2010−2020年草地土壤氮循环研究现状与发展趋势
2021-09-27孙盛楠严学兵
赵 仪,孙盛楠,2,严学兵
(1.扬州大学动物科学与技术学院,江苏扬州 225009;2.省部共建三江源生态与高原农牧业国家重点实验室,青海西宁 810016)
草地约占全球陆地总面积的25%,氮循环是草地生态系统最基本的循环过程,其4个主要过程(生物固氮作用、矿化作用、硝化作用和反硝化作用)均由微生物驱动。生物固氮作用将大气中的分子态氮在微生物体内由固氮酶催化还原成氨,是土壤氮素的重要来源。矿化是指土壤中有机态氮向无机态氮转化的过程,分为氨基化阶段和氨化阶段[1]。硝化作用作为土壤氮素循环的中心环节,连接固氮作用和反硝化作用,不仅决定着土壤中氮素有效性,还与硝酸盐淋失造成的水体污染密切相关[2],硝化作用必须在通气条件下分两个阶段完成,即氨氧化过程和亚硝酸氧化过程,为反硝化作用提供反应底物。在草地农业生态系统中,硝化作用和反硝化作用经常耦合发生,是氮肥损失、温室气体生成的主要途径。
土壤氮循环过程需要多种生物参与,受到多种因素影响。草地土壤中氮的利用取决于其在土壤中的存在形式,氮的存在形式受氮转化过程调控,尤其是矿化、硝化以及无机氮的固定[3]。有机碳为矿化提供能量来源,研究表明,总氮矿化速率和净氮矿化速率随有机碳含量的上升、氮浓度的增大而提高,但却与碳氮比成反比[4]。这可能因为,高碳氮比的有机质通常与土壤有机质中高浓度的顽固化合物有关,从而对氮矿化产生不利影响。矿化形成的铵态氮去路较多,一方面被植物直接吸收利用,通过硝化作用转化为硝态氮再利用,被土壤颗粒吸附;另一方面通过微生物的固定形成铵,以氨气的形式散失到大气中,通过淋溶损失或流入水体[3]。硝化过程是氮循环中不可或缺的一个环节,但对草地生态系统往往造成不利影响,硝酸根离子在土壤中移动性较强,易淋溶,容易降低肥料利用率,引起水体富营养化等环境问题[5]。土壤pH 是调节硝化过程的最重要的因素,研究表明,土壤pH<5时,硝化作用受到抑制[6]。同时,硝化作用也有可能造成土壤酸化(土壤吸附性复合体接受一定数量的氢离子或铝离子,使土壤中碱性离子淋失、pH 降低)。硝化过程形成的硝酸根离子除了通过淋溶、反硝化途径损失外,有小部分被微生物固定。微生物生物量氮库与其他氮库相比微不足道,但其快速周转能在植物需氮时为植物提供有效氮[7]。氮素的矿化、固定以及硝化是草地生态系统中调节氮素有效性最重要的几个过程,尤其是土壤中氮素矿化–固定周转速率,对氮素有效性起至关重要的调控作用[5]。
草地可利用氮素的供应和草地的生产力密切相关,氮素供应是草地生产力高低的限制因子。研究草地土壤氮循环对于提高草地生产力至关重要。国家在顶层设计和统筹谋划中,充分重视生态系统养分循环过程的研究,启动实施了包括战略性先导科技专项“青藏高原多圈层相互作用及其资源环境效应”、“土壤–微生物系统功能及其调控”和国家重点研发计划重点专项“典型脆弱生态修复与保护研究”、“全球变化及应对”等项目,其中生态系统氮循环是各项目的研究热点与核心。目前,国内外学者对草地土壤氮循环机制、影响氮循环的因素、氮循环对全球气候变化的响应以及碳氮循环耦合机制等做了大量研究。本文利用Citespace 软件对2010 − 2020年间相关文献进行可视化分析,确定草地土壤氮循环的研究主体、研究热点,分析草地土壤氮循环研究的发展趋势,绘制草地土壤氮循环研究图谱,为今后研究草地生态系统氮循环提供思路。
1 数据来源和研究方法
1.1 数据来源
本文数据采集于Web of Science(WOS)核心合集数据库,在数据库里面进行高级检索,将主题设置为(grassland OR steppe OR meadow OR pasture OR rangeland)AND (nitrogen cycle OR cycle of nitrogen OR nitrogen cycling),时间跨度设置为2010 − 2020年,剔除重复的、没有作者的文献、会议论文、编辑材料、书评等,最终得到2 455篇有效文献。
