5-Aza-dC对牛成肌细胞MyoD1启动子甲基化及mRNA表达的影响
2021-09-27田念念宋林锦许厚强
李 雄,田念念,宋林锦,陈 晨,许厚强*
(1.贵州大学 高原山地动物遗传育种与繁殖教育部重点实验室/贵州省动物遗传育种与繁殖重点实验室,贵阳 550025;2.贵州大学动物科学学院,贵阳 550025)
表观遗传调控多发生在相互作用水平上,在该水平上的研究可深入了解表观遗传学对生物学过程的促进作用。大量研究表明,表观遗传学是指基因的核苷酸序列不发生改变,而基因功能发生稳定的、可遗传的变化,主要包括非编码RNA调控[1]、DNA甲基化[2]、染色质重塑[3]、组蛋白修饰[4]等。DNA甲基化(DNA methylation)是指在DNA甲基化转移酶的作用下,活性甲基化合物(如S-腺苷基甲硫氨酸)将甲基催化转移到基因组CpG二核苷酸的胞嘧啶5’碳位共价键结合一个甲基基团,导致染色体结构、DNA构象、DNA稳定性等发生变化,从而控制基因表达[5]。对DNA甲基化研究主要集中于主动甲基化和去甲基化方面,DNA高甲基化能够减弱或者沉默基因的表达,而去甲基化时则能使基因活化或重新表达[6]。5-Aza-dC是目前用得较多的去甲基化方法之一,其对研究DNA甲基化对表观遗传的影响及特异性基因的表达和调控机理有着重要的作用。5-Aza-dC是一种胞唾陡类似物,常用名为地西他滨甲基化酶抑制剂,是目前已知的较为有效的一种去甲基化制剂[7]。5-Aza-dC大多应用于医学方面,如Greville等[8]发现,5-Aza-dC处理后,从化疗敏感/非转移细胞系中分泌的n-聚糖的分支和唾液化增加。这些变化与卵巢癌细胞系中MGAT5和ST3GAL4转录本mRNA表达水平升高有关。Xie等[9]研究发现,5-Aza-dC能有效抑制DNMTs的表达和活性,逆转MnSOD和GSTT1的表达。所以,通过药物、试剂等对相关基因去甲基化探究其表达影响,也是从侧面研究甲基化机制的一种方法。
近年来,大量研究表明,DNA甲基化所引起的表观遗传对于动物的生长发育有重要影响,并筛选与生长相关的甲基化位点作为分子标记[10]。肌肉组织是动物机体的重要组成部分,而DNA甲基化也能调控肌肉的发育过程[11]。Li等[12]研究了3个品种猪不同部位骨骼肌,发现品种间、性别间存在DNA甲基化的相似性和特异性,并鉴定了差异甲基化区域。李秀金等[13]通过对五指山和长白猪DNA甲基化和转录组数据的联合分析证实,DNA去甲基化酶Tet1在猪胚胎成肌细胞和C2C12细胞中通过Myogenin启动子去甲基化,上调Myogenin表达,从而促进成肌细胞的分化。