贵州茂兰喀斯特森林两树种叶片气孔形态特征及其对蒸腾的影响
2021-10-10丁访军谭正洪廖立国崔迎春
程 娟,丁访军,谭正洪,廖立国,周 汀,崔迎春
(1.海南大学生态与环境学院,海南 海口 570100;2.贵州省林业科学研究院,贵州 贵阳 550005;3.贵州大学生命科学学院,贵州 贵阳 550025)
贵州地处西南高原属于亚热带季风性湿润气候区,也是中国石漠化尤为明显地区[1]。为改善该地区的石漠化环境,在绿化树种选择、抚育及管理上进行了相关研究[2],但由于地处典型的喀斯特地貌区,岩石透水性强、土壤储水能力低、地下水文过程复杂,加之大气降水时空分布不均[3],目前在治理石漠化问题方面,尤其是适应性树种的筛选上仍无定论。一些森林植被中尤其是混交林,共存物种受水分条件的限制而不能维持正常生长或再生,长此以往会导致植被和生态系统退化[4-5]。植物完成正常的生长就必须进行蒸腾和光合作用,要完成这一过程离不开与外界环境进行气体交换,如CO2和H2O等。陆地植物与大气之间气体交换由气孔控制[6-7],气孔是反映光合作用、生产力、生态水文和气候胁迫的关键驱动因子,植物通过改变气孔数量及其开闭来调节叶片蒸腾速率和水势;气孔开放的变化可以显示出与植物周围环境条件的适应性关系[8-9],包括气孔密度、大小和形状的变化,而气孔在不同的植物中表现出不同的形状、大小和数量,不同植物的气孔形态特征存在着较大差异[10-11]。目前从气孔自身的形态特征研究发现,气孔密度越小,气孔开度越大,气孔导度就能保持相对稳定,满足自身正常的生命活动需求[12-13]。同时相关研究认为,物种气孔密度与气孔长度、气孔开度及叶面积有一定关系,但不同物种之间存在较大差异[14],气孔数量和形态及其调节用水效率对物种正常的生命活动至关重要。
植物进行生命活动离不开水的参与,水分只有在蒸腾拉力作用下[15]才能被运输到不同的组织器官,保障其完成正常的生命活动,已有研究表明,树干液流可以作为重要生理指标表征植物蒸腾[16]。茂兰喀斯特地区森林树种的蒸腾特征及其与环境因子的关系已有一定研究[17-19],但仍缺乏气孔形态特征对喀斯特地区林木蒸腾的影响,环境因子与气孔形态特征之间的关系,以及它们之间相互影响等方面的研究报道。为此,以贵州茂兰喀斯特地区森林生长季常绿树种胀果树参(Dendropanaxinflatus)和落叶树种瓜木(Alangiumplatanifolium)为研究对象,开展树干液流、环境因子及树种叶片气孔形态特征的观测,分析不同树种的蒸腾与叶片气孔形态特征,及在相似环境条件下叶片气孔形态特征变化对其树干液流的影响,进一步探讨在茂兰喀斯特生境下,叶片气孔对于林木蒸腾需求的响应及其环境因子的影响,从而揭示环境因子影响树木蒸腾的机理,了解类似气候条件下相似物种对外界环境条件的基本调节策略,为茂兰喀斯特地区森林的保护及石漠化治理中的树种选择、抚育、管理等提供理论参考[20]。
