昆明地区引种荷花遗传多样性分析
2021-09-27赵琳林森赵琬玥胡慧贞陈龙清
赵琳,林森,赵琬玥,胡慧贞,陈龙清
西南林业大学园林园艺学院/国家林业和草原局西南风景园林工程技术研究中心/云南省功能性花卉资源及产业化技术工程研究中心,昆明 650224
荷花(NelumbonuciferaGaertn.)为莲科(Nelumbonaceae)莲属(Nelumbo)的多年水生草本植物,是中国十大传统名花中唯一的水生花卉,也是重要的切花植物[1],能有效吸收水体中的氮和磷,净化水质[2]。荷花已有2 500多年的栽培历史,正式文献记载的名称全球已达2 080个[3],根据用途划分为花莲、籽莲及藕莲,是集观赏、食用和药用价值于一身的重要的经济植物。
物种适生性及遗传多样性研究有助于探究物种对环境变化的响应,采取人为手段帮助物种适应环境变化,对于保护和利用遗传资源以及改良植物性状具有重要意义[4]。荷花为异花授粉植物,高度杂合,遗传变异多样,可供选择的亲本数量庞大,通过分子标记技术手段能高效且精确地反映个体分子水平上的遗传差异优势,目前常用的分子标记有InDel、SSR、SRAP、Scot等。其中InDel和SSR作为共显性标记广泛分布于基因组中,数量众多、突变率高、杂合程度高、多态性含量较高,在植物遗传多样性分析中应用广泛。Indel标记是不同个体基因组同一位点序列核苷酸片段的插入和缺失,其检出效率较高[5]。SSR标记为简单重复序列,按其来源可分为基因组SSR和EST-SSR,广布于整个基因组中,在荷花的进化及多样性分析中得到广泛应用[6]。此外,SSR标记及SNP标记在鉴别品种中具有较高的灵敏度,被国际植物品种权保护联盟(International Union for the Protection of New Varieties of Plants,UPOV)确定为构建植物品种DNA数据库的首选标记[7]。
