法为主,采用模拟植物根系分泌的低分子量有机酸为提取剂的研究鲜见报道。贵州省东部地区锰矿资源丰富,约占全国锰矿资源的50%,锰产业主要以锰矿开采,锰粉加工和电解金属锰为主[13]。该区域已有的研究包括矿区废弃地Mn含量调查与风险评价、锰渣中Mn的溶出规律与改良剂筛选以及锰渣毒性效应等方面[14-16],缺乏植物生长对堆场内Mn迁移的调控效应及作用机理的相关研究。因此,本研究选取贵州省东部地区的松桃县孟溪锰矿采选渣堆场和镇远县青溪电解锰渣堆场上自然生长的优势植物作为研究对象,对植株的Mn含量和基质Mn含量进行了测定。在此基础上,以柠檬酸、苹果酸和草酸等作为提取剂对Mn形态进行分析,探讨植物根系分泌物对基质中Mn有效性的影响,旨在为该区域锰渣堆场的生态修复提供理论依据。
1 材料与方法
1.1 研究区概况
贵州省东部地区属中亚热带季风气候,年均气温在14~19 ℃之间,雨季明显,降水较多,年降雨量在1 000~1 600 mm,日照年均约1 200 h,无霜期长,为260~220 d;地貌以低山、丘陵为主。该区矿产资源丰富,主要有锰、磷、汞、铅、石煤等,其中以锰矿得天独厚,是我国三大锰矿基地之一。调查的采选渣堆放区(锰矿石开采过程中留下的残渣)位于贵州省松桃县孟溪镇,废渣堆放年限大约3~10 a;电解锰渣堆放区(锰矿冶炼过程中在碳酸锰矿粉中加入硫酸溶液后电解锰生产的滤渣)位于贵州省镇远县青溪镇,堆放时间大概为10~15 a。
1.2 样品采集与预处理
(1) 植物样品。课题组于2020年7月对贵州省东部地区的孟溪和青溪锰渣堆场生长的野生植物进行了调查。孟溪锰矿采选渣堆场内自然定居的植物较少,覆盖度为20%左右,全部为草本植物,主要物种有狗牙根(Cynodondactylon)、五节芒(Miscanthusfloridulu)、白三叶(Trifoliumrepens)、一年蓬(Erigeronannuns)和繁缕(Stellariamedia),其中优势种为五节芒和一年蓬;青溪电解锰渣堆场内自然定居的植物相对较多,覆盖度为60%左右,主要物种有草本植物狗牙根、夏枯草(Prunellavulgaris)、五节芒、蒲儿根(Sinoseneciooldhamianus)、一年蓬、野艾蒿(Artemisialavandulifolia)、看麦娘(Alopecurusaequalis)和灌木植物醉鱼草(Buddlejalindleyana),其中优势种分别是五节芒、一年蓬和醉鱼草。在每个堆场内根据与锰渣堆距离的远近分别设置3个样点,每个样点选取6~8株上述优势种植物,用聚乙烯塑料袋封装后带回实验室。根据植物特征,将五节芒分为根、茎和叶3部分,将一年蓬和醉鱼草分为根、茎、枝、叶和花5部分。先用自来水冲洗干净后,再用去离子水清洗3次,待植物表面水分风干后,于105 ℃杀青30 min,然后在60 ℃烘箱中烘至恒重,粉碎后过60目筛装袋备用。