1 材料与方法
1.1 研究区概况及树种选择
试验地位于贵州茂兰国家级自然保护区(107°52.2′~108°5.7′E,25°9.3′~25°20.8′N)的喀斯特森林生态系统定位观测研究站,属中亚热带季风湿润气候区;地貌以峰丛洼地为主,海拔平均在430~1 078.6 m,年平均气温18.3 ℃,年平均降雨量达到1 752.5 mm,集中分布在4—10月。保护区内母岩为碳酸盐岩,所成土壤为石灰土,裸岩率在80%以上,土壤稀少,土层薄且不连续,土壤透水性强,地下水文过程复杂,生境异质性大,具有典型喀斯特生境特征。
对原生乔木林群落类型设置固定样地(30 m×30 m),其中胀果树参(D.inflatus)属于常绿小乔木,生长在海拔740~1 800 m的常绿阔叶林或灌丛中,在贵州主要分布于荔波及独山地区;瓜木(A.platanifolium)属于落叶小乔木或灌木,生长于海拔2 000 m以下的向阳山坡或疏林中,在中国分布广,该两树种在茂兰喀斯特顶级群落里均存活有大量成熟植株。根据调查结果,选取接近样地内的平均胸径且生长良好、树干通直、无病虫害的该两树种作为监测样树,每个树种各选3株,基本特征见表1。

表1 贵州茂兰喀斯特研究地两样树基本特征Table 1 Basic characteristics of two species sample trees in Maolan Karst research area,Guizhou Province
1.2 边材面积及树干液流测定
在生长季5月(12、13、14、15、16日)选用3 cm长的热扩散探针(TDP-30,Dynamax Inc., Houston, USA)连续5个晴天,在8:00—17:00监测胀果树参和瓜木的树干液流速率(安装高度均为1.3 m),每个树种各选3株,为了尽可能防止外界环境对传感器的影响,用太阳膜和锡箔纸包裹探针和树干,并用胶带密封好。将TDP探针缆线与CR 1000数据采集器连接,设置数据采集的间隔期为10 min;结合Granier等[21]方法的经验公式无量刚J=0.011 9×[(ΔTmax-ΔT)/ΔT]1.231。经换算得树干边材瞬时液流速率[JS,g/(m2·s)]。式中:ΔTmax为上下探针间的最大昼夜温差,℃;ΔT为瞬时温差,℃;1.231为校正系数。
在样地外分别选取3株不同胸径的胀果树参和瓜木,在树干胸高(约1.3 m)处用生长锥钻取样芯。将样芯带回实验室后,用游标卡尺分别测量样芯的心材宽、边材宽、半径和皮厚等值。根据决定系数[19]R2=0.789 8计算样木边材面积和胸径,建立了边材面积与胸径的回归关系式:
SA=0.074 26×D1.807 7。
(1)
式中,D为样木的胸径,cm。
基于小时尺度下的树干液流速率(FS):
FS=JS×SA×3 600。
(2)
式中:FS为树干液流速率(液流通量),g/h;SA为胸径处边材面积,cm2;3 600为时间换算系数[17]。
