动态调水过程水文和理化因子共同驱动丹江口水库库湾浮游植物季节变化*
2021-09-23闫雪燕李玉英蒋叶青崔真真高肖飞吴乃成FohrerNicola韩雪梅
闫雪燕,张 鋆,李玉英,蒋叶青,崔真真,高肖飞,吴乃成,Fohrer Nicola,韩雪梅,4
(1:南阳师范学院水资源与环境工程学院,河南省南水北调中线水源区流域生态安全国际联合实验室,南水北调中线水源区水安全河南省协同创新中心,南阳 473061) (2:中国科学院城市环境研究所,城市环境与健康重点实验室,水生态健康研究组,厦门 361021) (3:德国基尔大学水文与水资源管理系,基尔 24118) (4:海南师范大学生命科学学院,海口 571158)
丹江口水库位于豫、鄂、陕交界的湖北省丹江口市和河南省淅川县境内,是南水北调中线工程的水源地,保障其水质安全具有重要战略意义[1]. 丹江口库区呈“V”字型,主要由汉江库区和丹江库区构成,汉库位于丹江口市,形状狭长;丹库位于淅川县,面积较大,较宽阔[2]. 每年随着水库调度运转变化,库区周边150~160 m高程形成大面积消落带. 丹库周边土地平坦,以缓坡丘陵为主,耕地占整个丹江口水库消落区耕地面积的72.5%[3],残留在耕地中的农药化肥等面源污染沉积物极易受到水体季节性侵蚀释放到水体中,从而对库区水质造成严重威胁[4].
随丹江口水库扩容、水位抬升、蓄水量增大,流域内地质和地表径流带来的外源性营养盐增加[5],加之水库运行管理(调水及供水)带来的水位波动、水力滞留时间、营养盐等水文条件和理化因子变化,均会增加水库富营养化潜在风险[6]. 以往研究多关注丹江口水库整个库体和部分支流的水质状况[7-8],对库湾水质和水生态的研究相对欠缺. 库湾是水库型湖泊水质安全的敏感区[9],污染物易在此区域滞留和富集. 随着自然和人为因素导致的污染物不断积累,库湾发生富营养化的风险增高,进而可能导致湖库整体水质恶化[10]. 受季节变化和水库调度运行影响,库湾的水位和水体交换能力等也会发生周期性变化. 因此,研究动态调水过程丹库库湾水质状况,有利于监测丹江口水库水质,预防水库水体富营养化,保障水库水生态安全.
浮游植物是水生态系统的重要初级生产者,群落结构和功能与水质密切相关[11-12],其丰度和组成随营养盐、温度、光照[13]、水文条件[14-15]等改变而变化,常被用作水体健康状况指示生物[16]. 有关湖泊的长期研究表明,浮游植物特定优势种组合常在固定时段重复出现,这一过程常受水体物理、化学、水文和生物等因子调控[17-18]. 溶解氧、pH值、总磷(TP)和水温是丹江口水库浮游植物发生演替的关键因子[19],而在镜泊湖中,水位波动、透光深度及TP与蓝藻生物量密切相关[20]. 水位升高会促进蓝藻的生长[21],入库流量减少和温度升高可能是导致美国水库有害藻类异常增殖的原因[22]. 尽管前人已经开展了许多浮游植物群落动态变化及驱动因素的研究,但丹江口水库作为主要水源地,具有其个性特征,如水位波动大、水体更新快等特点[23]. 因此,了解丹库库湾浮游植物群落季节变化规律,掌握引起浮游植物群落结构转变的环境因子,实施“一库一策”,这对于动态调水过程丹江口水库监测和有效管理具有重要价值. 本研究重点探讨了南水北调中线工程调水背景下丹库区域库湾浮游植物的群落特征,尤其是浮游植物季节演替规律及其与水文和理化因子的关系,为南水北调中线工程水源地丹江口水库的环境保护与科学管理提供重要理论支撑.
1 材料与方法
1.1 样点概况
丹江口水库位于北亚热带向温暖带的过渡带,此区域年平均降水量743.5 mm,年内分配极不均匀,每年7-9月降水量较大. 2012年丹江口大坝加高后,常年蓄水位介于157~170 m之间,年调水量达130×108m3,有效调水量为95×108m3,水库扩容后库容增加至290.5×108m3[24]. 丹库周边土地为缓坡丘陵,土地利用方式以耕地为主.