万火福等[14]用比色法测定不同品种肌肉组织DNA甲基化水平,发现DNA甲基化水平在不同品种及不同组织中会呈现一种动态变化,且高水平的DNA甲基化可能促进霞烟鸡腿肌组织的发育。综上表明,不同畜禽肌肉发育过程中的甲基化模式具有一定特异性,不同家畜中甲基化状态会产生特定变化,这种甲基化状态变化能够通过调控不同阶段相关基因的表达,从而保证肌肉发育正常进行。
动物肌肉发育是一个极其复杂的过程,动物体内相关基因的甲基化状态也呈现出动态变化趋势[15]。肌分化因子(myogenic differentiation 1,MyoD1)是一种骨骼肌特异性转录因子,能够识别、启动骨骼肌特异基因的转录,具有诱导成纤维细胞和其他非骨骼肌细胞分化为骨骼肌细胞的能力[16]。近年来,MyoD1甲基化在猪[17]、羊[18]、鸡[19]等家畜上主要通过对肌肉进行全基因组甲基化检测其甲基化水平;沈兰等[20]发现,MyoD1的甲基化谱可能有助于预测接受标准化学放射治疗的浸润性宫颈癌患者的反应。目前,MyoD1甲基化对牛骨骼肌细胞发育影响的研究报道较少。本试验以关岭牛为研究对象,利用甲基化酶抑制剂(5-Aza-dC)处理牛成肌细胞,研究MyoD1启动子区甲基化水平及其对成肌细胞生长发育和增殖的影响,为关岭牛的改良提供理论参考。
1 材料与方法
1.1 试验材料
牛成肌细胞来源于贵州大学动物科学学院动物种质资源库。
1.2 主要试剂
DMEM/F12培养基、胎牛血清、TRIzol 试剂购于Gibco公司;5-Aza-dC(5-氮杂-2′-脱氧胞苷)、细胞凋亡检测试剂盒、细胞周期试剂盒均购自美国Sigma公司;2×Es Taq Master Mix、DM 2000 Marker、丙三醇、无水乙醇、DEPC 水购自康为世纪生物科技有限公司;SYBR-Green染料(SYBR Green PCR Master Mix)购自Bio-Rad公司;二甲基亚砜购于Hyclone 公司;逆转录试剂盒和CCK8试剂盒购自MCE公司;一抗(Desmin)购自鼎国生物技术有限责任公司;二抗购自Jackson immuno research。
1.3 试验方法
1.3.1 细胞复苏及鉴定 取出冻存的细胞,复苏后转入含15%胎牛血清的DMEM完全培养基中。置于CO2培养箱(培养条件5% CO2、饱和湿度、37 ℃)中继续培养,让其长到对数生长期。将培养的细胞接种于6孔细胞培养板上,待其生长至85%左右,进行细胞鉴定;试验参考间接免疫荧光(indirect immunofluorescence,IIF)的方法执行。