1.3 环境因子测定
环境因子(辐射、温度、湿度)利用安装在样地内观测塔上的Campbell自动气象站测定,自动气象站距离地面10 m左右,与树干液流采集水平距离在10 m内,测定及数据采集时间与树干液流同步。测定因子包括:太阳辐射强度(Rn,W/m2)、大气温度(Ta, ℃)、空气相对湿度(RH,%)。采用水汽压亏缺 (vapor pressure deficiency,VPD,公式中以VVPD表示)综合表达气温和空气相对湿度的协同效应,计算公式为[22]:
(4)
1.4 叶片气孔形态解剖结构的测定
在试验期间,每天从8:00开始采集叶片,每隔1h采1次,直至17:00。由于该森林植被类型属于阔叶林,树冠不同树种枝叶交错复杂,为了更好地反映在自然气候状态下植物叶片的生理活动特征,从每个物种不同的3个个体上分别采集阳生、健康的叶片,放入装有液氮的恒温箱中带回实验室立即放进干燥皿干燥,用电子显微镜(EM—30,COXEM公司)观测叶片,在×500、×1 000、×2 000、×5 000下拍摄一定数目的视野图片,用ImageJ图形处理软件统计视野内的所有气孔个数,用来计算气孔密度(stomatal density,SD)即视野中单位叶面积的气孔数目,利用ImageJ软件计算气孔器大小(stomatal apparatus size,SAS),具体为:长轴长/短轴长;气孔长度(stomatal conductance,SC)为保卫细胞长度以及气孔开度(stomatal openness,SO)[12,23-24]。
1.5 数据处理
使用Excel 2010软件对数据进行预处理,采用Origin 2018软件进行数据方差分析、相关分析、回归分析并作图,数据处理为平均值±标准偏差。
2 结果与分析
2.1 两树种树干液流速率日变化特征
两树种树干液流速率日变化曲线见图1。从图1可以看出,落叶树种瓜木蒸腾作用开启后快速增强,在13:00左右达到最强,之后快速减弱。常绿树种胀果树参的蒸腾作用同样在开启后快速增强,在12:00左右达到最强,之后快速减弱;从它们的均值来看,落叶树种瓜木的液流速率[(585.25±53.46) g/h]显著高于常绿树种胀果树参的[(384.83±39.12) g/h](P<0.05,df=48,F=7.42),说明落叶树种瓜木比常绿树种胀果树参的蒸腾能力强。

不同小写字母表示差异显著(P<0.05)。下同。Different letters indicate significant difference(P<0.05). The same below.图1 胀果树参和瓜木树干液流速率日变化曲线Fig.1 Diurnal variation curves of stem sap flow rate of evergreen Denchropanax inflatus and deciduous Alangium platanifolium