依据动态调水过程丹库水流方向,从丹库主要供水支流丹江入库口黑鸡嘴(HJZ)到南水北调中线干渠陶岔调水口渠首(QS),共设置18个采样点(图1),除黑鸡嘴和渠首两个样点外,其余16个样点分别位于丹库库心(KX)和3个库湾. 3个库湾均位于丹库东岸,以往周边消落地以农田为主,每个库湾设置5个样点. 自北向南,依此为胡寨(HZ1~HZ5)、宋岗(SG1~SG5)和全店(QD1~QD5). 其中,胡寨位于水库准保护区范围内,在此区域有养鱼现象,受人类活动干扰和面源污染影响较大,平水期与库区隔离,形成独立区域;丰水期与库区相通,与丹库形成一个整体. 宋岗位于二级保护区内,是重要客运码头,人类活动干扰频繁. 全店位于一级保护区域内,受人类活动干扰相对较少.

图1 丹库样点分布Fig.1 Distribution of the sampling sites in Danjiang Section, Danjiangkou Reservoir
1.2 样品采集与测定
分别于2018年7、10月和2019年1、5、7月进行5次水样和浮游植物样品的采集. 柱状采水器采集0~50 cm表层水样2 L,装入聚乙烯采样瓶后,低温避光保存,用于水质测定. 柱状采水器另采集表层水2 L,加入30 mL鲁哥试剂固定,室内经分液漏斗静置沉淀48 h,吸去上清液,保留30~50 mL浓缩样品,定量分析浮游植物群落组成.
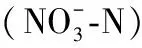
1.3 数据处理
物种丰富度指数(S)、Shannon-Wiener多样性指数(H′)和Mcnaughton优势度指数(y)用于表征藻类的多样性特征:S为物种数;H′=-∑PilnPi[31];y=fi(ni/N),当y>0.02时,定为优势种[32]. 式中,Pi=ni/N,Pi为第i个物种个体数量与所有个体总数量的比值,ni为第i个物种的个体总数,N为所有物种的数量,fi为第i种在各点位的出现频率.
采用单因素或多因素ANOVA探讨季节或样点间环境因子和浮游植物群落的差异,采用Pearson相关性分析浮游植物丰度、多样性指数和环境因子的相关关系,相关统计分析在IBM SPSS Statistics 23软件中完成. 主成分分析(principal component analysis,PCA)用于分析基于环境因子的样点分布特征,典范对应分析(canonical correspondence analysis, CCA)用于分析样点浮游植物群落分布与环境因子之间的相互关系[33],邻体矩阵主坐标分析(principal coordinates of neighbor matrices,PCNM)用于分析样点间的空间关系. 变差分解、PCA和CCA分析主要基于R3.6.3软件vegan包完成,(偏)Mantel相关分析基于R3.6.3软件ggcor包完成,样点分布地图基于ArcGIS 10.2 (ESRI, Redlands, CA, USA)完成.
2 结果
2.1 水文与理化因子分析
2018年7月-2019年7月,丹江口水库全年平均水位156.64 m,水位季节性波动明显,9月(秋季)水位达到最高,4月(春季)水位最低(图2a). 调水口流速全年平均值为0.76 m/s,流量平均值为203.48 m3/h (图2b). 流速与流量变化趋势基本一致,秋季丰水期达到最高,冬季平水期最低,与丹江口水库水位变化并不完全一致.
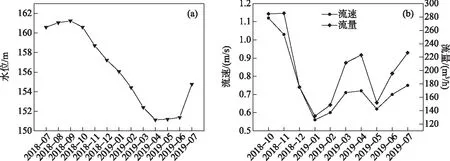
图2 丹库水位与调水口流速和流量的变化Fig.2 Changes of water level, water flow rate and velocity in Danjiang Section, Danjiangkou Reservoir
不同季节各样点主要环境因子经标准化处理后,进行PCA排序分析(图3). PCA分析结果显示,PC1轴解释度为19.04%,PC2轴解释度为18.46%,不同季节样点有聚集现象,但差异不明显;不同类型样点间聚集现象不明显,但发现在夏季,入库口黑鸡嘴、库心、调水口渠首与库湾样点区分明显,而库湾间差异较小;秋季,黑鸡嘴与库湾、库心和渠首样点的理化因子间存在差异.