1.3.2 CCK8检测不同浓度5-Aza-dC对成肌细胞增殖凋亡的影响 细胞复苏后,当细胞长到85%以上进行分瓶,当关岭牛成肌细胞处于对数生长期时,用0.25%胰蛋白酶消化约4 min,并进行计数,以每孔5×103个接种于96孔板内(每孔加入100 μL)。置于CO2培养箱内培养12 h;12 h后加入添加了不同浓度5-Aza-dC的培养基,浓度分别为0.05、0.1、0.2、0.3 μmol·L-1以及空白组(0 μmol·L-1),每组设3个 重复,分别处理0、24和48 h。在每个孔内加入10 μL CKK-8试剂,酶标仪(450 nm)检测吸光密度(OD);确定在本实验室条件下对细胞生长影响最小的5-Aza-dC最适浓度,并以此浓度进行后续试验。
1.4 流式细胞仪检测细胞凋亡
将对数生长期细胞接种于6孔板内,标记处理组和空白组置于CO2培养箱内培养12 h,用含有5-Aza-dC的DMEM/F12(“1.3.2”获得5-Aza-dC最适浓度)培养基换液,培养48 h,用3 mL PBS溶液轻轻润洗培养板内细胞;加入1 mL 0.25%不含EDTA 的胰酶消化;将细胞轻轻重悬于所保留的培养基中,密度大约为1×106个·mL-1,1 000 r·min-1离心6 min,弃去上清液,PBS洗涤2次,加入冰预冷的70%的乙醇固定,4 ℃,3 h;离心弃去固定液,3.5 mL PBS重悬5 min;将细胞悬液转移到干净的离心管内,加入2 μL PI,5 μL Annexin V-FITC,室温避光反应15 min;室温1 000 r·min-1离心5 min,去除上清;将细胞用0.5 mL Propidium Iodide预冷缓冲液,用流式细胞仪检测分析。
1.5 流式细胞仪检测细胞周期
5-Aza-dC处理细胞48 h后,用预冷PBS洗2次,加入1 mL 0.25%不含EDTA的胰酶消化,1 300 r·min-1离心6 min,获取细胞沉淀,弃上清,重悬细胞,迅速打入3.5 mL 70%预冷的乙醇中,吹打均匀,4 ℃储存过夜;离心乙醇固定过的细胞,弃上清;洗涤细胞3次以去除残留的乙醇;加入100 μL RNase A重悬细胞,37 ℃水浴30 min,加入500 μL PI染色液,4 ℃避光反应30 min;应用流式细胞仪检测。
1.6 qRT-PCR检测MyoD1、DNMT1及相关基因mRNA表达
1.6.1 RNA提取及第一条cDNA的合成 5-Aza-dC处理细胞48 h后,处理组和空白组的细胞用Trizol提取总RNA。将RNA定量至2 ng,根据逆转录试剂盒进行反应合成cDNA,反应产物置于-20 ℃保存备用。