1.2 研究方法
本研究使用Citespace软件对2010− 2020年间草地土壤氮循环方面的文献进行计量分析[8]。以一年为时间分割点,分别以作者、机构、国家、关键词以及共被引文献作为节点,对相关文献的关键词以及共被引文献等进行知识图谱的绘制,以展示该领域的研究热点和发展前沿。其中,文章中心性(测度节点在网络中重要性的一个指标)由Citespace可视化界面nodes→compute node centrality 直接计算而出。
2 结果与分析
2.1 草地土壤氮循环相关研究发文量年度分析
2010− 2020年间草地土壤氮循环方面的发文量呈上升趋势(图1)。其中,2010−2012年发展趋势趋缓;2020年增长幅度最大,一年间发文量增加63篇。从2014年起,草地土壤氮循环方面的文献涨幅逐步稳定,进入快速而稳定的发展阶段,这表明草地土壤氮循环已经成为一个研究热点。截止2020年,WOS核心合集数据库收录草地土壤氮循环文献共计2 455篇。我国草地氮循环领域的研究也在2011年进入迅速发展阶段,发表的文献比例逐年上升,2017年起比例就超过了40%,尤其在2018年达到峰值,发文比例高达50%。
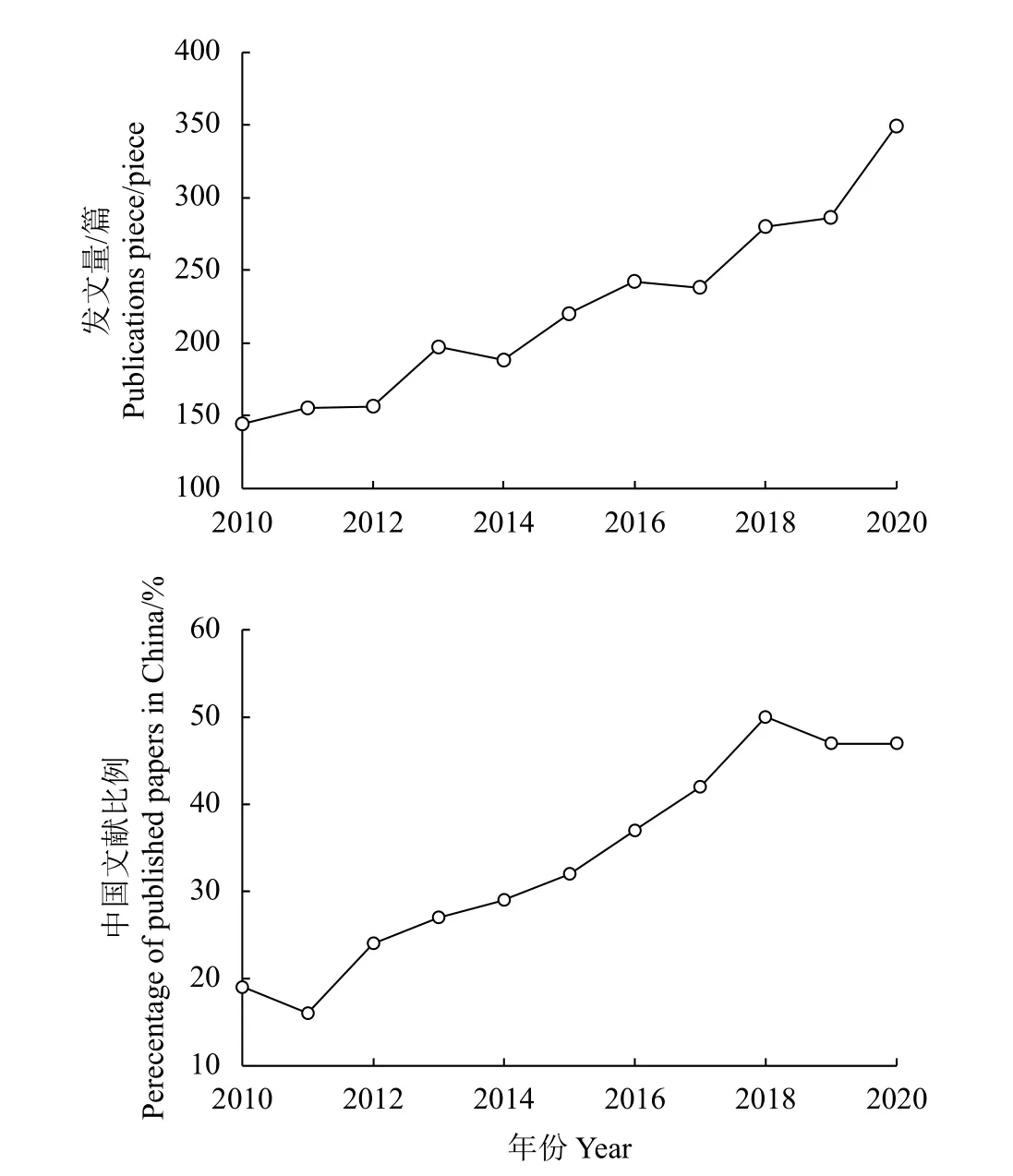
图1 草地土壤氮循环年度发文量统计及中国发文量占总发文量的比例Figure 1 Annual number of published studies of soil nitrogen cycle in grasslands(top panel)and the proportion of publications from China (bottom panel)
2.2 研究主体分析
发文量前五的国家依次为中国、美国、德国、澳大利亚和巴西,中国2010 − 2020年共发文895篇(表1)。中国科学院的研究团队是推动草地土壤氮循环领域发展的中坚力量,不仅发文多,文章中心性也位居第一,高达0.22(表1);但中国的文章总体中心性却只有0.01,表明国内各机构应加强交流合作以提高文章质量。其他国家发文量虽不及中国,但文章总体中心性都不低于中国,尤其是德国中心性较高。从机构发文中心性看,德国和中国并列第一,均为0.22(表1)。此外,对发文作者进行分析,可以发现韩兴国的发文量和文章中心性都稳居第一。
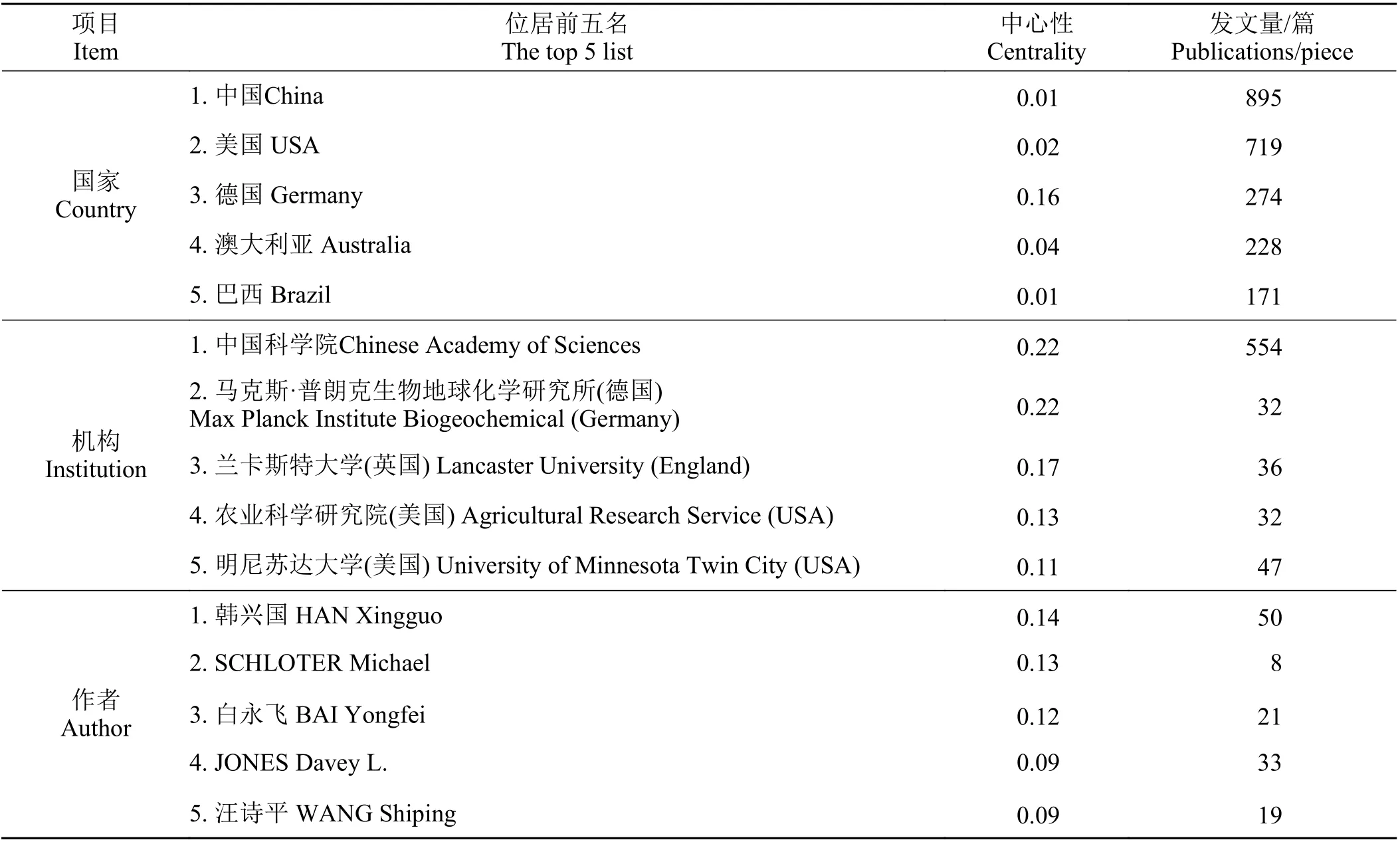
表1 WOS核心合集数据库中研究主体分析Table1 Analysis of research subjectsin the WOScore collection database
2.3 草地土壤氮循环基础研究分析
2010−2020 年草地土壤氮循环前10篇被引文献中,被引次数最多的是研究有机质的文献(表2)。有机质富含碳,其含碳量是大气或陆地植被的3倍多,部分有机质稳定性高能保存上千年,但有的却极易分解;有机质的分解与否往往取决于土壤中微生物和所处环境[9]。例如,Batjes[10]的研究指出,全球变暖能加速有机质分解,释放CO2到大气中,进一步加剧变暖趋势。土壤温度升高加速微生物的生长,促进有机质的分解[11]。被引次数仅次于有机质的是氮沉降方面的文献。农业集约化提高了氮肥使用频率,氮肥提高初级生产力,促进退化草地恢复,然而氮沉降却因此日益加剧。Bai 等[12]对内蒙古草原同一类型的成熟草地和退化草地进行氮添加处理,结果发现氮沉降导致草原生物多样性丧失,降低生态系统的恢复力。Bobbink 等[13]得出类似结论:氮积累推动物种间竞争,是物种组成变化的主要驱动因素。全球气候变化直接影响生态系统氮循环,也通过影响植物对氮的吸收、土壤微生物组成以及植物与微生物间的相互作用来间接改变生态系统对氮素负荷量的响应;例如,大气CO2浓度升高可能促进植物生产力,导致渐进氮限制,这是因为CO2浓度升高引起净初级生产力提高,不仅导致生态系统中碳储存,还导致氮储存,从而降低氮的有效性[14]。氮限制在生物群落中广泛存在且受地理位置和气候影响,草地对氮限制的响应随纬度增加而增加,与降水和温度无关[15]。这表明,全球的碳、氮循环存在着强烈的相互作用,地理因素也可以介导生物群落对氮限制的响应。