2013年中国古代莲全基因组测序完成[8];2018年 “Chinese Tai-Zi”染色体水平上的基因组组装完成[9];这2个基因组测序的完成为荷花功能组学研究和新基因的挖掘提供了丰富的遗传信息,为解析不同生物学性状提供了基础。Hu等[10]对58个荷花种质资源进行遗传多样性分析,藕莲和籽莲分别聚为不同类群,花莲在2个类群中均有分布。Yang等[11]利用36对SSR标记对来自不同地区的11个居群的83份荷花资源进行遗传多样性和群体结构分析,发现泰国莲与中国莲居群之间的亲缘关系比美洲黄莲近。Yang等[12]结合分子标记对210份荷花种质资源进行连续2 a的表型观测,对荷花重要观赏性状遗传定位进行了研究。AN等[13]对18个省的94份藕莲进行了遗传多样性分析,发现西南地区和华东地区的藕莲品种具有更高的遗传多样性。Zheng等[14]通过EST-SSR标记构建包括荷花在内的3类水生植物指纹图谱,为快速鉴定后代种子是否纯合提供了便利。
云南地区高原湖泊众多,地理和气候环境多样,植物多样性丰富。普者黑、鹤庆等地素有栽培荷花的传统,近期有研究指出,云南热带地区可能是野生荷花及重瓣型荷花的原产地[15]。云南乡土荷花品种 ‘普者黑红荷’‘普者黑白荷’‘小洒锦’‘宜良千瓣’ 等皆具有较高的食用价值及观赏价值[16],但目前云南地区栽种的荷花多为外地引进品种,品种混杂,缺乏系统研究。为明确昆明地区引种栽培的荷花种质资源的遗传背景,更好地发挥该地区的生态环境优势,本研究利用InDel、EST-SSR和SSR标记对94份在昆明地区收集的引种荷花种质进行遗传多样性分析,充分了解引种栽培荷花遗传多样性、明确遗传背景,以期为云南地区推广荷花栽培提供参考依据并为荷花育种提供优良亲本。
1 材料与方法
1.1 试验材料
94份荷花种质原产于重庆大足、广西贵港、湖北武汉及四川遂宁等地,其中52份引种后种植于西南林业大学荷花种质资源圃,其余试验样品在云南昆明、曲靖、西双版纳地区采集所得,其中有30份为实生选育品种。品种信息详见表1。
1.2 材料种植及管理