1.6.2 引物的设计与合成 根据NCBI公布的牛MyoD1(NM_001040478.2)、DNMT1(NM_182651.2)、Caspase-9(NM_001205504.2)、Bax(NM_173894.1)、Bcl(CX950904.1)、CyclinA2(NM_001075123.1)、CyclinB1(BC116011.1)、CyclinD(XM_024991034.1)以及内参基因GAPDH(NM_001034034.2)。利用Primer Premier 5.0软件进行定量引物设计,最后由重庆擎科生物有限公司合成,引物序列见表1。
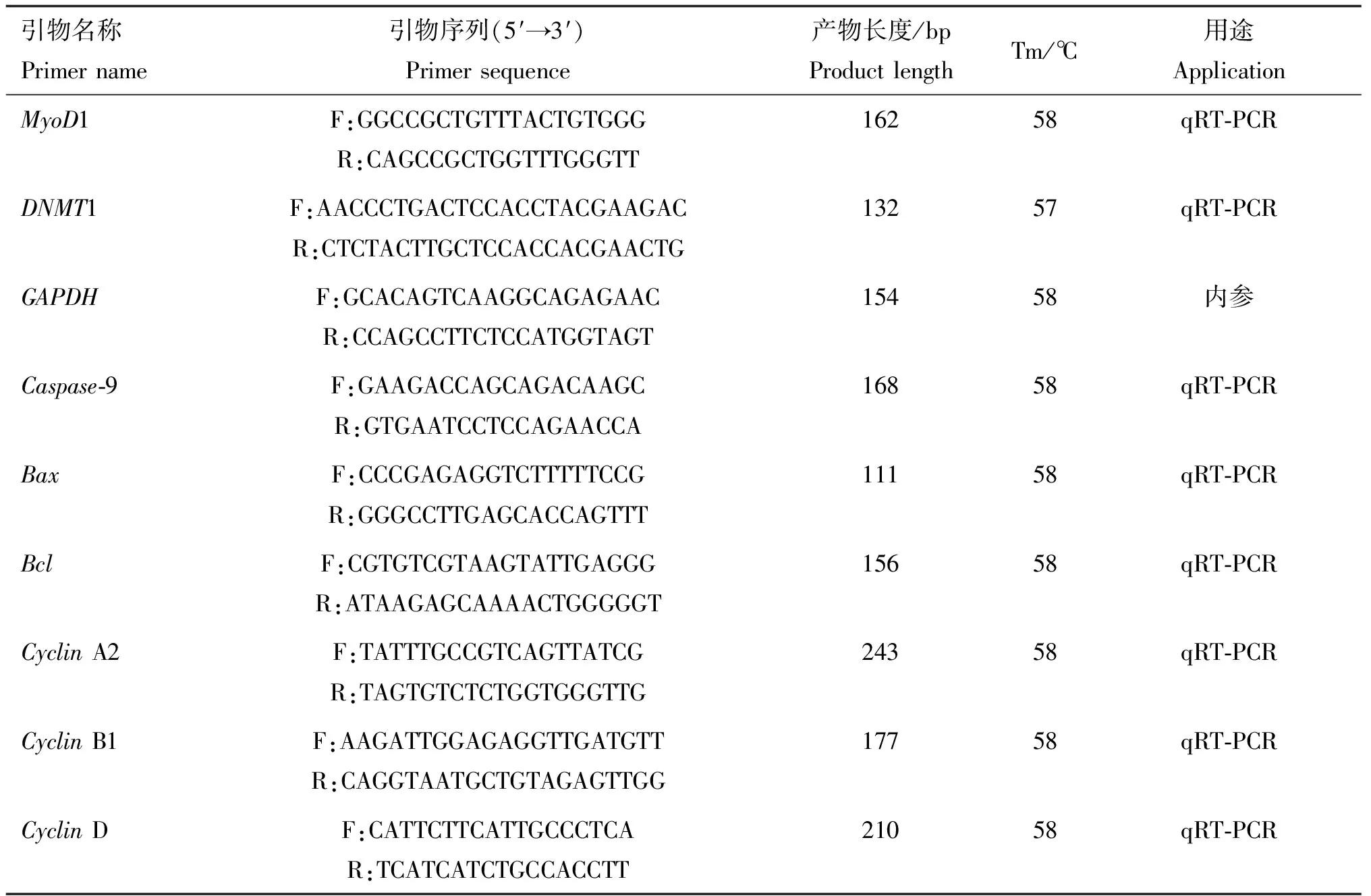
表1 引物序列信息
1.6.3 qRT-PCR 检测MyoD1、DNMT1及相关基因表达 以处理组和空白组cDNA为模板分别扩增MyoD1以及相关基因,采用SYBR Green 荧光染料法在实时荧光定量PCR仪上进行定量反应。根据伯乐公司Bio-rad iTaq Univer SYBR(1725121)荧光定量qPCR mix试剂盒制作上机 PCR 混池,混池体系为10 μL:ULtraSYBR Mixture(High ROX)5 μL,上、下游引物各1 μL,cDNA 模板0.7 μL,ddH2O 2.3 μL。反应条件:95 ℃ 1 min;95 ℃ 10 s;58 ℃ 30 s,40 个循环后,进行熔解曲线分析;以每5 s 上升0.5 ℃的速率从65 ℃升高到95 ℃,荧光信号在循环结束时检测,每个样品做3 个生物学重复。
1.7 高通量检测MyoD1启动子区甲基化
依据原始提供的牛MyoD1启动子区区域进行引物设计(表2),选取检测位置区域(NC_037342.1)将样品送往重庆擎科生物科技有限公司进行高通量测序。

表2 甲基化测序引物序列
1.8 统计分析
运用Excel 2010软件对各组织中目的基因和内参基因mRNA表达量进行处理,运用2-ΔΔCt法计算。采用独立样本t检验和单因素方差分析进行数据处理(SPSS 21.0),多重比较采用Duncan’s多重极差检验法,运用Graphpad6 作图,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结 果
2.1 间接免疫荧光细胞鉴定
如图1所示,Desmin抗体发生特异性结合,呈阳性反应显红色,细胞核在DAPI的染色下呈蓝色,Merge后能完全重合,表明培养的细胞为成肌细胞。