表2 共被引网络图中被引频次排名前10的文献Table2 Referencesin thetop 10 cited frequency in theco-cited network diagram
2.4 关键词分析
图2展示了频数 > 100的关键词。氮/Nitrogen 以及草地/Grassland 作为主要搜索词,出现频率较高,分别为746和547,且两者间存在紧密联系。根据关键词,将草地土壤氮循环的影响因素分为生物因子、人类活动和非生物因子,生物因子包括植物与微生物,人类活动主要包括放牧、开垦、火烧、施肥与灌溉、刈割以及围封,非生物因子主要是大气氮沉降、增温与CO2浓度升高以及降水。
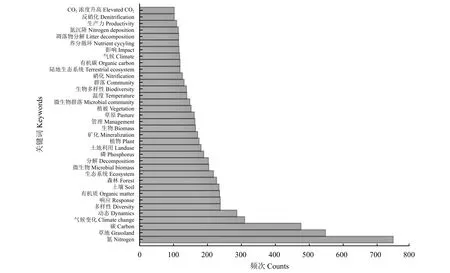
图2 2010–2020年关键词的频数Figure 2 Frequency of keywords in studies of grassland soil nitrogen cycle during 2010 – 2020
2.4.1 生物因子
陆地生态系统中地上、地下部分相互作用,传统研究对地上部分的关注多于地下部分,随着研究技术的进步,地下部分也得到了重视。植物与土壤微生物的相互作用是联系生态系统地上、地下部分的重要纽带。一方面,植物通过其凋落物和分泌物为土壤微生物的活动提供碳源。有学者对42项利用15N 标记研究中的424对数据进行分析,结果指出,根际分泌大量可利用碳,刺激了根际土壤微生物的生长且促进微生物的活性,从而提高土壤氮的有效性和植物的氮利用率[16]。另一方面,土壤微生物通过分解有机质促进养分循环以保证植物的正常生长发育。微生物能够调节土壤氮素的有效性,从而影响植物对氮素的吸收[17]。生长快、生命周期短且氮素获取速率快的植物往往在氮素有效性高的生态系统中占主导地位,但是这些植物的氮的利用效率较低,大多数的氮素损失在其衰老组织中,然后流向土壤;相反,生长慢、生命周期长的植物更能适应氮素有效性低的生态系统。氮素吸收速率快的植物有降低氮有效性的潜力,易导致土壤中的氮损失[18]。植物与微生物之间普遍存在互惠互作关系,以增加生态系统中的氮保留,减少土壤中的氮损失量;其中,最常见的就是共生固氮系统,尤其是豆科植物–根瘤菌体系,为生物固氮做出了巨大贡献。当生态系统处于氮限制时,植物可以通过共生和多营养级交互作用重组微生物群落来获取氮素[19]。丛枝菌根真菌是最古老、最广泛的共生真菌,该真菌能够吸收硝酸盐、铵和有机氮,并将他们转移到植物体内供植物吸收利用[19]。除了丛枝菌根真菌,外生菌根真菌、固氮细菌等也是常见共生微生物。细菌为主的微生物群落倾向于与生长快、凋落物质量高的植物建立共生关系,因为植物凋落物中的无机氮含量高,有利于细菌进行矿化作用,不过这样也容易通过反硝化和淋溶提高生态系统的氮损失量[20]。
在养分强烈受限的生态系统中,活根释放的根际分泌物为矿化微生物提供碳源,使得微生物群落扩张,需氮量上升,进而与植物在土壤中争夺养分,影响草地生态系统氮循环。此外,植物还进化出了直接干扰机制即通过释放次生代谢产物直接保护氮素,次生代谢产物不利于导致土壤氮素流失的微生物的生长[19]。这种抑制现象最先由Lata 等[21]在热带草原的研究证实,研究指出植物可以通过抑制因子来干扰硝化细菌的硝化能力以吸收更多的无机氮。Zhao等[22]在苜蓿(Medicago sativa)与鸭茅(Dactylis glomerata)间作的研究中也发现了类似现象,间作显著降低了植物根部氨氧化菌的丰度。目前,已有结果证实这些抑制因子的存在,植物根部通过分泌次级代谢产物,比如环二萜、3-(4-羟基苯基)丙酸等来阻断硝化作用,这种现象被称为生物硝化抑制[23-24]。植物与微生物间的负反馈调节机制的机理很多,但仍需要进一步研究以确定植物与微生物之间的竞争干扰策略,从而更深入地理解植物–微生物在土壤氮循环中的相互作用。
