树干甲烷的研究进展
2021-10-10丁九敏
郭 亮,丁九敏,徐 侠
(南京林业大学生物与环境学院,南方现代林业协同创新中心,江苏 南京 210037)
IPCC(Intergovemmental Panel on Cimate Change)发布的2018特别报道指出,全球平均升温1.5 ℃,将会对生态系统和气候造成严重的影响,导致热浪和强降雨等事件频繁发生,而限制升温的关键在于控制温室气体的排放[1-2]。甲烷 (CH4) 是仅次于CO2的重要温室气体,在百年时间尺度上的温室效应潜力是CO2的28~34倍[3],对全球变暖的贡献占20%~30%[4]。目前,大气中的CH4含量为1.8~1.9 mL/m3[5],这是其排放与消耗之间平衡的结果[6]。大气中的CH4有70%~80%是生物来源[7],它由产甲烷菌在缺氧环境中分解土壤有机物产生。全球CH4每年的排放量为539~609 Tg, 这些数据都是基于现有的全球碳循环模型而估算的,具有相当大的不确定性[8-9];而这些不确定性来自现有的数据对CH4源和汇测度的不全面性。
热带森林基于土壤排放的估算和卫星CH4通量之间的差异报告引发了人们对植物表面作为一个被忽视的来源的兴趣[10-11]。以前的研究更多关注的是森林土壤表面CH4动态,而忽略了植物表面所介导的CH4动态。有文献表明,仅植物介导的CH4排放源可能占全球CH4排放的5%~10%,甚至更多[12-13]。此外,高山森林或者旱地森林[14-15]大多数时候被认为是CH4的汇,但是也有研究表明这部分森林中地上植物的表面都有可能是CH4的源或汇[16-17],这主要是因为以前的研究大多数没有关注地表植物所介导的温室气体通量。通过现有的研究结论可知,陆地生态系统CH4动态比以前认为的复杂得多,这是由活树(树干、枝及叶)和死树介导的植物、微生物和非生物过程的组合[18]。这些研究结果对于准确评估全球CH4通量具有重要意义。鉴于此,笔者综述了树干 (除树干外的植物表面CH4通量相对较小[19]且研究较少) CH4的动态变化及其变化机制,并提出未来的研究方向,促进对全球碳循环和森林生态系统平衡的整体理解,以期预估森林CH4动态对未来全球气候变化的响应,并为适应全球气候变化的森林管理体系提供借鉴。
1 树干CH4的来源
树木释放的CH4来源是现阶段迫切需要解决的一个问题,这有助于预测未来气候变化背景下的全球碳循环。现有研究表明,通过树干释放出的CH4是来自土壤[20-23]或者心材[11, 19, 22,22-25](图1)。这其中的难点就在于区分树干释放的CH4到底是心材厌氧环境中产甲烷菌的产物[11],还是土壤中产生的CH4转运或者扩散到树干中从而释放到空气中[20]。然而,在不同生态系统、不同树种中树干CH4的来源到底是哪种产生机制占主导尚不清楚。
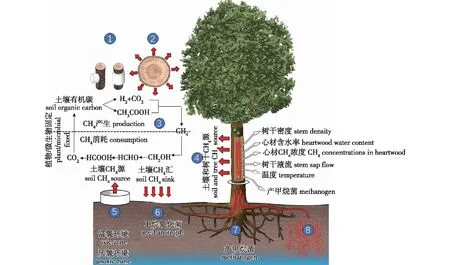
图1 树干CH4研究的概念框架[7, 18, 25]Fig. 1 Conceptual framework of research for stem CH4
1.1 心材源CH4
树干由外至内是由树皮 (皮层、韧皮部等)、边材和心材3个部分组成,这3个部分的CH4浓度逐步增加,但心材部位浓度非常高,表明树干CH4通量可能与心材CH4的含量有关 (心材产生的CH4径向扩散到树干表面)[19]。大多数森林中活木的心材中都或多或少会产生CH4[11]。树干心材中的CH4[11]和产甲烷菌[19, 24]被认为是心材中产CH4的直接证据。产甲烷菌主要有两种呼吸途径,即醋酸发酵(CH3COOH → CO2+ CH4)或消耗CO2(4H2+ CO2→ CH4)。研究发现,这两种情况都发生在活的树体木材中[18, 26]。Wang等[11]利用同位素标记法测定13C-CH4,发现在杨树中通过消耗CO2产生CH4。