2.2 两树种叶片气孔形态特征
两树种相似环境条件下,8:00—17:00时叶片气孔形态特征均值比较见图2。
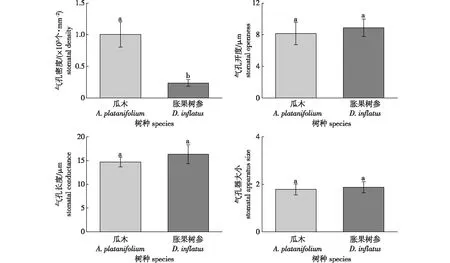
图2 胀果树参和瓜木叶片气孔形态特征Fig.2 The morphological characteristics of leaf stomata of evergreen D. inflatus and deciduous A. platanifolium
由图2可知,落叶树种瓜木的气孔密度(SD)(1 005.08±80.99)个/mm2显著大于常绿树种胀果树参的SD(237.16±21.67)个/mm2(P<0.05,df=48,F=7.08),但落叶树种瓜木的气孔开度(SO)(8.27±0.16)μm小于常绿树种胀果树参的SO(8.89±0.18)μm,且落叶树种瓜木的气孔长度(SC)、气孔器大小(SAS)分别为(14.67±0.31)μm和1.78±0.06,常绿树种胀果树参的SC、SAS分别为(16.34±0.77) μm和1.87±0.13,落叶树种瓜木的SC、SAS均小于常绿树种胀果树参,但两树种的SO、SC以及SAS均无显著差异(P>0.05,df=48,F=2.65),说明两树种气孔形态特征之间的差异来自气孔密度。
不同树种叶片SD与SO、SC之间的关系见图3,两树种的SD与SO及SC之间均存在线性关系,且呈显著负相关关系(P<0.05)。表明同一树种叶片的SD和SC会相互影响,SD大者,SO及SC相对较小,同时表明当叶片气孔孔径发生变化时,植物气孔密度也会作出相应的反应以保障气孔导度的相对稳定,满足自身正常的生理活动需求[14]。
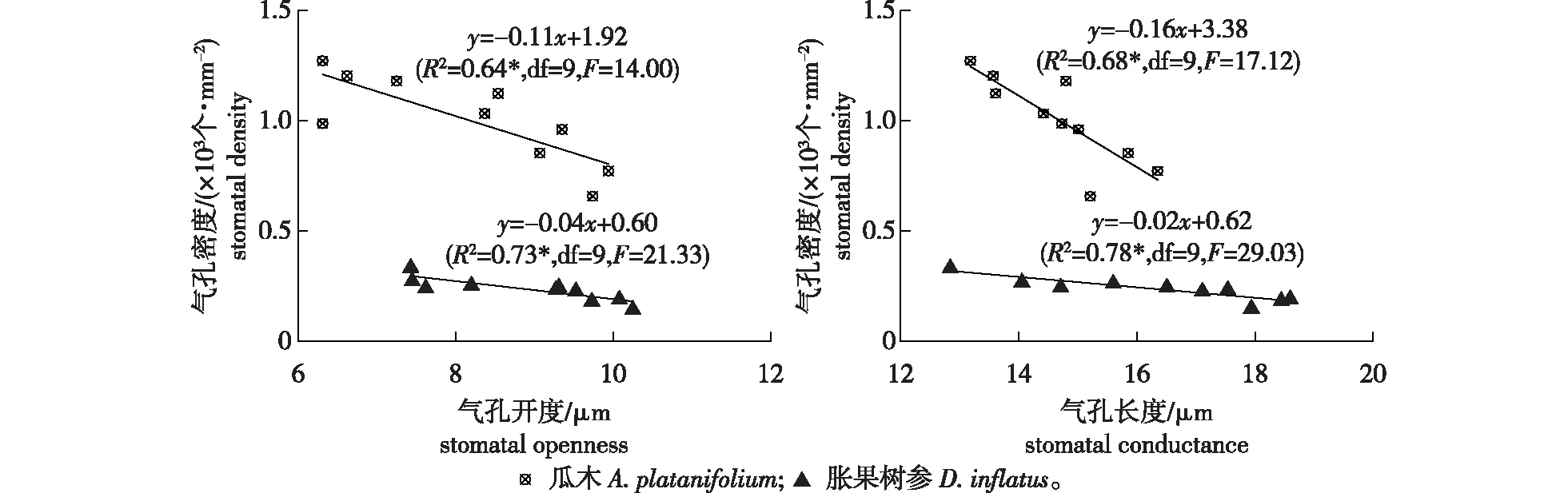
注:*.P<0.05.图3 胀果树参和瓜木叶片气孔密度(SD)与气孔开度(SO)和气孔长度(SC)的相关关系Fig.3 Correlations of stomatal density(SD) with stomatal openness(SO) and stomatal conductance(SC) of evergreen D. inflatus and deciduous A. platanifolium
2.3 叶片气孔形态对树干液流速率的影响及与环境因子的关系
通过对落叶树种瓜木和常绿树种胀果树参气孔形态特征与树干液流速率的相关性分析发现(表2),两树种叶片的气孔密度(SD)与液流速率(FS)均呈极显著正相关(P<0.01,df=48,F=16.03;P<0.01,df=48,F=32.10),说明叶片SD越大的落叶树种瓜木其蒸腾越强。