2.4.2 人类活动
放牧是对草地最直接的利用方式,放牧的强度、时间、制度以及动物的采食行为均会影响草地土壤氮循环,其中放牧强度是最重要的影响因子。研究表明,放牧加速草地养分循环,动物的踩踏增强土壤的紧实度,且随着放牧强度的增加,土壤中的总氮含量下降[25]。但并非强度越大,土壤中的氮储量就越低。Li等[26]研究发现,较高放牧强度反而提高高寒草甸生态系统氮储量,这可能因为高强度放牧改变了草地的物种组成和生物量分配格局。放牧强度越大,氮的矿化速率越高。放牧时,动物的排泄物经表层土壤中的微生物分解、动物踩踏凋落物混入土壤有机质中,提高了凋落物的分解速率和土壤微生物生物量,矿化作用得以增强[27];过度放牧导致草地的植被发生变化时,矿化作用受阻,可能是由于土壤孔隙度下降、通气透水不良、微生物活性受影响、土壤养分改变,氮损失超过了输入量[28]。近些年草地退化严重,有学者分别研究了利用强度对羊草(Aneurotepidimu chinense)草原和荒漠草原土壤氮循环的影响,得出一致结论:4种氮素转化细菌(自生固氮菌、氨化细菌、硝化细菌和反硝化细菌)的数量随放牧强度呈现倒“V”曲线.轻度放牧时,氮素转化细菌数量显著增加,有助于增强硝化作用、氨化作用,促进氮素循环;随着放牧强度提高,氮素转化细菌数量下降,抑制了土壤中的氮素循环过程[29-30]。放牧制度也时刻影响草地氮循环,例如轮牧,轮牧是指草场按照需要定期或间歇地放牧和休息[31]。研究表明,与连续放牧相比,轮牧下的植物均匀度和丰富度、土壤含水量、全氮以及全磷含量均有所增加[32]。放牧对草地土壤氮循环的影响因草地的类型以及放牧强度不同而有所差异,但众多研究表明放牧强度要在一个合理的范围,以保证草地的最大生产力和可持续发展[33-34]。
开垦是影响草地土壤氮循环的另一个重要人为因素。开垦改变了草地现有的土层结构、理化性质以及微生物构成。开垦破坏了草地的表层土壤,储存在表层土壤中的有机质大量分解、释放,土壤有机质含量下降,造成表层土壤的氮损失、水土流失。开垦后有机碳和全氮含量均存在下降的趋势,主要是因为土壤通气透水性得以改善,硝化作用增强,增加了草地中N2O、NO的排放量,并导致了硝态氮的流失[35]。开垦造成的土壤氮损失、水土流失以及全氮含量的下降,导致微生物可利用的碳、氮减少,微生物的生物量和活性下降,从而影响土壤中氮的矿化和固定。Roth 等[36]在德国Kleve 研究站采用机械翻耕、化学除草剂杀灭两种方法对比开垦方式对草地的影响,结果显示:两种方式都增大了N2O的排放通量,且机械翻耕法的排放量大于化学除草剂杀灭的排放量,排放量分别为未开垦草地的2.5、2 倍。开垦促使大量N2O排放和硝态氮的流失,加剧温室效应、地下水污染以及草原退化。修复退化草地往往需要耗费许多时间和劳力,应该根据草地类型、当地水热环境条件等因素,选择适宜的开垦方式,制定合理的草地管理制度,提高草地的蓄水保肥能力,促进草地生态系统的稳定发展。