最新研究表明,部分树种心材中CH4浓度与心材含水量呈幂函数关系,在心材中出现了一个水分含量的阈值,超过这个阈值就会产生大量的CH4[11]。这个阈值可能恰好给微生物生产CH4提供了所需的厌氧环境,这也是心材产CH4的力证。2012年Covey 等[27]对大西洋沿岸温带森林的调查发现,无论在低洼地还是高地,树干CH4浓度都是大气浓度的好几倍,同时心材中存在大量的腐生真菌和古生菌。真菌感染可能导致树木的茎部充满CO2,从而使环境(缺氧或低氧化还原条件)更适合产CH4微生物[25]。这些结论都表明树干表面CH4的来源可能正是树干内部微生物。此外,也有研究指出树干CH4通量与土壤湿度、土壤温度或地下水位深度没有统计学上的关联,这也间接说明了树干CH4通量可能与土壤环境关系不大。
1.2 土壤源CH4
树干的部分体积由气体占据,在被子植物中约占心材的25%,在裸子植物中约占50%[28]。各种充气组织是氧气、CH4等气体在树干中转运的主要途径。土壤产生CH4的树介导运输途径是通气组织,这是一种特殊的组织,其特征是暴露于低氧土壤条件下,在根和茎中形成增大的气体空间[29-30]。通气组织在湿地生态系统植物中普遍存在,是土壤和大气快速转运或者交换气体的组织[31]。Rice等[32]在温室条件下进行的阔叶树种试验表明,土壤生产可能是树干CH4排放的来源。在森林湿地中,Pangala等[33]报道了树干CH4排放与温度、地下水位深度和孔隙水中CH4浓度之间的相关性,认为CH4通量可能受湿地土壤中CH4产量的驱动。虽然树木在自由排水、较高海拔条件下不能形成通气组织,但它们仍然通过连接的充满气体的孔隙空间输送气体[34]。Megonigal等[35]在高地森林的研究认为土壤是CH4的汇,而树木是CH4的来源,这一观察结果表明,树木可能提供了一种通过木质组织的通量途径[13],避开了含氧土层中的甲烷氧化菌。此外,Machacova等[21]在北方森林的研究中也发现松树可能通过蒸腾流将土壤深处产生的CH4输送至气孔释放到大气中,从而防止上层土壤中CH4的消耗,这也是生态系统中以CH4为主的碳动态平衡的一部分。
在不同地区、不同树种条件下,现有树干CH4通量研究所得出的结论不尽相同。理解CH4生产相关的微生物和生化途径是理解树干CH4排放机制的关键步骤之一[25]。同时在评估微生物的作用时,从植物材料中提取微生物DNA或RNA是一项挑战[24, 36]。基于扩增和宏基因组的方法都可以提供有用的工具来描述植物内的一般微生物组,包括细菌、古生菌和真菌。扩增子方法最近被用来表明产CH4的古生菌和厌氧细菌可能支配美洲黑杨心材CH4的生产[24]。除此之外,同位素技术对于揭示CH4的来源也是一种有力的手段[37]。通过同位素标记的底物以及最终收集到CH4的标记物浓度[19],可以相对准确测得上述两种不同CH4来源到底是哪一种占主导或兼之。
2 树干CH4通量及其影响因素
森林生态系统CH4通量的量化非常具有挑战性,因为森林界面是CH4产生、消耗、运输和排放的复杂组合体[18]。研究表明树枝和树叶相对于树干的CH4排放量相对甚小,因此树干作为森林生态系统的一部分其CH4通量监测有必要单独进行。现有关于树干CH4的研究监测大多数是利用树干静态箱-气相色谱法[11, 38-40](图1),这是一种基于测定土壤CH4通量改造而成的方法。大多数的监测数据显示 (表1),树干CH4一般是净排放,一部分是零排放,极少数的是净消耗。树干表面CH4通量具有较大的时空差异性,这些差异主要来自树种、树龄、组织类型、立地特征和环境条件[41-48]。

表1 不同生态系统的树干CH4通量Table 1 Methane flux from tree trunks in different ecosystems
2.1 树种对树干CH4通量的影响