2018年4月上旬,选取顶芽保存完好、藕身具膨大的藕节2节及以上的种藕,种植于口径 65 cm×高 70 cm 的黑色塑料缸。每缸种植2支,每一品种种植5缸,采用“藏头露尾”的方式栽植,根据植株生长状况适时进行科学管理。
1.3 DNA提取及标记扩增
选取无病虫害刚出水面的卷叶,采用天根多糖多酚专用植物基因组DNA提取试剂盒(DP305-02)提取DNA;经琼脂糖凝胶电泳检测质量,Nanodrop 2000测各样本质量浓度并记录,将各样本稀释至50 ng/μL,于 -20 ℃保存备用。
50对InDel引物为课题组自主开发,50对EST-SSR和50对SSR引物来源于已发表的文献[6,11,17-18],均由上海生工生物股份有限公司合成。随机选择5份供试材料DNA进行扩增,每个标记筛选出各20对多态性好、条带清晰的引物用于94份种质荷花的遗传多样性研究。采用6% 的聚丙烯酰胺凝胶对 PCR 扩增产物进行电泳、银染、显影、人工读带。
1.4 数据处理
利用Excel对分子标记扩增条带进行统计整理。利用POPGEN 32及PIC_CALC进行遗传多样性指标分析,利用Excel 2007及NTSYS-pc 2.10e进行遗传相似系数分析及Mantel检测。使用MEGA-X采用UPGMA法进行聚类,iTOL在线工具进行美化。利用Structure 2.3.4进行群体结构分析,使用Structure Harvest在线工具计算最佳K值,确定群体数量。
2 结果与分析
2.1 InDel、EST-SSR、SSR遗传多样性分析
分别选取InDel、SSR、EST-SSR各50对引物进行扩增,筛选出InDel、SSR和EST-SSR各20对能扩增出丰富、稳定、清晰条带的标记对引种的94份荷花种质进行分析,发现筛选出的标记能较好地揭示94份荷花材料多样性及遗传差异(表2)。3种分子标记共扩增204条条带,总共检测到233个等位基因。每种标记都表现出较高多样性水平,平均Shannon指数≥0.85,基因多样性指数≥0.52,多态性信息含量≥0.44,其中SSR标记在94份荷花种质中表现出更高的多态性水平。