火烧是一项比较古老的草地改良措施,能够清除地上植被,增加土壤的灰分含量,烧死危害草地的昆虫并刺激部分生物体再生,从而促使草地植被的更新。火烧主要通过改变土壤养分状况以及土壤温度来直接或间接地影响草地土壤氮循环[37]。研究表明,火烧显著提高半干旱草地土壤有机碳、全氮以及铵态氮含量,这可能是因为火烧通过清除地面作物和凋落物增加了土壤表层养分,进而影响草地生态系统的碳、氮循环过程[38]。草地火烧的效果因草地植被类型不同而产生差异。矮草草原、荒漠等植被稀疏、气候干旱且有机氮含量较低,不适宜火烧;高草草原植被覆盖度高,适宜的火烧频次、强度以及时间利于该草原的牧草生长和土壤微生物活动[39]。火烧后的高草草原地表温度升高,固定在凋落物和植株中的养分以无机态补充到土壤中,氮转化细菌的活动逐渐频繁,促进了硝化、矿化等过程,草地地上植物群落组成也因此改变[40-41]。火作为影响草原生态健康、稳定的因子,其造成的后果持续时间比较长。相关研究发现15年的火灾显著抑制了落叶松(Larix gmelinii)凋落物的后期分解,且固定化过程远超过释放,土壤pH 持续增长,抑制微生物的活动和相关酶的产生,从而阻碍氮循环过程[42-43]。有研究证实,火能够改变草原土壤理化性质以及养分循环的过程,调控草原生态系统的结构和功能,但是火灾的长期影响仍不是很清楚。在进行日常草原管理时,可以选择在降水充沛时节进行适宜强度的火烧,以促进草地植被更新和提高营养物含量,保证草地的可持续发展。
施肥和灌溉是改良草地土壤的重要措施,水和肥的吸收具有相关性,二者相互结合,保证草地生产力。施肥、灌溉主要通过土壤性状、微生物的改变来间接影响氮循环。魏金明等[44]研究了水分、氮肥和磷肥添加对内蒙古典型草原土壤碳、氮、磷含量及 pH 的影响,结果表明,水肥处理对土壤全氮含量并无显著影响,但却显著增加了硝态氮和铵态氮含量。这可能是氮肥的施用刺激了土壤中微生物活性,加速了土壤有机氮的分解;另外,施肥后土壤中的无机氮优先供给地上植物的生长发育用以增加地上生物量,从而导致全氮含量无显著变化。
施肥和灌溉不仅直接影响土壤肥力和植被生长,还会造成氮循环相关微生物群落结构的改变。徐鹏霞[45]研究表明,长期施肥能够促进氨氧化细菌的生长,氨氧化细菌amoA基因的丰度与硝态氮显著正相关,这与农田施氮肥提高氮循环相关基因丰度的结果相符合[46]。土壤水分状况也会对硝化、反硝化作用产生一定影响。杨亚东等[47]在探讨灌溉频次对硝化作用的影响时发现,灌溉显著提高了土壤的硝化活性和氨氧化古菌amoA基因的丰度,降低了氨氧化细菌amoA基因的丰度;改变了氨氧化细菌和氨氧化古菌的群落结构,且土壤的硝化潜力与氨氧化古菌amoA基因的丰度正相关。但也有研究表明,硝化速率随水分增加呈下降趋势,并非灌溉的水量越多,硝化作用就越强;当土壤含水量为20%左右时,硝化作用较强[48]。
施肥方式对反硝化细菌群落调节作用的影响也存在很大差异。多数研究表明,有机肥比传统肥料更有助于提高微生物活性及生物量。相关学者评估有机、综合和传统肥料处理的苹果(Malusdomestica)园中的反硝化潜力以及其他土壤生物、化学特性时发现,施用有机肥减少了土壤硝态氮的淋溶量,提高了反硝化活性和效率[49]。反硝化作用的加强得益于有机肥对反硝化微生物的刺激作用,反硝化群落扩张、反硝化菌的比例提高。由于反硝化作用的增强,氮素多以气态的形式损失,这是土壤硝态氮淋溶量较低的一个重要原因;此外,有机肥的施用提高了地上植被覆盖度,增加了硝态氮截留量。其他研究也得出类似结论,作物收获后土壤剖面以及土壤溶液中的硝态氮浓度随施氮量的增加而显著提高,随灌溉量的增大而下降;持续性降水是硝酸盐淋溶的主要原因[50]。虽然反硝化作用能够有效降低硝态氮的淋溶量,但此过程释放的N2O是主要的温室气体,施用有机肥显著增加土壤N2O的排放量且pH 是控制其排放的主要变量[51-52]。因此,考虑到全球气候变化和硝酸盐污染,对水、肥进行科学管理可以降低环境污染,促进草地土壤氮素良性循环。