(2) 基质样品。参考文献[17—18]方法,收集0—30 cm厚度层次中根表面5 mm以内的生长基质进行混合,获得根际样品;在相同区域采集距离根表面 10 cm以外的基质混合,即非根际样品。将采集的基质密封后带回室内自然风干,剔除杂物后磨碎过147 μm尼龙筛,装袋备用。
1.3 测定指标与方法
1.3.1 重金属全量 准确称取废渣基质样品0.15~0.30 g(精确到0.000 1 g),在HNO3-HCL-HF-H2O2体系(体积比为3∶1∶1∶1) 下,按120 ℃(7 min)-150 ℃(9 min)-190 ℃(25 min)程序进行微波消解至液体透明澄清[19];准确称取植物样品0.1~0.3 g(精确到0.000 1 g),采用HNO3-H2O2消解体系(体积比为5∶2),按120 ℃(20 min)-160 ℃(20 min)-190 ℃(40 min)程序进行微波消解至液体透明澄清[20]。消解溶液经转移、定容、过滤等程序后,Mn元素采用火焰原子吸收光谱仪(novAA 350)测定,其他重金属元素采用电感耦合等离子质谱仪(ICP-MS)测定。在所有的样品处理过程中,利用标准物质(土壤为GSS系列,植物为GSV系列)、平行样、空白对照等进行质量控制试验,所用试剂均为优级纯。
1.3.2 转移系数和富集系数 植株地上部重金属含量=(茎干重×茎中重金属含量+枝干重×枝中重金属含量+叶干重×叶中重金属含量+花干重×花中重金属含量)/(茎干重+枝干重+叶干重+花干重);生物富集系数(BCF)=植物各器官重金属含量/土壤中重金属含量;转移系数(BTF)=植物地上部分重金属含量/植物根部平均重金属含量。
1.3.3 Mn形态 参考卢豪良等[21]的方法,采用柠檬酸、苹果酸、草酸以及这3种酸的混合液(体积比为1∶1∶1)模拟植物根系分泌低分子量有机酸作为基质可交换态Mn的提取剂,以欧共体标准物质局(BCR)对重金属的连续提取法作为参照,即0.5 g基质中分别加入20 ml 0.11 mol/L的上述有机酸,22 ℃条件下连续振荡16 h,离心(3 000 r/min)。上清液过0.45 μm滤膜后用火焰原子吸收光谱仪(novAA 350)测定其中的Mn,同时设置空白对照。
1.4 数据处理与分析
用Excel 2007进行数据的处理分析,相关指标数据以平均值表示,运用SPSS 26对植株Mn含量和基质Mn含量进行相关分析和显著性检验。
2 结果与分析
2.1 优势植物对Mn的吸收及分布特点