2.2 InDel、EST-SSR和SSR标记相关性检测
对3种标记进行Mantel检测,相关性分析结果(表3)显示,InDel、EST-SSR和SSR三者间相关性除EST-SSR和SSR≥0.529 0,其他均小于0.5,说明3种标记间的相关性较差,但3种标记组合与InDel、EST-SSR和SSR遗传相似系数矩阵相关性系数均大于0.789 4,呈高度正相关,表明可以采用3种标记组合进行遗传多样性分析。
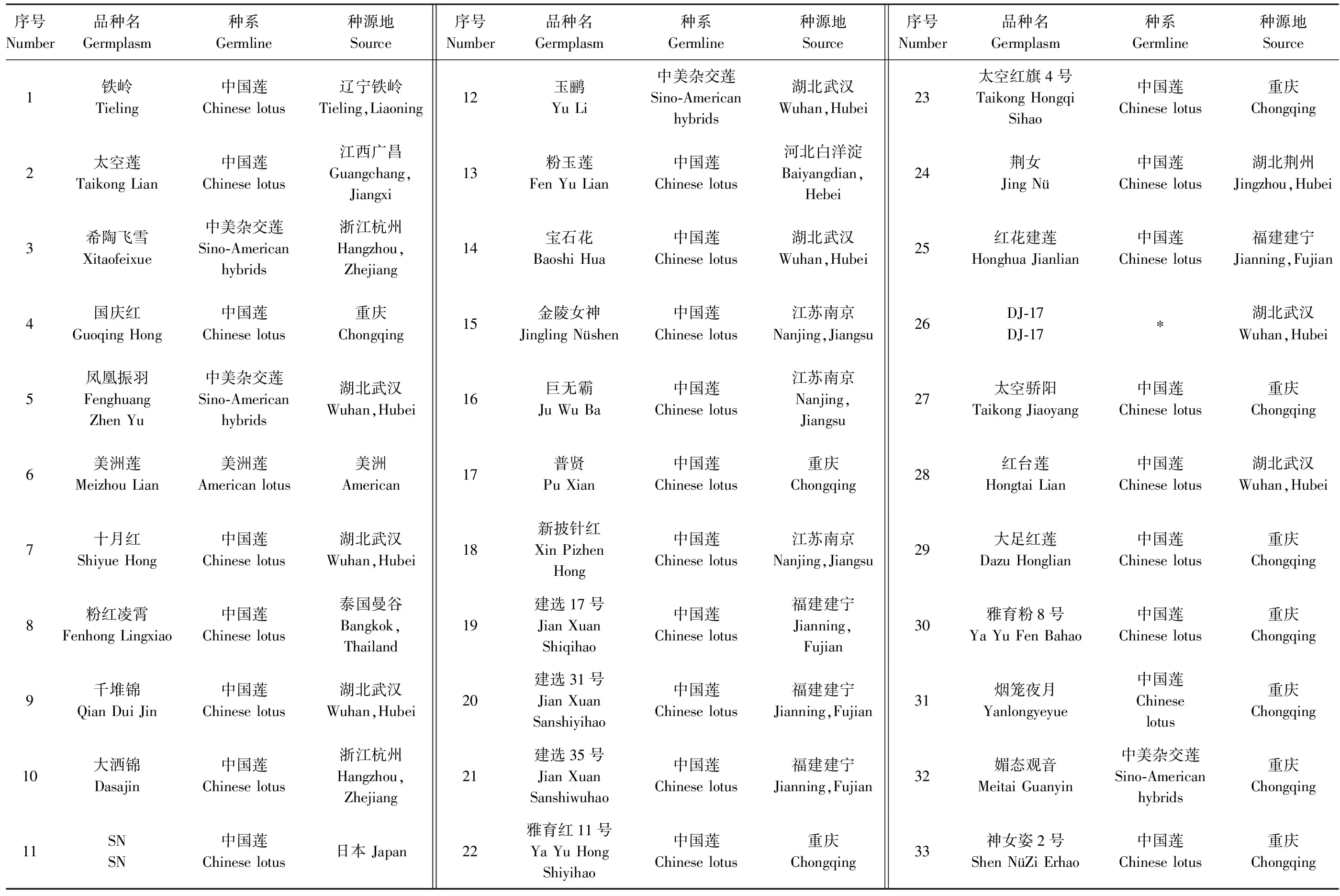
表1 供试种质及其来源 Table 1 Ninety-four Lotus accessions used in this study and their origins
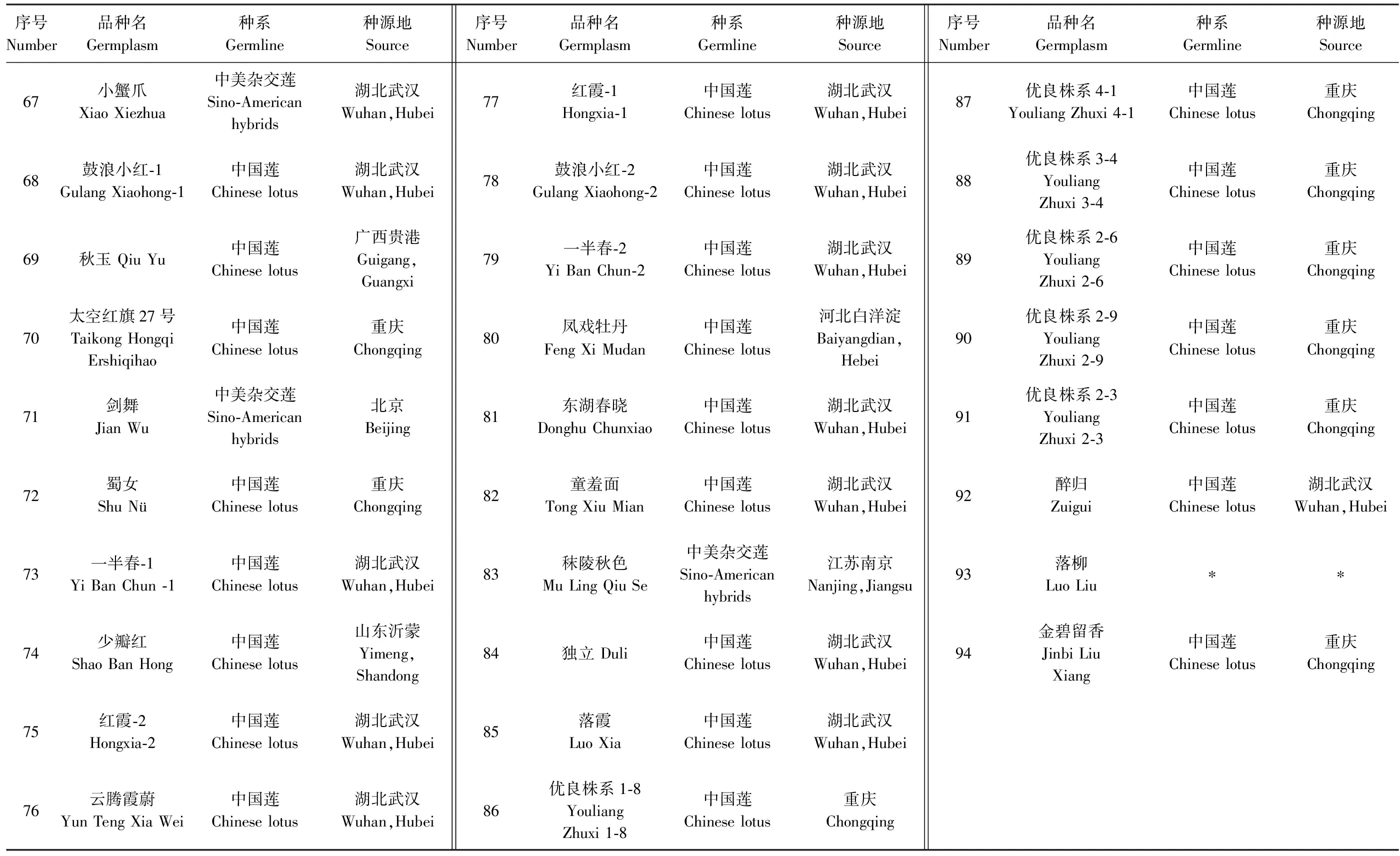
续表1 Continued Table 1

注:1)带“∗”表示该材料信息不详; 2)‘优良株系X-X’为自主培育的新品种; ‘秋日红花’ ‘秋星’ ‘秋艳’‘秋阳’‘秋玉’为秋荷品种,为中国莲与泰国莲杂交的后代。Note:1) Materials with ‘ ∗ ’mean unknown information; 2)‘ Youliang Zhuxi ’is new varietiy cultivated independently; ‘ Qiuri Honghua ’ ‘ Qiuri Xing ’ ‘ Qiu Yan ’ ‘ Qiu Yang ’ ‘ Qiu Yu ’ are autumn lotus varieties,the off-springs of crosses between Chinese lotus and Thai lotus.
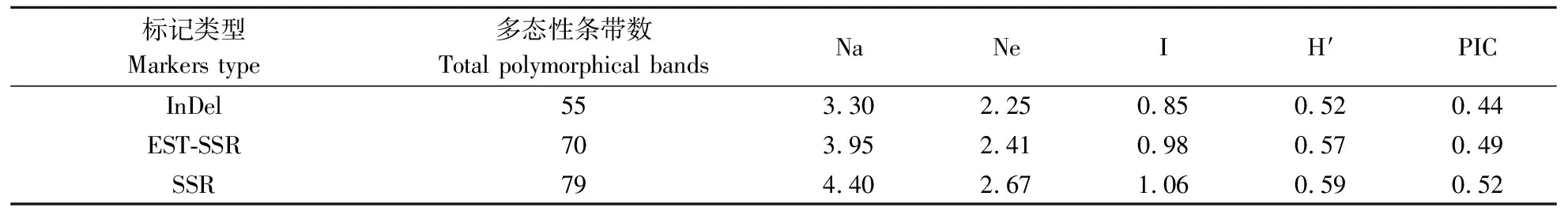