不同树种树干CH4的排放有很大的差异[19-20, 39, 43]。如表1所示,不同树种间树干CH4通量大小不一。这可能是由于某些物种有通气组织(或者通气组织更多)、或心材更容易腐烂、或木材密度不同,因此,在木材气体扩散率上存在巨大的差异。较高的木材密度可以通过减缓氧气扩散和增加茎的缺氧来提高CH4的产生[18]。Wang 等[11]的研究结果显示,22种山地森林树种按树干内CH4浓度持续高、持续低和可变分为3个类群。在不同树种之间,皮孔的存在、数量、类型、开放程度、发育阶段和面积各不相同[49]。同时树干皮孔密度的任何变化都可能影响茎和根通气组织的发育,从而可能改变CH4的转运速率[50]。Pangala等[50]研究表明,茎皮孔密度与树木介导的CH4排放之间存在很强的正相关关系。裸子植物和被子植物的解剖学差异 (如管胞等) 也会影响树干内微生物的扩散或定植,从而影响CH4的排放[25]。此外,不同树种心材中产甲烷菌的碳源类型差异会造成CH4通量的差异。根据对土壤的研究,当碳来源是高度芳香或顽固碳时,CO2的消耗占主导地位,而醋酸发酵则在小分子碳水化合物占主导地位时发生[51]。所以对于不同树种CH4通量的监测对全球温室气体的整体预算非常重要。
2.2 树龄对树干CH4通量的影响
树木直径是对树木大小的度量,通常被解释为物种年龄的替代指标,对排放可能有直接的物理影响(例如扩散)[52]。随着树龄的增加,产CH4种群有更多的时间在树干心材中定居。不同生态系统类型中,小树和大树的排放往往不同。在森林湿地中,幼树的CH4排放量往往高于成熟树[12, 33],而在山地森林中,情况往往相反[11, 20]。除此之外,有证据表明,其他与林龄相关的环境因素,如pH下降,可能会限制老龄树干中微生物群落产CH4的潜力[24]。因此需要测量更多不同直径的树木[18]及不同种类和年龄的树木,采用分层设计以描述从地块到生态系统尺度树干CH4排放的空间变异性。
2.3 组织类型对树干CH4通量的影响
树干CH4排放的研究显示,随着树干高度的增加,CH4排放总体呈下降趋势。特别是当土壤被认为是茎干排放CH4的可能来源时变异性很强[12, 19-20, 33, 42]。这也是缺氧的泥炭土是茎排放CH4主要来源的间接证据。由于边材和心材的厚度随着树干高度减小,树干基部外力造成的损伤也会造成树干CH4通量的空间差异[25]。有关树枝和树叶表面的CH4通量也有少量研究[12, 19, 53],但大多数从无净通量到净排放不等,而且量微。因此,需要在多个树的不同高度上进行更多的测量,以便更好地量化树干CH4通量的模式。
2.4 立地特征和环境条件对树干CH4通量的影响
尽管存在潜在的树木特异性排放,但环境条件在控制整体树干排放的空间和时间变异性方面发挥着重要作用。不同土壤湿度条件、土壤温度、地下水位或土壤孔隙水CH4浓度的研究表明,不同物种的茎部排放存在差异。在高地森林和森林湿地,地下水位深度是一个控制土壤氧气含量和氧化还原电位的主变量[54],是调节旱地土壤CH4消耗[55]、湿地土壤CH4排放[56],湿地和高地树干CH4排放[20, 33, 41]的主要环境因子。一般来说,湿地树木的CH4排放量要高于旱地树木[10, 12, 38, 41],这可能是由于湿地土壤孔隙水CH4浓度较高,也说明了森林湿地中土壤来源对树干CH4排放量的贡献很大。这其中最具代表性的当属亚马逊热带雨林,由于其特殊的高温高湿环境条件,平均树干排放量比迄今为止研究的任何其他系统都高出多个数量级[12],说明气候和环境条件对树干CH4通量具有很大的影响。然而,相关的基础性研究相对较少,需要大量的跨物种和生态系统的研究,以便更好地描述陆地生态系统树干CH4排放的异质性。
3 树木CH4通量对陆地生态系统碳预算的影响