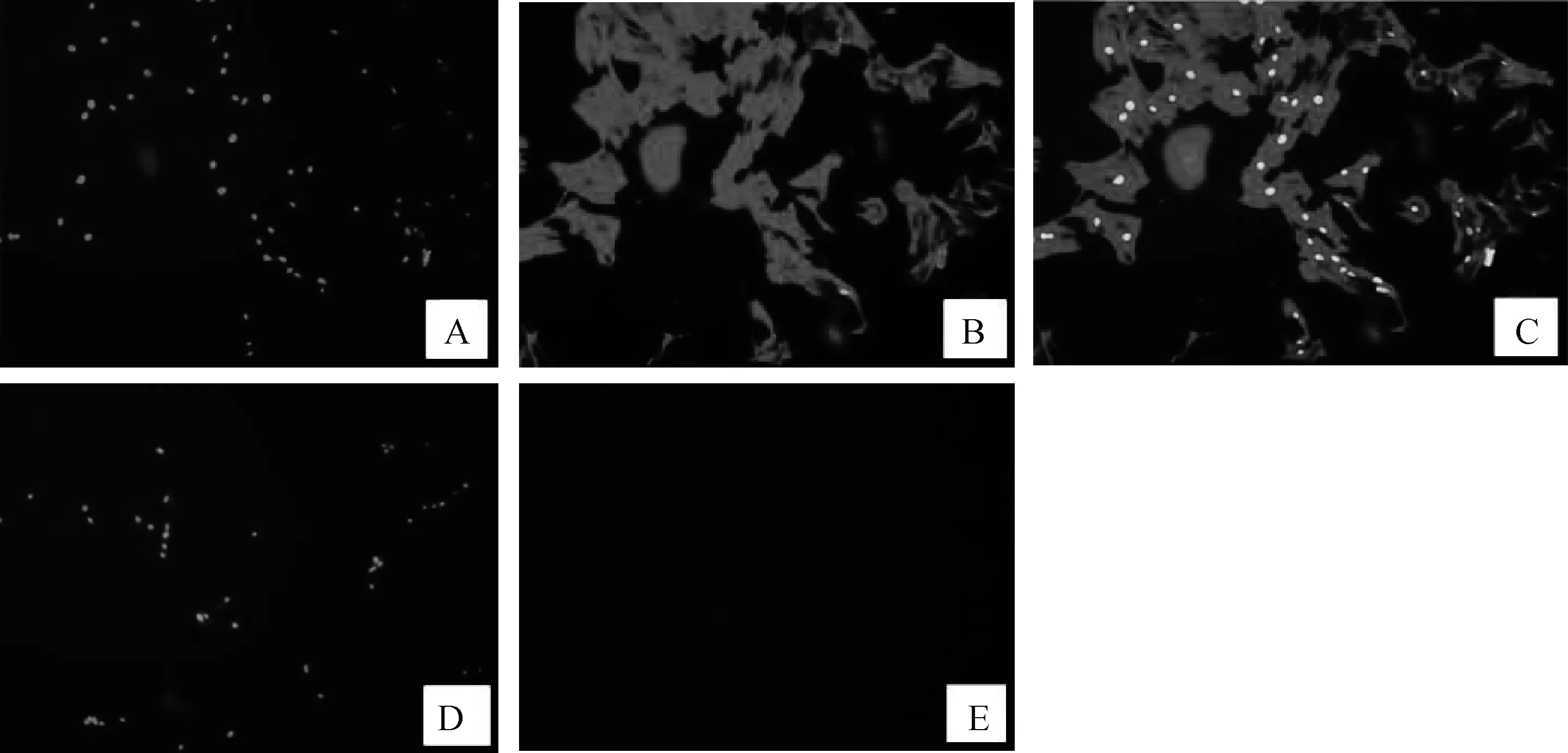
A.DAPI核染;B.Desmin染色;C.合图Merge;D.DAPI核染对照;E.不加Desmin一抗对照染色
2.2 CCK8检测不同浓度5-Aza-dC对成肌细胞增殖的影响
本试验在成肌细胞中添加不同浓度的甲基化转移酶抑制剂5-Aza-dC后,用CCK8检测0、24、48 h成肌细胞增殖情况。表3可以看出,与对照组相比,成肌细胞中添加0.05或0.1 μmol·L-15-Aza-dC时细胞存活率呈上升趋势,但是随着浓度增加细胞存活率逐渐下降,其中0.3 μmol·L-15-Aza-dC浓度下细胞存活率极显著下降(P<0.01);处理24 h后,0.1 μmol·L-15-Aza-dC处理细胞的存活率呈显著上升(P<0.05),在0.1 μmol·L-1存活率最高。处理48 h后,细胞存活趋势与24 h相同,但是同24 h相比提高不明显。表明0.1 μmol·L-15-Aza-dC处理组适合用于后续试验。

表3 不同浓度5-Aza-dC对细胞存活率的影响
2.3 流式细胞仪检测成肌细胞凋亡
从图2可知,0.1 μmol·L-15-Aza-dC处理的细胞凋亡率较空白组极显著增加(P<0.01)。为进一步检测甲基化转移酶抑制剂5-Aza-dC是否与Bcl、Bax、Caspase-9等细胞凋亡因子有关,通过qRT-PCR检测,0.1 μmol·L-15-Aza-dC处理的细胞能够极显著的降低细胞抗凋亡因子Bcl的表达(P<0.01),极显著的提高促凋亡因子Bax表达(P<0.01),显著提高促凋亡因子Caspase-9的表达(P<0.05)。
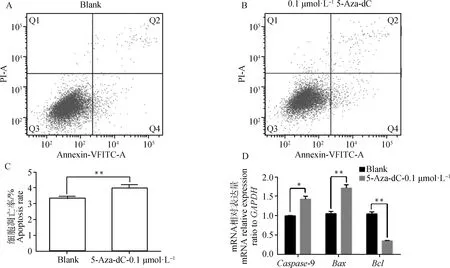
A.空白组细胞凋亡;B.试验组细胞凋亡;C.细胞凋亡率;D.细胞凋亡因子mRNA的表达。Q1.机械性损伤细胞;Q2.晚期凋亡细胞;Q3.正常细胞;Q4.早期凋亡细胞
2.4 流式细胞仪检测成肌细胞周期
通过流式细胞仪检测0.1 μmol·L-15-Aza-dC处理的细胞及空白组细胞,结果显示(图3),0.1 μmol·L-15-Aza-dC处理组G0/G1期细胞比例较高于空白组,但差异不显著(P>0.05),S期及G2/M期细胞低于空白组,差异也不显著(P>0.05)。为进一步检测甲基化转移酶抑制剂5-Aza-dC对细胞周期相关因子CyclinB1、CyclinA2、CyclinD 表达的影响,通过qRT-PCR检测相关细胞因子的表达,发现5-Aza-dC显著的提高了周期因子CyclinA2的表达(P<0.05),对细胞因子CyclinB1、CyclinD的表达影响不显著(P>0.05)。