表2 InDel、EST-SSR、SSR引物多态性分析 Table 2 The polymorphism analysis of InDel,EST-SSR and SSR
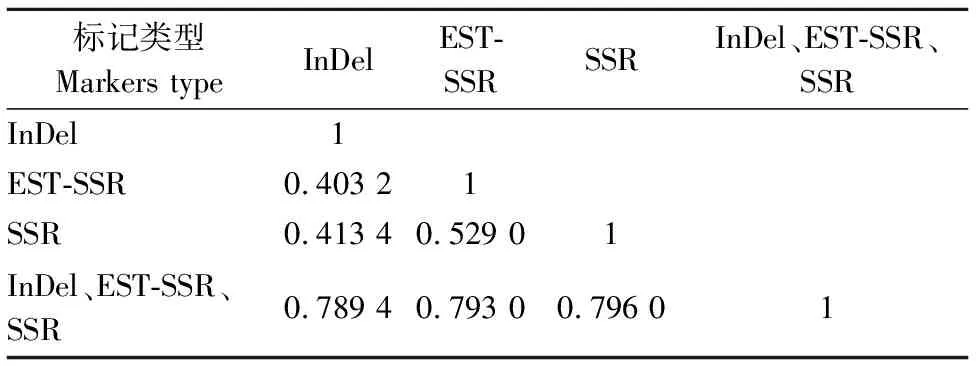
表3 InDel、EST-SSR、SSR标记遗传相似系数相关系数 Table 3 Correlation coefficients of InDel,EST-SSR,SSR genetic similarity coefficients
2.3 基于InDel、EST-SSR和SSR的聚类分析
聚类分析将94份种质资源划分为5大类(图1)。结合种源地信息、形态特征和聚类结果综合分析,结果表明:可以将类群 Ⅰ 细分为6个亚群。类群 Ⅰ-Ⅰ 有 33份材料,种源地为重庆及武汉的材料居多,除‘金凤凰’‘如意观音’为中美杂交莲,31份为中国莲,4份为实生苗选育品种,均为单瓣品种,其余29份荷花材料为重瓣荷花;除‘金凤凰’为黄色,‘水中月’为白色以及‘大洒锦’为洒锦色系以外,其余均为紫堇色或淡紫堇色系。类群 Ⅰ-Ⅱ 有8份材料,仅‘玉鹂’为中美杂交莲,其余均为湖北武汉地区的中国莲,除‘艳阳天’外均表现为中小株型重瓣荷花,类群 Ⅰ-Ⅲ 有7份材料,仅‘锦红袍’为中美杂交莲,其余均为中国莲品种,2个为秋荷品种,3个品种为太空育种品种,‘千瓣莲’被划分在此类群内,花色多为紫堇色。类群 Ⅰ-Ⅳ 有17份材料,均为中国莲品种,无明显的种源地划分;花色均为紫堇色或淡紫堇色系,大株型品种较多,3份秋荷品种聚类到一起,单瓣大株型籽莲均在此类群内,遗传距离较近。Ⅰ-Ⅴ 仅‘新披针红’一份材料,引自江苏南京,为中小型重瓣荷花。Ⅰ-Ⅵ 类群有19份材料,有4份为美洲莲品种,以湖北武汉地区引种品种较多,多为当年莲子播种的中小株型荷花。
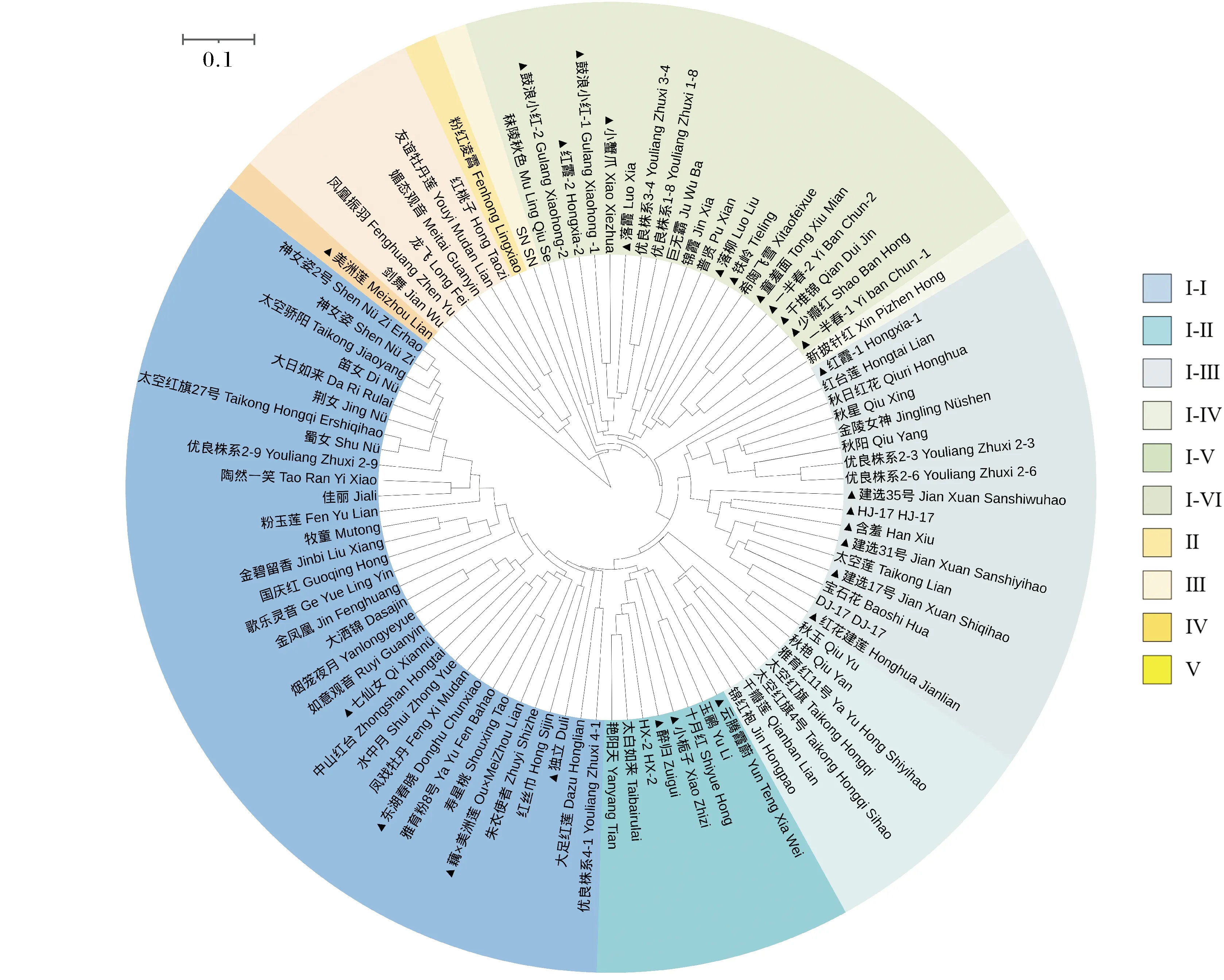