对两个堆场内优势植物各器官中的Mn及其他重金属含量进行了测定,结果详见表1。植物体Mn含量介于34.47~680.70 mg/kg,其中孟溪堆场的五节芒和一年蓬根部、青溪堆场的醉鱼草根部Mn浓度超过了一般植物的正常含量水平上限(400 mg/kg)[22]。植物体内其他重金属元素含量大小顺序为:Zn>Cu>Pb>Cd,均在植物正常含量范围内(Pb 0.1~41.7 mg/kg,Zn 1~160 mg/kg,Cu 0.4~45.8 mg/kg,Cd 0.2~0.8 mg/kg)[23-24]。从分布来看,Mn在一年蓬、五节芒和醉鱼草各器官中的含量均为:根>叶>茎或枝,除五节芒各部位Mn含量均超过100 mg/kg以外,其余植物体内Mn的累积波动较大。在调查期内一年蓬已大量开花,其Mn含量仅次于根部。此外,研究还发现调查区内一年蓬和五节芒对Cu的分布与Mn相似,以根部吸收为主,醉鱼草对Pb,Zn和Cu等元素主要以地上部吸收为主,说明Mn和其他重金属元素在植物体内的分布主要受植物类型的影响。

表1 锰渣堆场优势植物各器官重金属含量
2.2 植物根际效应对基质Mn有效性的影响
由表2可知,调查区内基质中重金属平均含量从大到小顺序依次为:Mn>Zn>Cu>Pb>Cd。其中Mn在松桃孟溪锰渣堆场基质中含量为68 149.50~75 115.58 mg/kg,是贵州省背景值的85.83~94.60倍,在青溪电解锰渣堆场基质中含量在20 217.14~38 045.75 mg/kg之间,是贵州省土壤背景值的25.46~47.92倍。土壤环境质量标准未对Mn元素作出规定,但有研究表明,土壤中Mn含量的适中标准为170~1 200 mg/kg[25],则堆场内基质Mn含量是此适中标准上限值的16.85~62.60倍。因此调查区内Mn严重超标,是最主要的污染元素。此外,两个采样点基质中的Zn和Cu元素也高于贵州省土壤背景值,分别是其含量的1.39~3.17,1.01~2.39倍。在青溪锰渣堆场,还存在Pb局部超过背景值的现象。调查还发现调基质中Zn元素浓度的最大值分别是《农用地风险管控标准》管制值的1.05倍,其余元素含量均在规定值以下。从分布来看,一年蓬、五节芒和醉鱼草根际基质中的Mn含量均低于非根际。除了青溪堆场的醉鱼草(Pb)、一年蓬(Zn)和五节芒(Zn)生长区少部分元素外,其余重金属元素在几种优势植物对应生长基质中的分布均为:非根际>根际,说明在优势植物根系生长条件下,其根际基质中的Mn及其他重金属元素含量与非根际相比总体上表现为下降趋势。

表2 优势植物根际及非根际基质重金属含量的变化
利用4种低分子量有机酸作为提取剂对锰渣堆场基质中Mn的有效性进行了分析,结果详见表3。柠檬酸、苹果酸、草酸和混合酸对孟溪堆场基质Mn的提取效率为分别26.39%~62.85%,39.03%~65.55%,25.72%~64.26%和42.04%~60.47%,表明单一有机酸在该采样点优势植物生长区基质上的提取能力相差较大,而混合酸整体效率较高,变化范围也较小。