A.空白组细胞周期;B.试验组细胞周期;C.细胞周期不同时期比例;D 细胞周期因子mRNA的表达
2.5 qRT-PCR 检测MyoD1、DNMT1 mRNA相对表达量
2.5.1 PCR扩增结果 由图4可知,PCR扩增产物条带清晰,没有拖带和二聚体,片段大小符合试验预期。
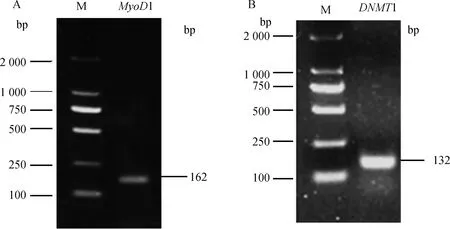
A.关岭牛MyoD1基因荧光引物电泳图;B.关岭牛DNMT1基因荧光引物电泳图。M.DNA相对分子质量标准
2.5.2 Real-time PCR 检测结果扩增曲线和峰图 由图5、图6、图7可知设计的引物通过实时荧光定量PCR检测,扩增曲线拐点清晰,熔解曲线均为单峰,无杂峰,引物特异性好。

A.MyoD1扩增曲线;B.MyoD1熔解峰
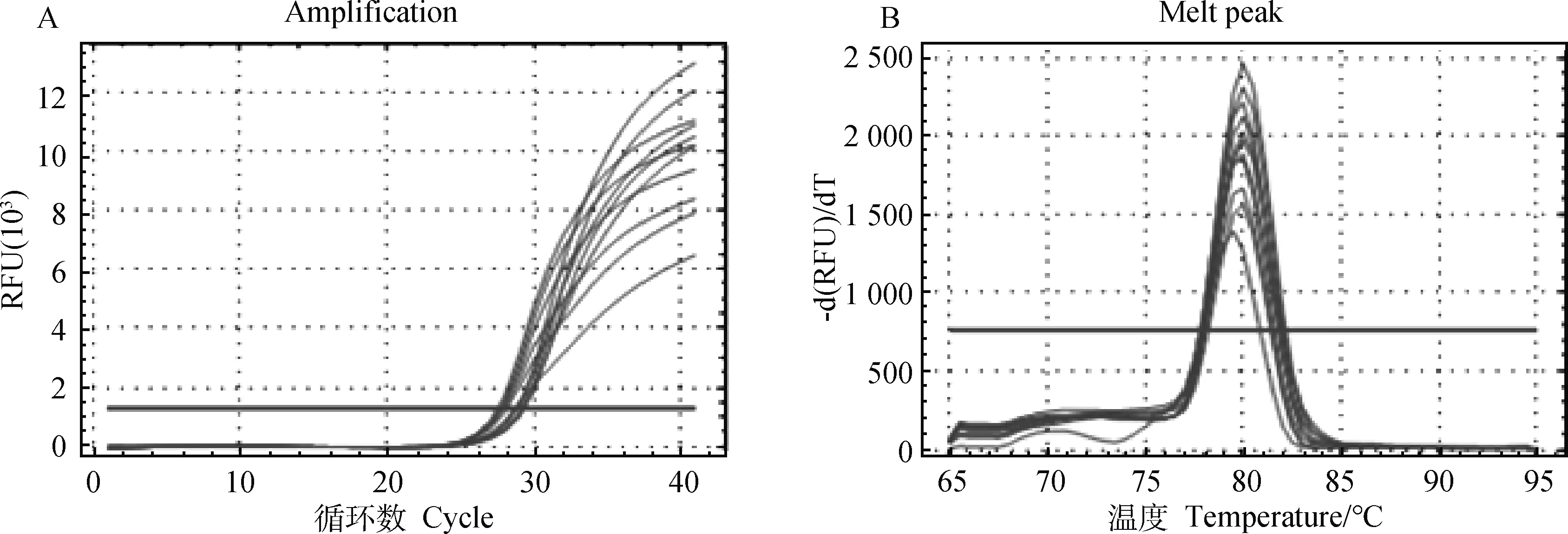
A.DNMT1扩增曲线;B.DNMT1熔解峰
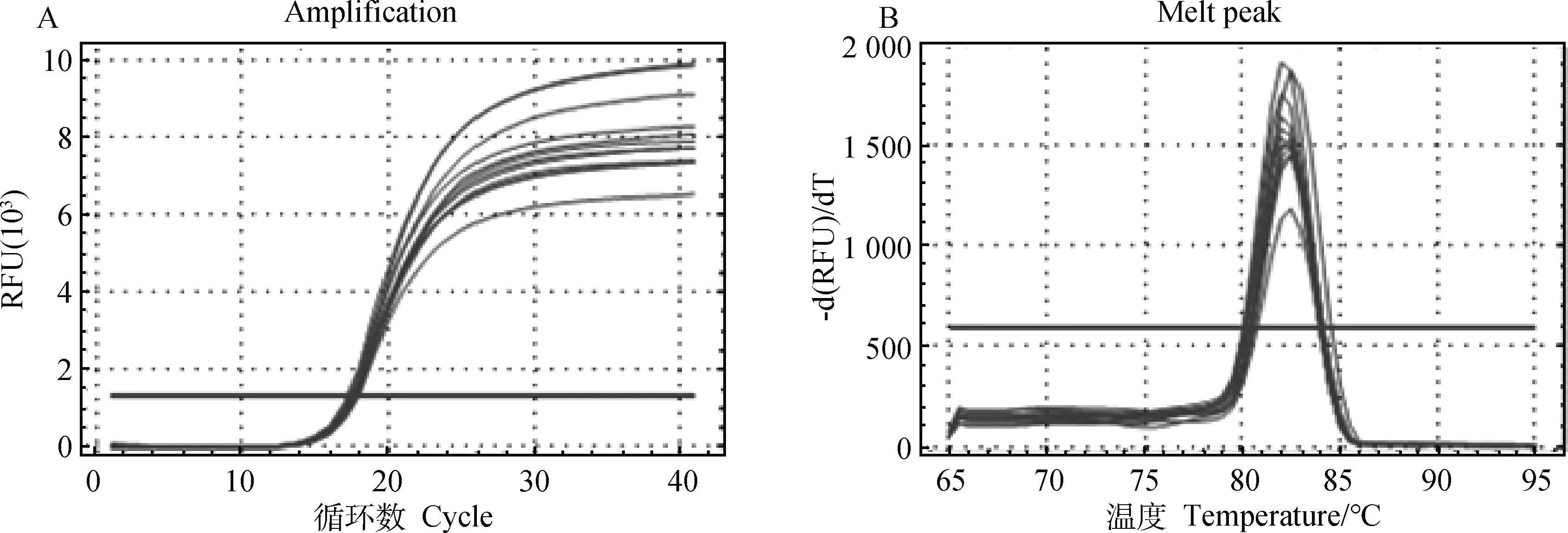
A.GAPDH扩增曲线;B.GAPDH熔解峰
2.5.3MyoD1、DNMT1基因表达分析 由图8可知,空白组中MyoD1的mMRNA的表达量极显著低于试验组的表达量(P<0.01),DNMT1的表达量也极显著低于试验组(P<0.01)。
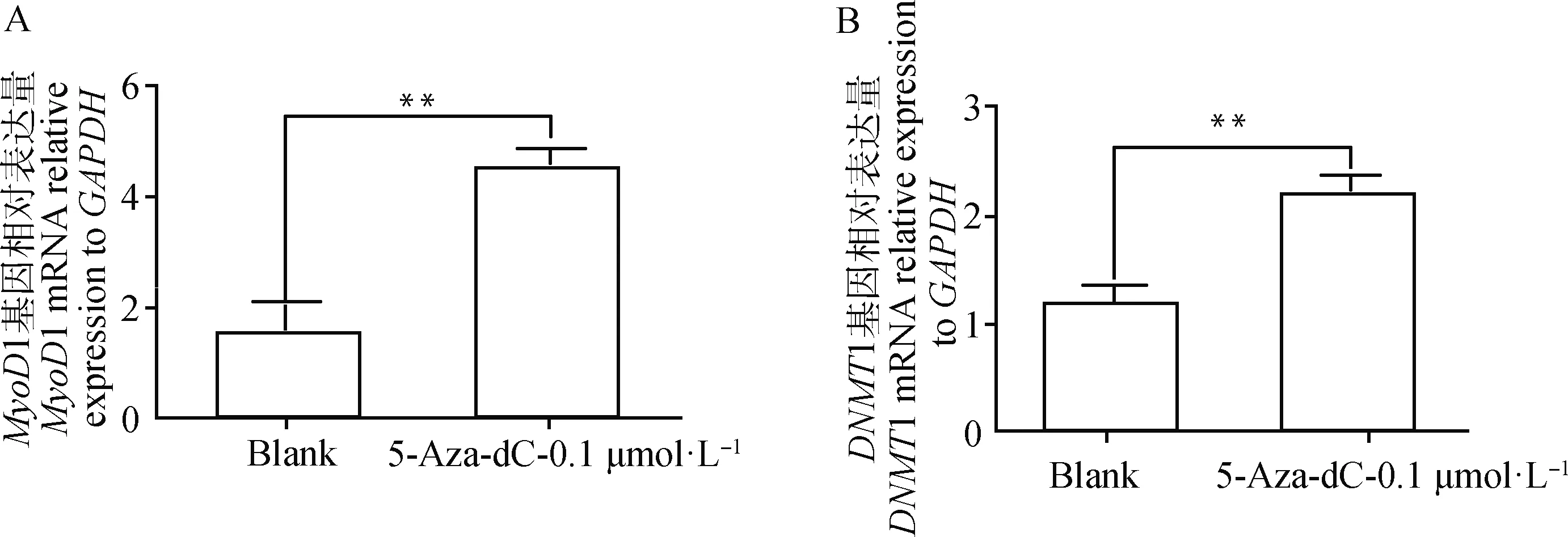
A.MyoD1相对表达量;B.DNMT1相对表达量
2.6 MyoD1启动子区甲基化检测
由表4可知,处理48 h后,空白组的MyoD1启动子区CpG甲基化率为28.0%,CHG甲基化率为2.0%,CHH甲基化率为1.9%,试验组的MyoD1启动子区CpG甲基化率为19.8%,CHG甲基化率为1.3%,CHH甲基化率为1.8%。并且0.1 μmol·L-15-Aza-dC处理的试验组极显著的降低了MyoD1启动子区的甲基化率(P<0.01,图9)。

表4 5mC 位点覆盖统计

图9 MyoD1基因启动子甲基化率
3 讨 论