带▲为实生苗栽培品种 The cultivars with ▲ are indicated seeding culticars.图1 94份荷花种质资源聚类分析Fig.1 Cluster analysis of 94 lotus
类群 Ⅱ,‘SN’独聚为一支,该品种来自日本,与其他荷花品种地理距离较远。
类群 Ⅲ,‘粉红凌霄’独聚为一支,为热带莲品种。
类群 Ⅳ 有6个材料,除‘红桃子’无法确认其种源外,其中5个均为中美杂交莲,花色除‘媚态观音’及‘红桃子’为复色品种外,其余以黄色和白色为主。
类群Ⅴ,‘美洲黄莲’独聚为一支。
2.4 群体结构分析
为了明确不同种质间的群体结构关系及遗传背景差异,对94份材料的群体遗传结构进行分析,从图2可以看出,94份荷花材料在K=5时,ΔK出现峰值,说明在此群体中有5种主要的基因组结构成分。
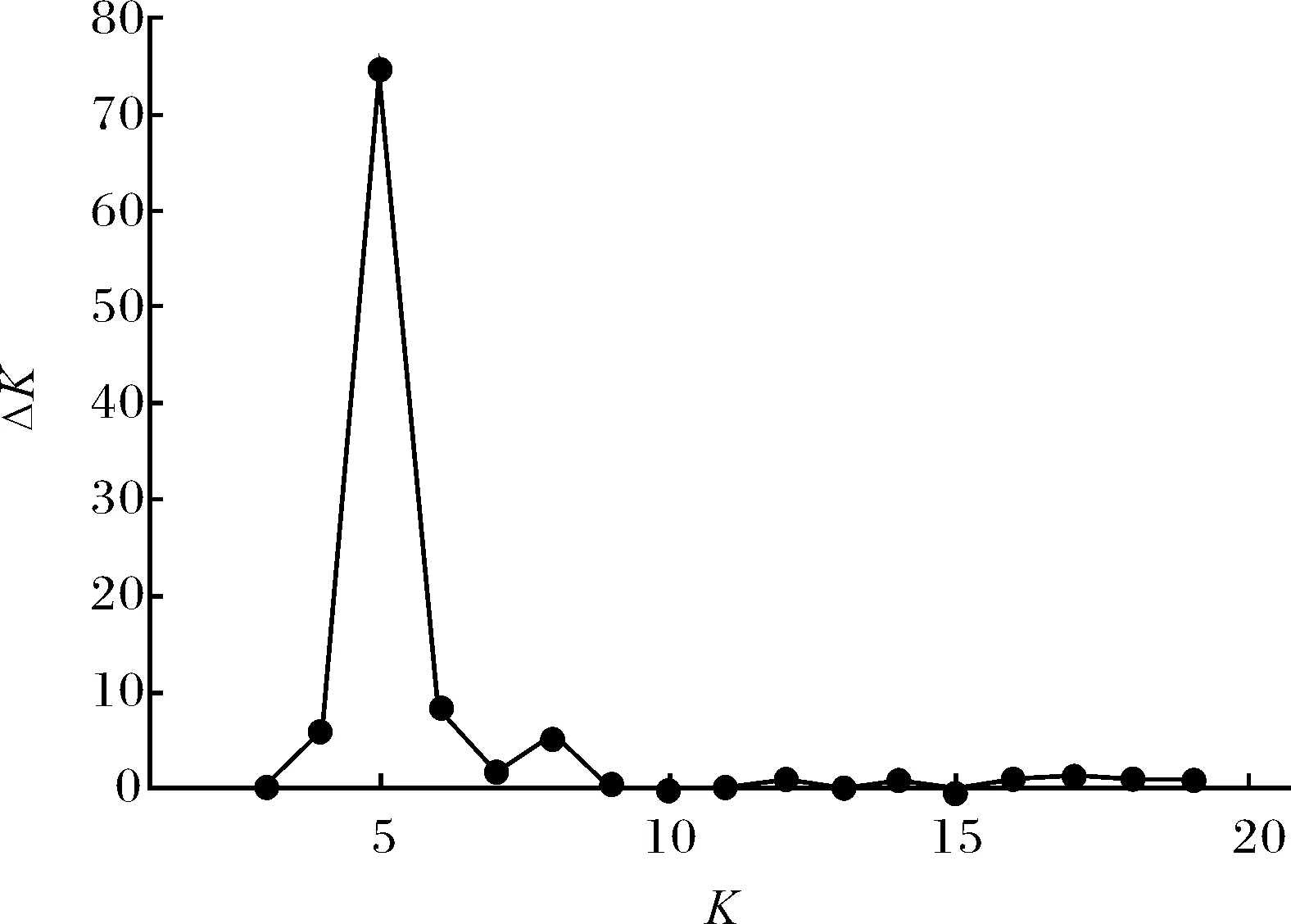
图2 ΔK随K变化的折线图Fig.2 ΔK values changing with K value
群体结构分析的分组结果(图3)显示,根据颜色的不同可将94份荷花的主要遗传组分划分为5种;根据遗传组分的相似性可以将94份种质聚类成5个亚群,各亚群所包含的种质材料与聚类分析的结果有相似之处。
遗传组分的分组结果显示群体R与聚类分析中的 Ⅰ-Ⅳ 种质资源较一致,除‘媚态观音’外,其余均为中国莲,种系不明的‘红桃子’可能为中美杂交莲;大部分为子莲及实生苗选育品种。群体G中与聚类分析中的类群 Ⅰ-Ⅰ 中的大部分种质一致,大都为中国莲品种,其中仅1份为黄色系中美杂交莲‘金凤凰’,但其遗传组分与美洲黄莲相似度较低。群体B中除‘水中月’为白色外,均为堇紫色品种。‘锦红袍’‘陶然一笑’‘如意观音’与美洲莲遗传组分的相似度很低,所以聚类结果中类群 Ⅰ-Ⅰ 及类群 Ⅰ-Ⅲ 均为中国莲遗传组分占比高的种质资源。群体Y主要分布在类群 Ⅳ、Ⅴ 中,均为美洲莲及中美杂交莲。群体P与类群 Ⅰ-Ⅴ 中的种质较一致,以武汉地区中国莲为主,大部分为中小型荷花品种实生苗;其中‘玉鹂’与美洲莲相似的组分很低,所以与中国莲聚类在同一个组中;来自日本的‘SN’也在此群体内,同一品种的不同实生苗遗传组分也有所差异,且其遗传组分组成较复杂。