刈割作为草地利用和管理的主要方式,对草地生态系统起至关重要的调控作用,包括碳氮含量、养分循环过程等变化。刈割主要从频次方面对土壤氮循环产生影响。Li等[53]对半干旱草地进行不同频次的刈割(2年1次、3年2次、1年1次和1年2 次),研究发现中频刈割(2年1次、3年2次和1 年1次)下土壤碳氮储量显著上升,且净氮矿化速率提高;而高频次的刈割(1年2次)显著降低土壤碳氮储量。这与研究刈割频次对呼伦贝尔草原碳氮影响的[54]结果较为一致,造成这一现象的原因是适宜频次的刈割通过提高草地植物物种多样性提高了总氮含量;而高频次刈割通过清除地上生物量,减少了凋落物的沉积和光合碳向根系的转移,导致草地生态系统养分的输出大于输入、净氮矿化速率降低[55-56]。刈割通常与施肥管理相配合,施肥通过改善土壤性质以加速养分循环,进而提高草地生产力,但在一定程度上降低了植物多样性;而刈割是维持草地植物多样性的有效手段[57]。因此,在生产实践中,要综合考虑草地生产性能和生态功能,选择适宜的刈割频次,同时配合适当的施肥管理,使草地在刈割状态下也能保持养分平衡,从而实现草地生态系统的可持续利用。
随着放牧和人为扰动的加剧,草地退化日益严重,为了防止草地的持续退化,必须采取相应措施以维持草地生态系统的稳定。围封被认为是恢复退化草地和防止草地退化的一项简单、经济有效的重要措施[58]。相较于放牧草地,围封10多年的草地植被高度和盖度均有所增加,有效降低了风蚀和水蚀的程度,进而减少氮素流失,显著增加了土壤中的全氮浓度[58-59]。草地围封后免受动物践踏,土壤理化性质得以改善且凋落物的积累增多,有利于土壤有机质的分解和养分循环。已有研究表明,一方面,围封使得以枯落物和根系分泌物的形式输入土壤的有机质增加,土壤全氮含量得以提高;另一方面还可以提高土壤微生物的活性,增强土壤氮素的固定和转化,进而提高氮素有效性[60-61]。然而,草地土壤氮循环对围封的响应因围封时间长短而异。Hu 等[62]研究发现,长期围封(38年)使内蒙古半干旱草地土壤剖面中的总氮、铵态氮和总磷含量下降,尤其降低了表层土壤养分的矿化程度。此外,长期禁牧围封会导致严重的土壤酸化,土壤酸化是指土壤吸附性复合体接受了一定数量的氢离子或铝离子,使土壤中碱性离子淋失、pH 降低形成酸性土的过程[63]。因此,草地围封时间不宜过长,否则不但不利于草地植被的正常发育反而阻碍植被更新再生,影响草地的养分循环;实际工作中应根据草地退化程度确定合理的围封年限,恢复草地的同时维持草地较高生产力。
2.4.3 非生物因子
氮沉降是生物地球化学循环中的一个重要环节,显著影响草地土壤的碳氮循环。化石燃料的燃烧、化肥的使用以及其他人为活动,使20世纪内大气氮沉降量迅速增加。我国作为农业大国,农业集约化日益加强,氮沉降速率迅速增长,从20世纪80年代的13.2 kg·hm−2增加到21世纪初的21.1kg·hm−2[64]。氮沉降主要通过影响土壤氮的有效性、土壤性质、微生物以及凋落物的分解来影响氮转化过程。作为氮输入的重要来源,氮沉降直接影响草地氮储量和有效氮含量。在贝加尔针茅草原模拟氮沉降时发现,氮沉降显著增加了土壤中总氮含量,有效氮、NO3−-N含量均有所提高[65]。这一结果与在青藏高原高寒草地得出的研究结论一致[66]。但在亚高山草原的氮沉降模拟试验却表明氮添加对草地土壤总氮含量无影响,这可能与草地的氮限制以及土壤微生物与植物对资源的竞争有关[67]。在氮富集引起的土壤酸化背景下,微生物组成改变、呼吸作用减弱,减少了对凋落物的分解量,导致有机层碳含量增多,打破了植物源碳对微生物的控制,削弱了氮循环过程中植物、微生物的相互作用[68]。但是,对高寒草原、温带草原进行长期施氮处理结果发现,高寒草原的石灰性土壤具有强酸缓冲能力,因此总氮含量的上升、氮素有效性的提高刺激了微生物生长,加强了矿化作用和硝化作用;而温带草原的中性土壤对pH较为敏感,细菌、真菌的丰度下降,导致草原土壤的自养硝化速率和氮固定速率显著下降[69]。