图3 丹库不同季节样点环境因子PCA排序Fig.3 PCA of environmental variables in different seasons in Danjiang Section, Danjiangkou Reservoir


表1 丹库水体主要理化因子参数*
4个季节水体理化因子变化情况见表2. 水温、ORP、Cond、TN和TP浓度均呈现明显的季节变化规律,秋季ORP最高,夏季ORP最低. Cond与之相反,夏季最大,秋季最小. TN浓度在秋季达到最高值,其他3个季节水平一致. TP浓度则是冬季最高,春、夏季最低,其他指标4个季节无显著差异.
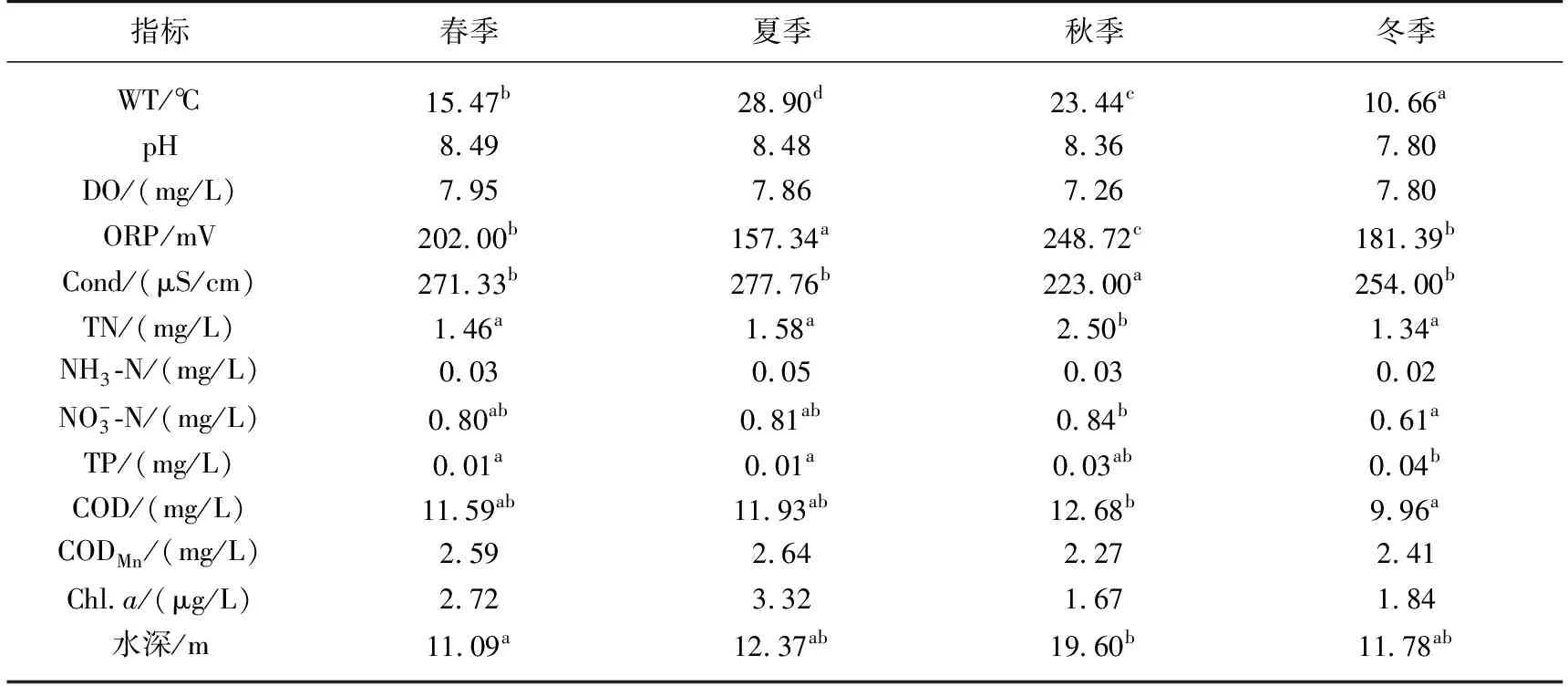
表2 丹库不同季节水体理化因子
2.2 浮游植物群落组成特征
本研究共鉴定出浮游植物7门68属92种,其中绿藻门种类最多(35种),占浮游植物总种类数的38.04%;其次为硅藻门(29种),占31.52%;蓝藻门13种,占15.29%;隐藻门和裸藻门均为5种,分别占5.90%;甲藻门4种,占4.70%;金藻门仅发现1种. 浮游植物总丰度全年变化范围为0.43×103~4.7×106cells/L,平均丰度为1.94×105cells/L. 浮游植物季节差异性明显(图4),春季硅藻和绿藻为主要优势门,夏季大部分样点绿藻代替硅藻成为第一优势门,蓝藻在各库湾所占比例上升;秋季蓝藻成为库湾的绝对优势类群,而其他3个样点依然是绿藻为第一优势门;进入冬季后,硅藻比例上升,浮游植物组成又呈现多样化特征,但渠首蓝藻比例依然较高.