DNA甲基化作为一种相对稳定的修饰状态,在DNA甲基转移酶的作用下,可随DNA的复制过程遗传给新生的子代DNA,是一种重要的表观遗传机制[21]。DNA甲基化通过DNA甲基转移酶(DNA methylthansferase,DNMT)来实现,并分为2类,即维持DNA甲基化转移酶(DNMT1)和从头甲基化酶[22-24]。随着对DNA甲基化的继续深入研究,发现一些可以抑制甲基化酶的药物,甲基化酶抑制剂是一种嘧啶核苷类似物,可以致使多种抑癌基因重新激活恢复功能,重新发挥抗肿瘤的作用,以及诱导细胞凋亡,抑制肿瘤细胞的生长,因而有其广阔的临床应用前景,应用较广泛的是甲基化抑制剂5-氮杂-2甲基脱氧胞苷(5-Aza-dC)。利用药物手段调节DNA甲基化水平可以帮助了解基因甲基化对其的影响。
细胞凋亡是细胞的一种基本生物学现象,在多细胞生物去除冗余或异常细胞中起着必要的作用,在生物体进化、内环境稳定以及多个系统发育中起着重要作用[25-27]。Kiianitsa等[28]发现,PARP1在5-Aza-dC处理的人成纤维细胞中形成共价DNA加合物并可以诱导凋亡,阻断DNA复制。Shire等[29]发现,5-Aza-dC恢复了SULF1阴性细胞系中SULF1 mRNA的表达,与此相关的是硫酸酯酶活性的增加和促进肝癌细胞凋亡。李瑾[30]研究发现,5-Aza-dC通过影响DNA甲基化水平,引起细胞周期阻滞,使细胞发生凋亡从而具有抗肿瘤作用。
吴元等[31]用5-Aza-dc处理T24细胞,结果发现,Bcl2蛋白表达降低,Bax、Cleaved Caspase-3表达上升,但不显著。Kiianitsa等[32]发现,在5-Aza-dC处理的人成纤维细胞中PARP1形成共价的DNA加合物并诱导细胞凋亡。而本试验在牛成肌细胞中添加不同浓度的甲基化转移酶抑制剂5-Aza-dC后,用CCK8检测0、24、48 h细胞增殖情况。结果显示,0.1 μmol·L-15-Aza-dC处理组是本试验组中最适浓度并用于后续试验,通过流式细胞仪对细胞凋亡和细胞周期进行检测,发现0.1 μmol·L-15-Aza-dC处理组能够极显著促进成肌细胞凋亡(P<0.01);对细胞凋亡因子检测表明,该浓度极显著地降低细胞抗凋亡因子Bcl的表达(P<0.01),极显著地提高了促凋亡因子Bax表达(P<0.01),显著提高了促凋亡因子Caspase-9的表达(P<0.05);这表明5-Aza-dC能够极显著促进关岭牛成肌细胞的凋亡。同时对细胞周期的检测发现,5-Aza-dC对细胞周期的影响差异不显著(P>0.05),为进一步验证5-Aza-dC对细胞周期的影响,通过qRT-PCR发现,周期因子CyclinA2表达显著提高(P<0.05),而细胞因子CyclinB1、CyclinD表达未发生显著变化(P>0.05)。李凤珍[33]研究发现,低浓度范围的5-Aza-dC(0.005 μmol·L-1)不会显著的影响细胞周期。