图中数字为种质材料编号。The accession numbers are indicated in the figure.图3 群体结构Q图Fig.3 Q-plot of the population structure
3 讨 论
3.1 组合分子标记对遗传多样性的检出率高
本研究中的94份荷花种质来源广泛,遗传背景复杂,观赏性状及农艺性状丰富。不同的分子标记其来源不同,所揭示的基因位点的遗传信息有所差异,可以为遗传多样性研究及种质资源分类提供更丰富的遗传信息,本研究通过Mantel检测发现组合标记与单个分子标记的r值均大于0.75,采用组合分子标记发现供试群体具有丰富的遗传多样性。Hu等[10]利用AFLP和SSR标记进行遗传多样性分析,发现AFLP、SSR联合分析的遗传相似系数分别与AFLP和SSR标记高度匹配,使用联合标记可以更好地进行聚类。Yang等[19]使用SSR、SRAP对43份荷花材料进行基因分型,SSR-SRAP联合分析分别与SSR、SRAP高度相关。本研究的群体结构分析显示,同一品种的实生苗在遗传组分上存在一定差异,说明联合标记检出率更高,在一定程度上能揭示较多的遗传信息。
3.2 形态和分子标记相结合更利于对引种荷花的评价
结合3种分子标记的结果进行聚类分析,发现荷花种质在分子水平上与形态水平上对资源的分类并不完全一致,但具有一定的关联性,袁王俊等[20]在菊花的遗传多样性分析中也发现,传统的分类方法不能完全反映菊花的亲缘关系,说明分子水平上揭示的遗传信息差异不一定能在形态上表现出来;在关联分析的研究中发现,分子标记位点间的相互连锁会导致同一个分子标记位点同时影响多个相互关联的表型性状[21]。传统的形态学特征与荷花的经济价值直接相关,将形态学标记与分子标记结合分析对育种实践具有重要的指导意义。聚类的群体与种质资源的种源地没有明显的关联,这与尹静静等[22]和李青竹等[23]对种藕的研究结果类似。