现有的研究结果指出土壤CH4通量已不能等同于森林生态系统的CH4通量,但量化CH4通量是一项挑战,因为森林是CH4生产、消耗、运输和排放的复杂组合体[18]。有关植物所介导的CH4排放研究是从Keppler等[57]的研究之后才逐渐开展的。到目前为止,由于缺少大量的区域基础数据,对全球树木贡献的CH4预算估计都不准确[16, 58]。现有的研究结论仅代表了整个陆地生态系统中极小的一部分,但是意义重大。Pangala等[12]在亚马逊的研究表明树木的排放量相当于整个北极地区的CH4排放量,约占全球湿地CH4排放总量的15%。同时,此研究结果也表明,连同已经确定的排放路径 (土壤释放),亚马逊雨林占全球湿地CH4排放量的1/3[12],这比以前报道的要大得多,可能对亚马逊地区产生更大的影响,并且对全球大气中CH4浓度变化产生的影响比想象的要大。张雨雪等[38]对河滩秋茄林的研究结果显示中潮滩树干CH4通量约占整个区域CH4通量的1/3。此外,Jeffrey等[47]对红树林的研究表明活树或者死树的树干也向大气排放了大量的CH4。在温带森林湿地中,白桦等树干的排放占生态系统CH4排放的27%[33]。总的来说,在湿地或者热带雨林这样的潮湿环境中,树干所介导的CH4排放量在该地区总的CH4排放量中占比很大,因为土壤和树木都是碳源,先前对森林湿地的研究似乎低估了这些无处不在的生态系统的碳预算。
然而在自由排水的山地森林中,树木和土壤通常对森林CH4预算有相反的影响,土壤通常主要作为净汇,而树干主要作为净源。山地树木作为CH4来源的后果具有潜在的重要性,因为山地土壤是陆地上最大的单一CH4库,净消耗量估计为36 Tg[58]。Wang等[19]在温带山地落叶林的研究发现树干CH4排放量相当于土壤CH4年消耗量的63%。这一估计值可能是树木对自由排水土壤的贡献的上限,推测是因为大多数杨树心材中含水率都很高,而其他物种的树干CH4通量估计相对较低。在芬兰一个潮湿的高地森林,树干CH4排放抵消了35%的土壤汇[21]。但是也有部分研究指出,树木的CH4排放对山地森林的CH4预算几乎没有影响。Pitz等[22]对北美温带森林的研究显示树木的排放抵消了生长季土壤中16%的CH4汇。Warner等[43]的研究结果显示树干的CH4排放抵消了净土壤和粗木制残体吸收的3.5%的汇。最后,部分地区树木的CH4净效应也会增加土壤的碳汇。在一项涉及树干通量的田间试验中,重新种植的林分CH4吸收量是未种植林分的2倍[45]。这一结果可能是由树木极低的CH4排放率和未知因素 (如由呼吸驱动的土壤水势降低或下层草本植物的存在) 导致的土壤CH4吸收增加共同造成的。综上所述,在不考虑树干CH4通量的前提下,湿地生态系统或漫水环境的CH4释放量部分被低估,而旱地或者高山森林中CH4的吸收量部分被高估。
4 结 语
树干在碳循环科学中的作用是一个新兴的研究领域,对理解不断变化的环境条件下全球碳循环产生重大影响。越来越多的关于森林生态系统中CH4动力学的研究表明,它们是比以前认为的要复杂得多的生物地球化学环境,而且以前对土壤过程的关注不足以清晰地理解森林与温室气体的平衡。建议未来的研究应同时探索不同的方法和挑战,阐明树干CH4产生和通量变化的机制,包括高频测量 (天、月、季、年) 或景观尺度的调查活动,以考虑区域和全球尺度。特别重要的是要结合生物地球化学、木材解剖学和树木生理学的理论和实践知识,因为基于这些的综合分析可以了解调节树木表面CH4生产和交换的机制。同时,需要进一步的研究来完善对整个生态系统的评估,以确定最合适的尺度,并解决树木CH4通量路径之间的差异。目前,实验室研究可以分离出CH4生产或消耗的特定途径,但无法捕捉到驱动原位通量的实质性时空变化尺度。除了通量测量,还需要对现有跨学科的技术进行深思熟虑的集成。除非进行更多的综合经验研究,并开发和测试基于过程的模型,否则森林对全球CH4动态的贡献仍将难以预估。