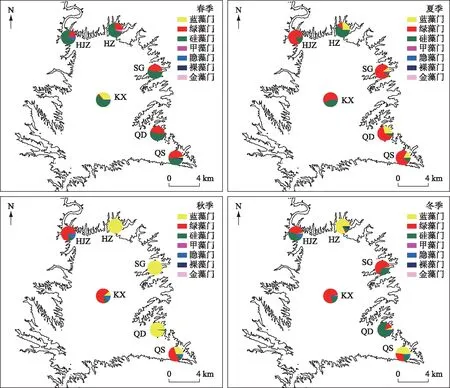
图4 丹库不同季节各样点浮游植物相对丰度 (HJZ:黑鸡嘴;HZ:胡寨;SG:宋岗;QD:全店;KX:库心;QS:渠首)Fig.4 Phytoplankton relative abundance in different seasons in Danjiang Section, Danjiangkou Reservoir
18个样点共筛选出34个浮游植物优势物种(y>0.02). 春季,16个优势物种主要隶属于硅藻门和绿藻门,其中硅藻门小环藻(Cyclotellasp.)的优势度最高(0.281),黑鸡嘴与库心的优势类群相似,库湾全店和胡寨与渠首的优势类群相似,库湾宋岗的优势类群与其他样点差异较大. 夏季,21个优势物种主要属于硅藻门、绿藻门和蓝藻门,其中硅藻门颗粒直链藻极狭变种(Melosiragranulatavar.angustissima)的优势度最大(0.384),黑鸡嘴与库心的优势类群相似,库湾胡寨、宋岗和全店的优势类群相似,渠首的优势类群与其他样点差异较大. 秋季,12个优势物种主要属于硅藻门和蓝藻门,依然是硅藻门的颗粒直链藻极狭变种优势度最大(0.289),黑鸡嘴、库心、宋岗和全店的优势类群相似,胡寨和渠首与这4个样点差异显著. 冬季,13个优势物种主要属于硅藻门,硅藻门的颗粒直链藻(Melosiragranulata)优势度最大(0.236),全店、宋岗和黑鸡嘴优势类群相似,渠首、库心和胡寨与其有较大差异.
浮游植物多样性指数双因素方差分析表明,总丰度、物种数和Shannon-Wiener指数在季节、地点和两者交互作用上均存在极显著差异(表3,P<0.01). 物种数变化范围为8~16,Shannon-Wiener指数为1.064~1.698. Tukey多重比较表明,总丰度夏季最高,秋季最低;物种数夏季最大,秋冬季最小;Shannon-Wiener指数春季最高,秋季最低,从春季到秋季持续降低,冬季有所上升(表4). 春季,库心与黑鸡嘴Shannon-Wiener指数相似,其他样点Shannon-Wiener指数均升高;夏季,与入库口黑鸡嘴相比,仅库湾胡寨Shannon-Wiener指数降低;但秋季,这种差异不明显;冬季,与其他样点相比,渠首Shannon-Wiener指数升至最高.