Yeshitila等[24]利用9对SSR标记对来自不同自然地理分布的15个群体共计216个热带莲样本进行遗传多样性分析,发现遗传距离与地理距离存在正相关关系,但是Islam等[25]利用12对SSR引物对326个美洲莲的遗传多样性分析,发现遗传距离与地理距离无显著相关性。在本研究中,‘SN’‘粉红凌霄’‘美洲黄莲’等种源地与中国莲种质资源明显分隔的品种,无论是在聚类还是群体结构分析中与中国莲品种均有差异,而群体内来自中国各地区的种质资源无明显划分,可能是由于引种的品种多为近期人工杂交选育,各地种质资源交流频繁导致基因交流频繁,基因组分较为复杂。籽莲品种均聚类在同一个群体或类群内,大部分的研究表明藕莲、籽莲在分子水平上有较大的差异,它们复杂的遗传关系可能是由于长期人工选择和无性繁殖逐渐形成的遗传差异[19,26]。本研究中花莲观赏性状丰富,且部分品种与籽莲遗传分化明显,可选择优良品种与结实较多的籽莲进行杂交,选育新品种。
在聚类及群体结构分析中发现,部分有记载种系来源的荷花品种与聚类结果有差异,可能是由于标记数量较少导致,同时聚类及遗传结构分析也能帮助重新注释品种的背景,为后续的研究提供科学的指导。
聚类结果以及群体结构分析皆显示,美洲莲被独聚为一支,与亚洲莲遗传距离较远,与前人研究结果一致[15]。中美杂交莲在聚类的过程中没有完全聚在一起,这可能是由于不同品种的中美杂交莲遗传组分构成不同,通过群体结构分析也可看出,部分中美杂交莲遗传组分与美洲莲相似度极低,与中国莲相似度更高。秋荷品种也并未完全聚类到一起,除秋玉外,其余品种遗传组分类似,与种源地为泰国的‘粉红凌霄’遗传组分部分相似。
综上,InDel、EST-SSR和SSR 3种分子标记组合对荷花多样性研究有较高的检出率,94份荷花品种遗传多样性高,亚洲莲及美洲莲有明显遗传分化,不同品种的中美杂交莲或秋荷在遗传组分存在差异,研究结果可为后续的优良品种选育及选配适合的杂交组合提供种质资源及技术支撑。