相比之下,青溪堆场基质中4种有机酸提取态Mn占总Mn含量的比例分别为39.79%~68.71%,26.12%~43.36%,41.57%~78.30%和45.26%~67.28%,除苹果酸外,整体高于孟溪堆场,而苹果酸对青溪堆场所有基质样品中Mn的提取能力均为最弱,说明基质性质是影响该区域Mn形态的主要因素之一。从表3还可以看出,一年蓬和五节芒作为调查区共有的优势植物,其生长基质中低分子量有机酸提取态Mn的含量及比例在两个采样点有所不同。其中孟溪堆场一年蓬生长基质中Mn提取能力的大小顺序为:柠檬酸>混合酸>苹果酸>草酸,而青溪堆场一年蓬生长区提取能力大小则为:混合酸>草酸>柠檬酸>苹果酸。五节芒生长基质Mn提取能力最弱的是混合酸和柠檬酸,最强的在孟溪是苹果酸,在青溪则为草酸。从分布来看,一年蓬根际基质的柠檬酸和苹果酸提取态Mn低于非根际,而草酸和混合酸提取态Mn则大于非根际基质;五节芒根际基质的4种低分子量有机酸提取态Mn占比均大于非根际基质;醉鱼草根际基质的苹果酸和混合酸提取态Mn百分比分别为39.65%和57.08%,大于非根际,而另外两种酸提取态含量则低于非根际基质,各种酸在根际基质的提取能力大小顺序为:混合酸>柠檬酸>草酸>苹果酸,而非根际基质为:草酸>柠檬酸>混合酸>苹果酸。综上所述,在优势植物根系生长条件下Mn的有效性发生了变化,表现为根际基质中的低分子量有机酸提取态Mn所占比例普遍大于非根际基质。
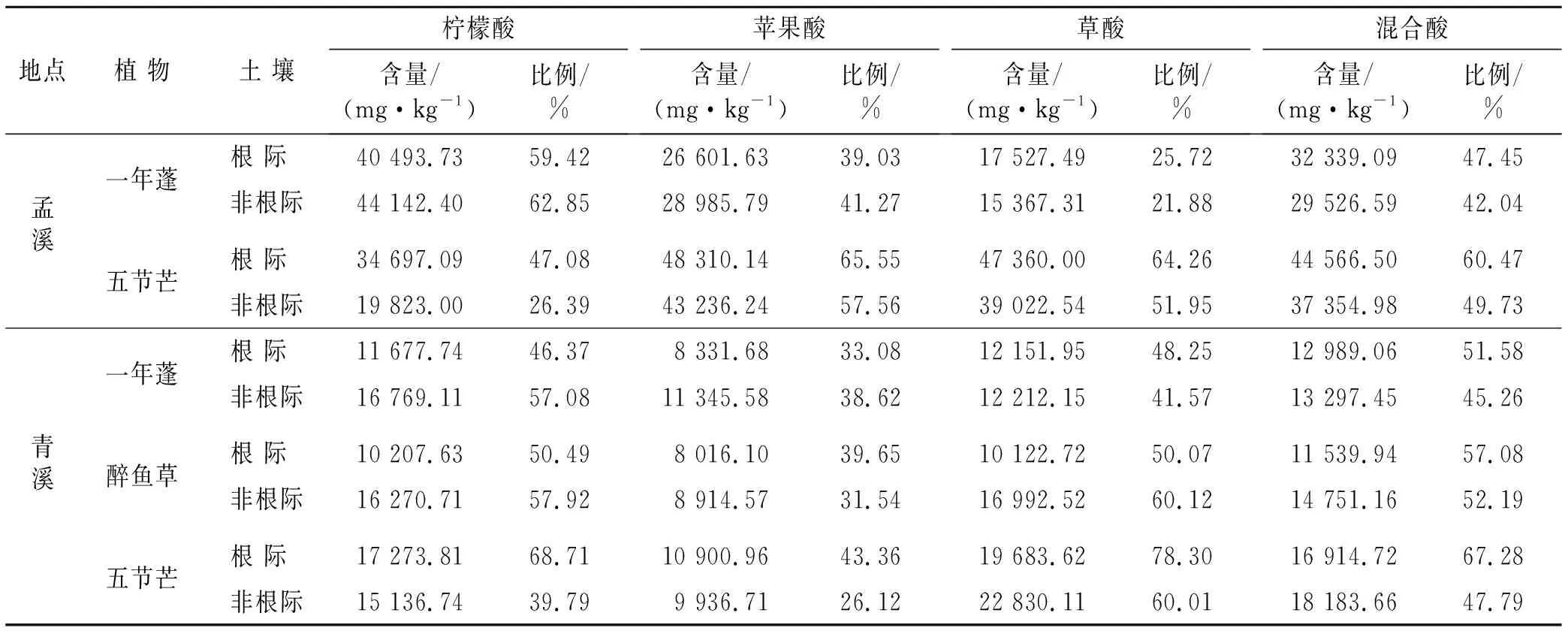
表3 锰渣堆场土壤有机酸提取态Mn含量及比例
2.3 植株Mn的累积与基质Mn含量的关联性
富集系数是反映植物吸收重金属能力大小的重要评价指标,转移系数是反映植物由地下部向地上部转移重金属能力大小的重要评价指标[28]。通过计算发现(表4),锰渣堆场内几种优势植物各器官中重金属含量以Mn元素最高,但对Mn的吸收能力却不强,其富集系数最高为0.031,最低仅有0.001,明显低于Cd,Pb,Zn和Cu等元素。除了青溪堆场五节芒茎部和醉鱼草茎部对Cd的富集系数分别1.091,2.300以外,几种优势植物各器官对其他重金属元素的富集系数都小于1。表4中的数据还显示,调查区内优势植物对Mn的转移系数都低于1,其中由根部转移Mn到地上部能力最强的是醉鱼草,转移系数为0.62,最弱的是一年蓬,在两个堆场的转移系数分别只有0.08,0.23。除了五节芒对Cd、醉鱼草对Cd,Pb,Zn和Cu外,优势植物对其他重金属元素的迁移能力与Mn趋势一致,转移系数均小于1。
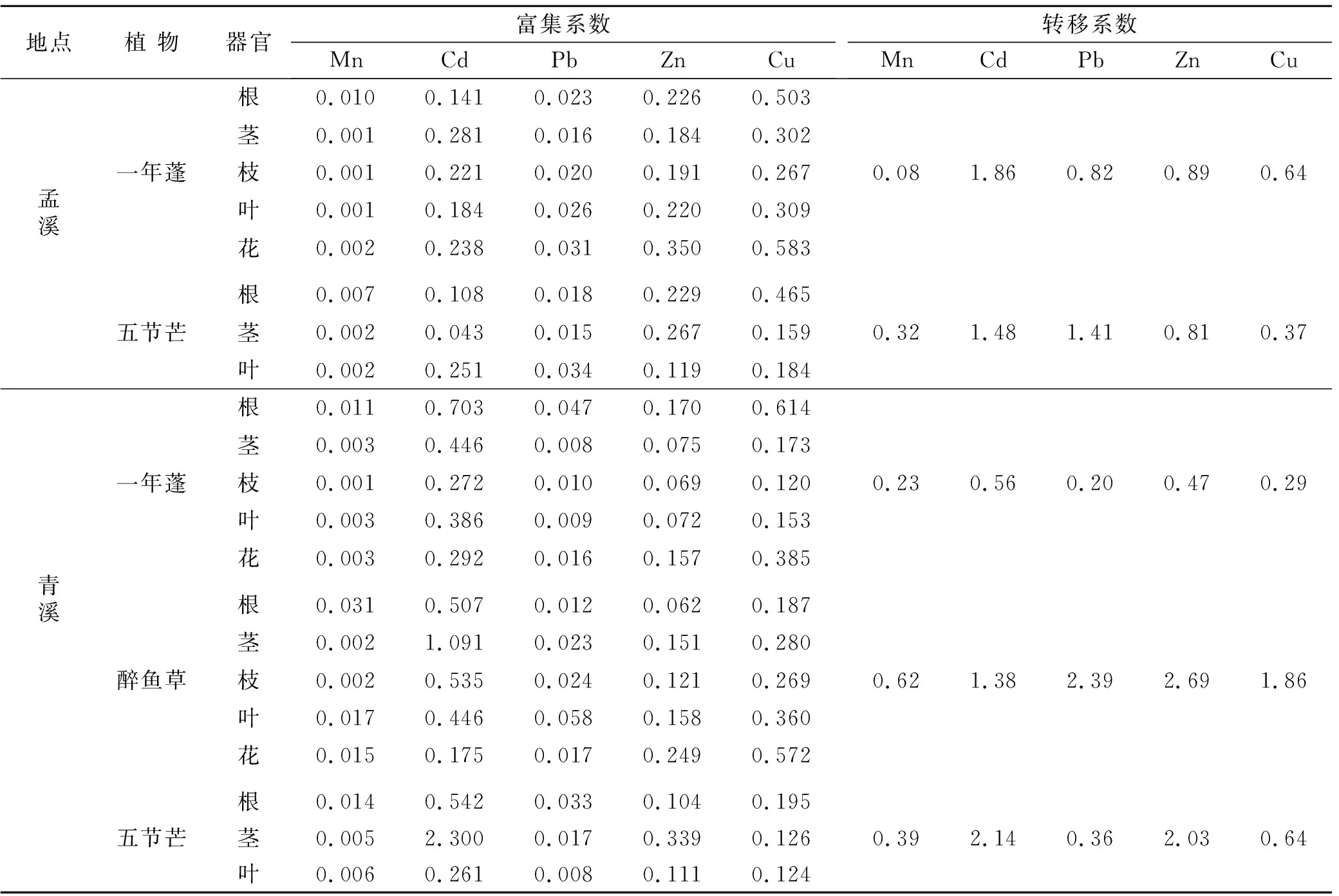
表4 优势植物对重金属的富集系数和转移系数