草地生态系统存在地上和地下两个营养库,凋落物是连接两个营养库的重要纽带。氮沉降引起氮有效性、土壤pH 改变,导致凋落物质量、微生物的群落组成以及酶活性发生变化,进一步影响凋落物的分解。目前,国内外研究者对氮沉降背景下草地凋落物分解的关注度较高,但结论存在不一致性。在半干旱草原模拟氮沉降结果表明,随着氮添加速率增加,凋落物的分解速率持续加快。凋落物分解速率加快主要有两大原因,一是氮沉降驱动土壤pH 下降;二是C/N的下降促使真菌与细菌比值显著增高,从而促进凋落物的分解[70]。而Freedman 等[71]的研究却指出,氮沉降显著增加土壤中碳储存量,凋落物的分解代谢并不受氮沉降的影响,这可能与分解凋落物的腐生细菌、腐生真菌的代谢潜力以及酶活性等有关系。氮沉降对凋落物分解的影响因草地类型、环境条件、氮添加水平而异,具体的影响机制还需进一步研究。
氮沉降增加了草地土壤的总氮含量,但硝化作用、反硝化作用的增强,加剧了土壤N2O气体的排放。研究表明,内蒙古温带草原的N2O排放与模拟氮沉降水平呈线性关系,特别是高水平时显著增加了N2O 的排放量,这主要归结于温带草原良好的水热条件对硝化、反硝化的促进作用[72]。然而,很多研究指出,氮沉降后N2O量的变化取决于输入的氮的形态。许可等[73]通过不同形态氮添加的处理发现,氮添加能显著增加N2O排放量,且混合态氮处理组的排放量高于铵态氮处理组和硝态氮处理组。这表明不同形态的氮处理对N2O排放的影响取决于N2O是主要来源于硝化途径还是反硝化途径[72]。除此之外,人为活性氮的释放引起的氮沉降缓解了草地土壤氮限制,提高了氮的矿化–固定化周转速率,使更多还原态氮被硝化成氧化态氮,而氧化态氮易通过淋溶和反硝化作用流失,这也是氮沉降后氮素损失的一个重要原因[69]。今后应关注不同氮输入量及氮输入类型在不同草地类型、不同土壤性质下的潜在机制的对比研究,以便更准确地获得氮沉降对草地生态系统的不良影响,制定合理措施以维持草地生态系统的稳定。
近几十年来,工业化的迅速发展导致全球碳排放量增加、全球平均温度升高,这势必会对土壤氮素转化产生直接或间接影响,CO2浓度升高以及其带来的气候变暖问题对氮循环的影响已经成为全球的研究热点[74]。相关研究[75]表明,大气CO2浓度升高促进了植物的光合作用和生长速率,使地上与地下的碳分配发生改变,从而影响生态系统的养分循环。其中,升高的CO2会影响土壤氮循环过程,调节植物氮的有效性,氮有效性的增加可以促进植物对氮的吸收,从而提高草地生态系统的净初级生产力[76]。例如,Luo等[77]的研究指出,升高的CO2会增加土壤中碳的输入,碳通过菌根侵染或细根量的变化来增加微生物对氮的需求,造成微生物与植物对土壤有效氮的竞争,改变了氮的供给机制,导致生态系统中的渐进氮限制现象。CO2浓度升高还会显著促进N2O的排放。Liu 等[75]的研究中发现,随着CO2浓度上升,草地N2O的释放量增加4.6%。CO2浓度升高可能通过两种方式来改变土壤中N2O的排放。首先,CO2富集促进植物的生长发育,生物量(凋落物、根际分泌物等)增加,进而提高基质碳的有效性,为硝化、反硝化提供更多能量。其次,CO2浓度升高,土壤含水量上升、呼吸作用增强,营造了厌氧环境,更有利于反硝化的进行。随着大气CO2浓度不断上升,生态系统的碳输入越来越多,如果碳固定没有随之增加,那草原可以通过N2O排放的正反馈转化为温室气体源。然而,Kettunen等[78]却得出相反结论,通过两个水平的CO2和氮肥处理,对一个混合林进行N2O值测定,结果表明,除非处于有利的反硝化条件,即高地下水位,否则增加大气CO2供应可减少矿质土壤中N2O的排放。CO2对N2O排放的影响因试验环境、生态系统类型、管理实践等的不同而产生差异,反硝化是产生N2O的主要途径,如果只是测定CO2浓度升高情况下的N2O的通量则会低估反硝化过程中的氮素损失。