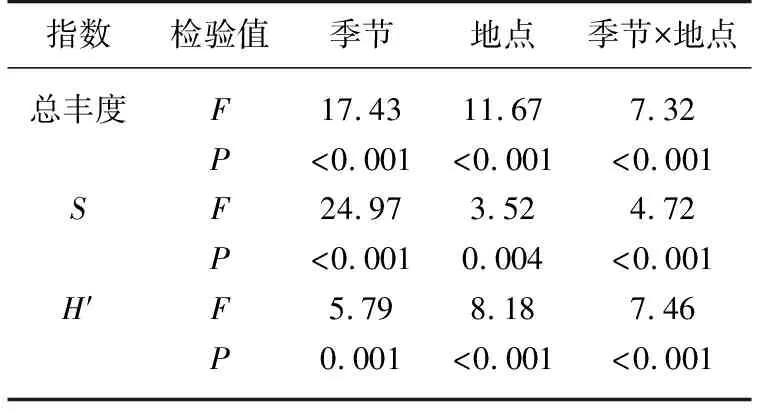
表3 丹库多样性指数主体间效应检验结果
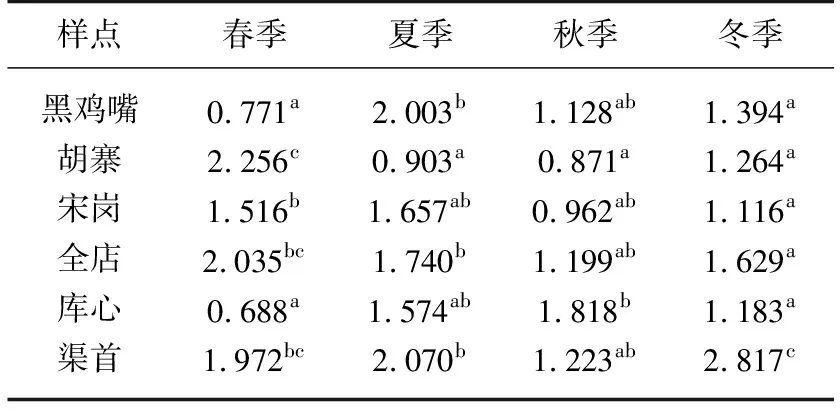
表4 不同季节丹库不同样点间浮游植物 Shannon-Wiener指数的差异
2.3 水文因子对浮游植物群落结构的影响
为了进一步了解调水过程水文因子对浮游植物群落组成的影响,将优势门丰度和优势物种丰度与丹江口水库水位和调水口渠首流速和流量进行Pearson相关性分析(图5). 蓝藻门丰度与流速、流量呈极显著正相关,其中蓝藻门的优势种鱼腥藻、微囊藻、水华束丝藻丰度均与水位、流速和流量呈极显著正相关. 硅藻门丰度与水位和流速呈极显著负相关,与流量呈显著负相关,硅藻门的针杆藻(Synedrasp.)、菱形藻(Nitzschiasp.)、肘状针杆藻(Synedraulna)和小环藻丰度均与水位呈显著负相关,梅尼小环藻(Cyclotellameneghiniana)丰度与流量呈显著负相关. 绿藻门丰度与这3项水文因子无显著相关性,但绿藻门的空球藻丰度与水位呈显著负相关. 将浮游植物Shannon-Wiener指数与水位、流速和流量进行线性回归分析表明,Shannon-Wiener指数与水位、流速和流量呈极显著负相关(图6),但R2值均较小,水位对Shannon-Wiener指数的影响最为明显,流量影响最小.

图5 丹库浮游植物优势物种丰度与环境因子的相关性热图Fig.5 Heatmap of correlation between abundance of phytoplankton dominant species and environmental parameters in Danjiang Section, Danjiangkou Reservoir

图6 丹库浮游植物Shannon-Wiener指数与水文因子的线性回归分析Fig.6 Linear regression of Shannon-Wiener indices of phytoplankton and hydrological factors in Danjiang Section, Danjiangkou Reservoir
2.4 理化因子对浮游植物群落结构的影响
为掌握不同类型环境因素对丹库浮游植物群落组成的影响,对季节、地点、理化因子(含水深)和PCNM进行方差分解(图7). 这4类生态因子共解释了浮游植物群落组成变化的39.0%,其中理化因子的解释率最高,达到23%;其次为季节因子,解释率为17%;地点差异的解释率为12%;PCNM解释率最低,仅为9%. 季节与理化因子共同解释率为9.0%. 地点、理化因子和PCNM三者共同解释率为6.0%,说明样点空间格局导致理化因子差异,进而影响到浮游植物群落分布.