对4种优势植物各器官中的Mn含量与根际基质中的Mn含量进行相关性分析。由表5可知,植物根部Mn含量与基质中的总Mn和混合酸提取态Mn含量呈显著正相关关系(p<0.05),相关系数分别为0.908,0.819;植物根部Mn含量和基质中的柠檬酸提取态Mn的相关系数达0.969,为极显著水平(p<0.01),说明随着基质Mn含量及形态的变化,植株根部的Mn含量也呈现相同趋势的变化。此外,还发现植物茎和枝部Mn与基质中草酸提取态Mn也呈显著正相关关系(p<0.05)。植物叶和花中Mn浓度虽然也较高,但是由于在不同类型植物(尤其是醉鱼草)内分布波动较大,因此这两部分Mn含量和基质中的Mn含量之间并没有显示出显著的相关性。

表5 植物Mn与基质Mn含量的相关分析
3 讨 论
3.1 优势植物对锰的累积特点
Mn是植物体的必需微量元素,但过量Mn会影响细胞代谢和矿质养分吸收,对植物造成极大损害[29]。唐文杰等[30]对广西3锰个矿区20种主要优势植物组织重金属含量进行了测定,发现植物体内各组织中重金属含量大小顺序趋势为:Mn>Zn>Cr>Cu>Pb>Cd,其中商陆和油茶表现出很强的Mn累积特征。本研究也证实一年蓬、五节芒和醉鱼草等生长在锰渣堆场内的优势植物各器官中的Mn浓度均高于Cd,Pb,Zn和Cu等其他重金属,甚至在个别样品中出现了超过一般植物Mn正常含量范围的现象[31]。Mn在3种优势植物组织中的分布趋势与其他重金属基本一致,主要为:根>花>叶>枝>茎,说明植物根部对Mn的吸收能力相对较强,而被转移到地上的部分主要集中在叶或花等组织上,茎、枝作为输导组织并未有大量的Mn富集[32]。
雷梅等[33]将植物对重金属的吸收机制分为富集型(accumulators)、根部囤积型(root compartments)和规避型(excluders)3类,其具体状况和锰渣特性、植物种类以及自然地理环境等有关。李礼等[34]在对重庆秀山锰矿废弃地进行植物调查时,发现优势植物对Mn的转移系数达0.52~2.86,但富集系数只有0.008~0.025,属于根部囤积型;余光辉等[35]对湘潭锰矿红旗分矿开采区的研究表明,商陆的地上部分Mn含量达到1 564.5 mg/kg,属于富集型。在本研究中,优势植物对Mn的富集系数只有0.001~0.027,对Mn的转移系数也全部低于1,各器官中Mn含量离Mn超富集植物的标准(10 000 mg/kg)也相差甚远,因此所调查的优势植物全部为Mn规避型植物,可通过自身的排斥机制,减少锰渣堆场Mn向植株体内的迁移和富集,降低Mn的毒害作用。在3种优势植物中,五节芒在两个堆场均生长旺盛,具有根系发达、地上部生物量大、各部位Mn含量较高等特点,可作为堆场生态恢复的先锋植物。
3.2 植物根际效应对基质中Mn的作用机制