李瑾[30]研究发现,10 μmol·L-1的5-Aza-dC处理人肺癌细胞A549 48 h后,人肺癌细胞A549细胞周期阻滞于G2/M期,且细胞内的活性氧水平显著升高(P<0.05)。本试验发现,使用0.1 μmol·L-15-Aza-dC处理后,周期因子CyclinA2的表达显著提高(P<0.05),推测可能是由于5-Aza-dC浓度偏高。细胞凋亡和细胞周期都是维持体内细胞数量动态平衡的基本措施以及维持胚胎正常发育、机体健康的基本生物学现象[34-37]。本研究发现,5-Aza-dC能够促进细胞凋亡,但是对于细胞周期影响不明显,对于5-Aza-dC的最适浓度和时间还要进一步验证。
MyoD1基因在调节肌肉细胞的生长、代谢及促进肌肉细胞的增殖、分化等功能上均具有的重要作用[38]。Megiorni等[39]发现,5-Aza-dC处理后的DNA去甲基化能够上调miR-378a-3p水平,提高MyHC基因表达量。MyoD1同是重要的肌肉发育基因,通过0.1 μmol·L-15-Aza-dC处理细胞后发现,试验组极显著的降低MyoD1启动子区的甲基化率(P<0.01)。而空白组中MyoD1的mRNA的表达量极显著低于试验组(P<0.01),同时也发现,DNMT1的表达量极显著低于试验组(P<0.01)。可能由于5-Aza-dC的作用主要是在 DNA复制过程中能够与甲基转移酶(DNMTs)结合形成共价复合物而达到去甲基化,能够明显抑制该酶的甲基转移活性但是并不影响DNMTs表达。本研究结果提示,5-Aza-dC通过调节Caspase-9、Bcl、Bax和CyclinA2 mRNA表达水平影响牛成肌细胞的增殖和凋亡,并且低浓度水平5-Aza-dC(0.1 μmol·L-1)极显著降低了MyoD1启动子区甲基化水平。
4 结 论
本研究发现,5-Aza-dC能够通过改变Caspase-9、Bcl、Bax、CyclinA2等基因的表达来调节关岭牛成肌细胞的增殖、凋亡。检测发现,0.1 μmol·L-15-Aza-dC能极显著降低MyoD1基因启动子区甲基化水平(P<0.01),极显著提高其mRNA的表达量(P<0.01)。因此,低浓度水平5-Aza-dC(0.1 μmol·L-1)能有效降低成肌细胞中MyoD1启动子区甲基化水平进而提高MyoD1 mRNA的表达量。所以,低浓度的5-Aza-dC能通过促进细胞凋亡及调控关岭牛MyoD1的甲基化水平来调控MyoD1的表达;同时,MyoD1启动子甲基化可作为遗传标记,为后续关岭牛的品种改良提供理论依据。