从两树种叶片气孔形态特征与环境因子作相关分析发现,落叶树种瓜木叶片的SD与太阳辐射强度(Rn)和大气温度(Ta)呈显著正相关(P<0.05,df=48,F=7.66;P<0.05,df=48,F=5.50),SC与Rn和Ta呈显著正相关(P<0.05,df=48,F=13.06;P<0.05,df=48,F=4.59),SO与Ta呈极显著正相关(P<0.01,df=48,F=6.10),说明Rn是影响落叶树种瓜木气孔形态变化的主要环境因子。而其FS与水汽压亏缺(VPD)呈极显著正相关(P<0.01,df=48,F=21.30),与Rn呈显著正相关(P<0.05,df=48,F=4.56);常绿树种胀果树参叶片的SD与大气温度(Ta)呈显著正相关(P<0.05,df=48,F=5.02),SC与Ta呈显著正相关(P<0.05,df=48,F=6.32),SC与Rn呈极显著正相关(P<0.01,df=48,F=2.15),SO与Ta和Rn均呈极显著正相关(P<0.01,df=48,F=17.20;P<0.01,df=48,F=14.81),同样说明Rn是影响常绿树种胀果树参气孔形态变化的主要环境因子,同时其还受Ta的影响[25-26]。而其FS与VPD呈显著正相关(P<0.05,df=48,F=16.73),与Rn呈极显著正相关(P<0.01,df=48,F=42.03)。另外落叶树种瓜木的SC与FS呈显著正相关(P<0.05,df=48,F=6.12),SO与FS呈显著正相关(P<0.05,df=48,F=9.25),常绿树种胀果树参的SC与FS呈显著正相关(P<0.05,df=48,F=11.06),SO与FS呈极显著正相关(P<0.01,df=48,F=29.41),结合两树种的蒸腾差异以及环境因子对它们气孔形态的影响不同,说明即使生长在相似环境下,当环境条件发生改变时,两树种气孔形态的反应不同,常绿树种胀果树参气孔形态特征(SD、SC、SO)对环境因子的响应比落叶树种瓜木气孔形态特征(SD、SC、SO)对环境因子的响应更强。所以导致两树种蒸腾差异的原因,一方面是两树种叶片的气孔密度自身存在较大差异;另一方面是叶片气孔形态对环境因子的反应敏感性不同。
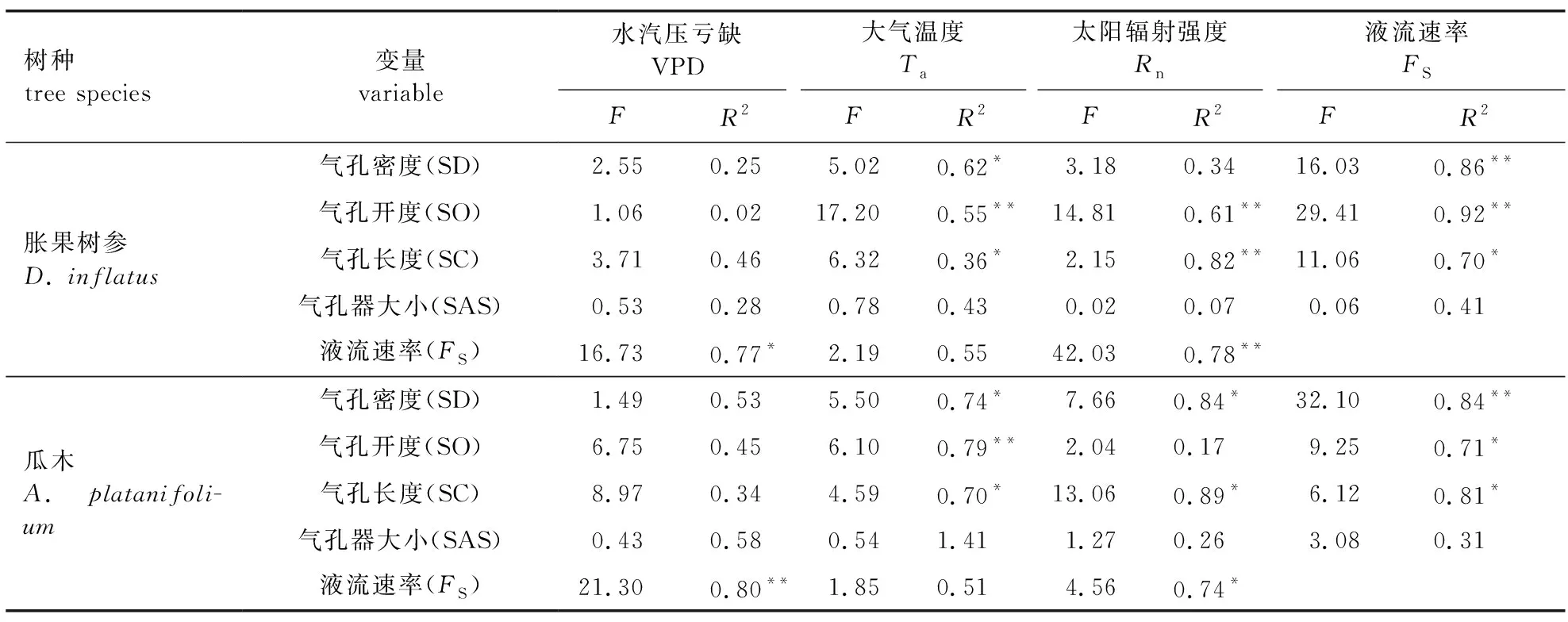
表2 胀果树参和瓜木叶片的气孔形态和树干液流速率与环境因子的相关性分析Table 2 Correlation analysis among stomatal morphology and stem sap flow rate with environmental factors of different leaves of evergreen D. inflatus and deciduous A. platanifolium
3 讨 论