对优势植物根际和非根际生长基质中的重金属进行研究,既能反映堆场环境的污染状况,又能揭示植物根系活动对基质中重金属总量和有效性的影响。本次调查的两个不同类型的锰渣堆场优势植物生长基质中重金属含量均以Mn最高,远超过贵州省土壤背景值和一般土壤适中范围上限值,潜在生态危害较大,这与朱佳文等人[36]的研究一致。此外,Zn和Cu元素在调查区内也存在超过贵州省土壤背景值以及农用地风险管控筛选值的情况,具有一定污染风险。从分布来看,孟溪锰矿采选渣堆场基质的Mn含量要高于青溪电解锰渣堆场,可能是因为两地的冶炼矿石类型、矿石品位、冶炼工艺、堆存时间和气候条件等因素不同,导致废渣中Mn的含量和迁移特征不同[37-38]。
同一堆场内,一年蓬、五节芒和醉鱼草根际基质中的Mn和其他重金属元素含量均低于非根际基质,其原因可归结为两点: ①植物的吸收作用。优势植物在生长过程中,各器官对Mn和其他重金属均有一定的吸收作用,使植株根际基质中的重金属迁移到根部或地上部,而凋落部分并未完全腐殖质化返回基质,因此造成了总量的减少; ②植物的根际效应改变了Mn的状态,增加了其迁移速率。前人研究表明,植物根系分泌的有机酸等物质可通过低环境pH值、改变氧化还原电位和微生物状态等途径来对重金属元素进行活化反应,使其移动性增加,导致根际土壤或基质中各类或者某一类重金属含量得以下降[39-40]。表3的数据显示,锰渣堆场的Mn形态在根际和非根际基质中存在差异。模拟植物根系分泌的低分子量有机酸对根际生长基质样品的提取率整体上高于非根际基质,说明不同植物的根系在生长过程中通过根际效应活化了基质中的Mn[41-43],促进了植物根部和地上部对Mn的富集,使根际基质Mn含量低于非根际基质。
本研究对贵州省东部地区两个不同类型锰渣堆场内的优势植物和生长基质Mn含量进行了调查,初步分析了优势植物对Mn的富集特点及其作用机制。然而,在分析Mn形态时所采用的有机酸浓度为0.11 mol/L,和实际根际土壤溶液中的含量不一定匹配,也缺乏关于在优势植物不同生育期内堆场Mn的动态监测及Mn耐性机制等问题的探讨。因此,在今后研究中,还需要从植物根系分泌物测定、植物生长条件下堆场基质理化性质和微生物特性变化、植物生理、改良剂—植物联合运用、植物群落修复模式等方面进一步开展工作,为锰渣堆场的污染治理和生态恢复提供依据。
4 结 论
(1) 锰渣堆场上生长的优势植物体内Mn含量在34.47~680.70 mg/kg之间,在一年蓬、五节芒和醉鱼草各器官中的分布顺序为:根>叶>茎或枝。
(2) 锰渣堆场优势植物生长基质中的总Mn含量在20 217.14~75 115.58 mg/kg之间,其分布主要表现为:根际基质>非根际基质;植物的根际效应提高了Mn的有效性,使根际基质中的低分子量有机酸提取态Mn百分比普遍高于非根际基质。
(3) 锰渣堆场优势植物对Mn的富集系数和转移系数均小于1,为Mn规避型植物;优势植物各器官中,以根部Mn含量与生长基质中的总Mn含量以及柠檬酸提取态Mn含量相关性最高,分别达显著和极显著水平。