图7 丹库浮游植物群落结构方差分解 Fig.7 Variation partitioning of phytoplankton community structure in Danjiang Section, Danjiangkou Reservoir
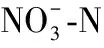

表5 丹库不同样点浮游植物群落与理化因子的偏Mantel系数1)

图8 丹库浮游植物群落与理化因子的CCA排序Fig.8 CCA of phytoplankton communities and physico-chemical factors in Danjiang Section, Danjiangkou Reservoir


图9 丹库理化因子与浮游植物群落不同门物种丰度的相关分析Fig.9 Physico-chemical variables and abundance of phytoplankton phyla composition in Danjiang Section, Danjiangkou Reservoir
将浮游植物优势物种丰度与理化因子进行Pearson相关性分析(图5)发现,蓝藻门的水华束丝藻(Aphanizomenonflosaquae)和微囊藻(Microcystissp.)丰度与ORP呈显著正相关,束丝藻(Aphanizomenonsp.)丰度与ORP呈显著负相关. 硅藻门的颗粒直链藻极狭变种丰度(r=0.82)和脆杆藻(Fragilariasp.)丰度(r=0.70)均与Chl.a浓度呈极显著正相关,颗粒直链藻丰度与COD和水温呈显著负相关,梅尼小环藻和等片藻(Diatomasp.)丰度均与水温呈显著负相关,小环藻丰度与水深呈极显著正相关. 绿藻门的单角盘星藻(Pediastrumsimplex)丰度与ORP呈极显著负相关,卵囊藻(Oocystissp.)和栅藻丰度均与水深呈极显著正相关,双对栅藻(Scenedesmusbijugatus)和衣藻(Chlamydomonassp.)丰度均与水深呈显著正相关,衣藻丰度还与DO浓度呈显著正相关. 隐藻门的嗤蚀隐藻(Cryptomonaserosa)丰度与DO浓度呈显著正相关.
3 讨论
3.1 丹库浮游植物季节变化特征
丹江口水库丹库浮游植物群落季节差异明显. 春季硅藻和绿藻为优势类群,水温是影响藻类群落组成的重要因素[34],在横山水库[35]、香溪河水库[36]及大溪水库[37]等水体中均发现水温能够影响浮游植物群落结构. 适宜温度可以提高藻类的光合作用,加快细胞新陈代谢,促进其生长繁殖[38]. 硅藻门丰度与水温呈正相关,春季有利于硅藻丰度增加.但硅藻门的优势物种颗粒直链藻、梅尼小环藻、等片藻丰度却与水温呈显著负相关,说明适宜的温度(10~25℃)有利于硅藻生长,但温度的过分升高反而会抑制硅藻的丰度. 因此,研究发现,进入夏季后硅藻门的优势地位丧失. 丹江口水库春季水位最低,硅藻门的针杆藻、菱形藻、肘状针杆藻和小环藻丰度均与水位呈显著负相关,说明低水位的扰动有利于硅藻门某些藻类的生长繁殖. 夏季水温升高,光照增强,绿藻依然为优势类群,但硅藻优势地位下降,蓝藻比例上升. 较高温度能促进藻类的生长繁殖,并使蓝藻成为优势类群[39],蓝藻优势形成是其自身生理特征以及营养盐、温度、光照、水文和气象条件等诸多因素综合作用的结果[40]. 进入秋季,优势类群为蓝藻,这是因为这个时期丹江口水库进入丰水期,伴随降雨量的增加,进入库内的面源污染物增加,为蓝藻繁殖提供了丰富的营养盐. 从水质结果也可以看出,秋季水体中ORP、TN、TP和COD浓度均高于其他季节. 也有研究指出,营养化水平较低水体以硅藻型浮游植物群落为主,而营养化水平较高水体以蓝-绿藻型群落为主[41],蓝藻门的大量繁殖与TP浓度等营养物密切相关[42]. 冬季,随着水位的下降,进入水体中的营养盐和有机物数量降低,蓝藻和绿藻比例下降,硅藻比例上升,蓝藻、绿藻和硅藻成为这个季节的共同优势物种.