大气CO2浓度以及温度升高还会促进植物的生长发育,通过光合产物以及根际分泌物的变化对土壤环境产生直接或间接影响,从而间接地影响氮循环微生物的生物化学活性。研究表明,CO2浓度升高抑制了稻田土壤中反硝化菌活性,主要是因为开放式大气CO2浓度升高促进了植物生长及土壤其他微生物活性,使反硝化微生物在资源竞争方面处于劣势,导致反硝化速率下降;但这种抑制作用在施入氮肥后得到一定的缓解[79]。然而,CO2浓度升高,不稳定碳通过根系进入土壤,增加根系生物量的同时为土壤微生物提供更多的能量,进而显著提高了反硝化速率和N2O释放量[75]。气候变暖使土壤温度升高,刺激土壤微生物活性,进而促进有机质的降解、提高矿化速率[80-81]。同时,CO2浓度升高和气候变暖在一定程度上改变了土壤氨氧化古菌(ammonia-oxidizing archaea,AOA)丰度和反硝化细菌多样性,反硝化细菌群落却没有因此产生明显差异[82]。但是,在青藏高寒草甸进行模拟增温试验时发现,单独增温并不会对土壤矿质氮库、植物氮吸收、AOA 和氨氧化细菌(ammonia-oxidizing bacteria,AOB)丰度产生显著影响[83]。因此,大气CO2浓度上升及气候变暖对氮循环相关微生物的间接影响至今还没有统一的结论,这种结论间的较大差异可能与植物类型、CO2浓度、土壤的理化性质等有关。
随着气候变暖,全球降水格局发生变化。草地土壤氮循环与降水关系密切,Yang 等[84]在中国松嫩草甸草原研究半干旱草地对干旱的响应机制时发现,夏季干旱主要通过非生物因子的变化(土壤水分、总氮含量的下降)来改变氮有效性,而冬季干旱通过生物因子(植物和微生物生物量的降低)的变化影响氮有效性。一般来说,降水提高了土壤湿度,可以刺激土壤氮素转化过程,氮吸收、矿化和硝化作用随降水量的增加而增强,但降水会导致无机氮淋失,使无机氮含量下降;相反,降水量下降致使土壤湿度降低,抑制土壤微生物活性,降低氮矿化速率,土壤无机氮含量上升[85-86]。Wang 等[87]模拟降水对碳氮循环的调节作用发现,土壤水分通过影响土壤微生物的休眠与否对碳氮循环产生强有力控制,土壤干燥时,微生物活动受抑制,降低异氧呼吸和氮矿化。但也有相关研究指出,矿化在降水梯度上没有显著变化,可能是研究所设置的各处理间土壤微生物的变化差异较小[88];也有可能是因为土壤微生物对干旱土壤具有一定抵抗能力[89]。此外,降水还会影响草地土壤中温室气体的排放,研究表明,降水量增加的同时也增加了N2O和CH4的排放通量,这是因为草地的植物群落组成在降水条件下发生变化[90]。基于目前全球降水格局的变化,研究降水变化对草地土壤氮循环的影响,对在全球变化下科学管理草地生态系统的养分循环、维持生态系统的健康与稳定至关重要。
3 总结与展望
氮是限制草地生命力的主要元素之一,草地土壤作为生物地球化学循环的核心循环,影响调控着全球粮食供给的安全性、草地土壤的退化与修复、水土资源的保持与保护、全球气候变化等,进而影响人类社会的可持续发展。梳理和总结草地土壤氮循环的影响因子,可为科学揭示氮循环过程和影响机制、最终调节氮转化过程进而提高土壤氮素有效性且降低其造成的负面生态环境影响奠定基础。
基于上述对草地土壤氮循环研究现状的分析,提出如下建议:1)利用同位素示踪与标靶基因相结合的方法完善基于土壤基质的氮保留与损失机制以及微生物驱动机制,有助于研究全球变化背景下生态系统的调控和适应策略。2)运用化学计量学、生物计量学、生态计量学的基本原理和方法,结合同位素标记和分子生物学技术,对氮循环过程中氮的长期分配进行量化分析以完善氮循环模型,将实证研究、概念分析、模型开发三者综合以弥补目前草地土壤氮循环方面的关键知识空白。3)草地土壤氮循环过程以及其影响因素具有复杂性,目前的研究多局限于某一因素或某一环节,未来需要理清多个全球气候变化因子耦合对氮循环过程的影响,形成系统的研究网络体系以便更好地预测氮循环过程。4)基于野外长期模拟试验,利用稳定同位素示踪等技术,系统研究水、碳、氮耦合对草地氮循环过程中功能微生物的种群结构适应性机制和种群演替规律,揭示在全球变化格局下微生物功能多样性对元素循环的响应,建立植物–土壤–微生物耦合的调控途径与机制。