植被在生长过程中与周围的水文环境相互影响[27],其蒸腾速率大于对水分的吸收能力,长时间条件下会导致植物不能完成正常生理活动而死亡[28],因此植物会做出一系列调节策略降低水分亏缺可能产生的损害[29],其中通过调节植物叶片气孔的开闭或气孔密度,对环境信号如光、温度、湿度等作出反应,以优化植物气体交换,避免干旱胁迫[7,21]。本研究发现,在喀斯特生境生长季(5月)5个连续晴天的条件下,两树种蒸腾表现不同,从叶片气孔形态特征上看,主要存在两个方面:
1)与自身叶片气孔形态变化有关。一方面表现在两树种自身气孔密度(SD)差异上,植物蒸腾是通过气孔导度、开度、密度的变化调节对水分的吸收来控制的[8-9],同时有研究发现,植物叶片气孔密度与植物蒸腾速率呈显著正相关[29-30],气孔密度越大,蒸腾速率越高,而植物气孔最大导度是由气孔密度的长期变化来调节的,这些气孔密度的变化可以表示为单位面积的气孔密度[31-32];同时研究发现,气孔密度的数量与自身遗传因素有关,即其最大值自身已经限定[33]。所以,研究结果表现出气孔密度越大的落叶瓜木蒸腾速率越高。另外,Brodribb等[34-35]研究叶片脱水过程发现,植物具有比较高的气孔密度和相对较小的气孔(气孔长度较短)时更适合生长在水分亏缺条件下;罗春梅[36]研究小麦叶片气孔分化及密度时发现,小麦气孔总数在春季多雨的年份最少,说明生活在湿润环境下植物叶片的气孔密度较低。因此,气孔密度越高的植物抗旱能力越强,相比而言,落叶树种瓜木更适合生长在裸岩率较高,土壤稀少异质性较高,土层薄且不连续,地下水文过程复杂的喀斯特生境[37]。
2)两树种叶片气孔形态对环境因子反应的敏感性不同。研究发现两树种的蒸腾与环境条件(光照、湿度)有关[20];结果还显示,太阳辐射强度是影响两树种气孔形态变化的主要环境因子,但对两树种气孔形态的影响不同,而常绿树种胀果树参的气孔形态变化还受大气温度影响。Driesen等[38]综述了环境因子(光照、湿度、温度、水状态)对植物叶片气孔开放的影响其表现因物种而异,但都会对气孔的开放产生影响。如Salvucci等[39]研究高温对气孔的影响发现,激活酶对热敏感,而这种酶在植物光合作用中起着重要作用,所以高温对光合作用产生了负面影响,又因植物光合作用与蒸腾同在气孔内进行,它们势必会相互影响;Thomas等[40]研究辐照度信号对烟草(Nicotianatabaccum)发育成熟叶片的影响,发现气孔长度和气孔密度均受到影响;Aliniaeifard等[41]和Fanourakis等[42]]研究高水汽压亏缺对植物气孔开放结果显示,在高水汽压亏缺条件下生长的植物,植物气孔变小(气孔长度变短),气孔将关闭,以响应较高的水汽压亏缺或较低的相对湿度,从而使植物能够在干燥的大气中控制水分流失。本研究结果说明,在连续晴天的茂兰喀斯特生境下,两树种的蒸腾强弱表现不同,与自身参与生理活动的叶片气孔的形态变化有关,主要表现在两树种自身气孔密度差异以及气孔形态对环境因子反应的敏感性不同,从而导致树种产生蒸腾差异。因此,类似叶片气孔密度高的物种更适合在喀斯特生境下生长。