与其他学者的研究结果对比发现,调水前后丹江口水库浮游植物群落组成的季节变化差异明显. 申恒伦等[11]指出2007-2008年春季丹江口水库浮游植物以硅藻-蓝藻型为主,夏、秋季演替为隐藻-蓝藻型,冬季发展为硅藻-隐藻-甲藻型,春季和秋季硅藻所占比例最大;谭香等[7]在2009-2010年的调查发现,春、秋和冬季硅藻在数量上均占绝对优势,夏季蓝藻为优势门. 王英华等[19]发现,2014-2015年春季和冬季硅藻均为丹江口水库优势类群,但夏季绿藻占优势. 本研究结果与王英华等的研究结果更为相像,笔者推测,这可能是因为2012年丹江口大坝水库扩容,影响了丹江口水库的物质交流和能量转化,从而改变了浮游植物群落季节动态. 水文调控导致的湖泊高水位运行和水动力减弱,会使湖泊沿岸带生境逐渐丧失,湖泊内有机碳和营养盐滞留效应增强,促进了浮游生物的生长[43]. 冯宇墨等也发现好汉泊水库扩容后,浮游植物丰度和生物量均有所增加,其中绿藻门和蓝藻门上升最为明显[44].
3.2 调水影响丹库浮游植物群落组成
在秋季高水位期,调水口具有较高的流速和流量,浮游植物群落多样性较低,Shannon-Wiener指数与水位、调水口流速和流量呈显著负相关,其中与水位的关系更为密切. 由此可以看出,水文因子波动引起的强扰动会抑制某些浮游植物的生长[45]. 李哲等发现,流量与降雨作为主要的物理扰动因子,引起澎溪河浮游植物多样性变化. 蓄水期水位上升、流量骤降,水体扰动强度加剧导致藻类多样性下降[46],这与笔者的研究结果相一致. 张毅敏等也发现,流速和风力等因子扰动较强时,会降低藻类丰度[47]. 因此,秋季调水口较低的浮游植物多样性与该季节较高调水量带来的强扰动有关. 水动力作为水生态系统中的主要驱动力,会直接或间接地引起其他环境条件的改变,从而影响浮游植物的生长繁殖,进而影响更高级营养级,甚至整个生态系统[48].
3.3 丹库库湾浮游植物群落特征
库湾浮游植物群落结构与入库口黑鸡嘴、库心和调水口渠首区分明显. 调水口浮游植物群落组成受入库口和库湾群落的共同影响,库湾水体富营养化程度直接影响到调水的质量. 库湾是水库中受人类活动干扰最强烈的区域,水质产生明显变化,进而影响到浮游植物的群落结构[49],夏季和秋季这种效应最为明显. 夏季宋岗和全店两个库湾的浮游植物均为绿藻-蓝藻型,进入秋季后,3个库湾的浮游植物几乎完全由蓝藻构成,而黑鸡嘴、库心和渠首3个样点则由绿藻和蓝藻共同构成. 研究指出,随着水体富营养化的发展,水体中浮游植物群落逐渐从以硅藻和绿藻为主演替为以蓝藻为主[50-54],蓝藻门的微囊藻、鱼腥藻、颤藻和束丝藻为优势种[55-56],这些藻类也是本次研究发现的优势类群. 蓝藻是水体富营养化引起水华的主要藻类,浅水湖泊和水产养殖池塘的水体易暴发蓝藻[57]. 库湾受人类扰动强烈,在夏季和秋季有发生水体富营养化的风险. 而且笔者也发现,夏、秋季浮游植物物种多样性指数均下降,物种多样性的降低意味着生态系统结构的简单化,进而影响到生态系统的生产力和稳定性.
库湾内部样点间浮游植物群落结构存在差异. 尽管夏季库湾胡寨蓝藻相对丰度较低,但从绝对丰度来看,夏、秋季,胡寨蓝藻丰度远远高于其他两个库湾,藻类总丰度也最高. 胡寨位置受人类活动干扰较多(附近有渔业养殖),养殖活动增加了水体中有机物和营养盐的浓度,提高了蓝藻的丰度. 研究结果也显示,与其他两个库湾相比,胡寨水体Cond、TN、TP和Chl.a浓度最高,这与笔者的推测相一致. 同时,胡寨周边主要为农田丘陵,夏、秋季降雨频繁,降雨量大,多种污染负荷物随雨水流入河道和湖泊,导致较多有机污染物和营养盐排入库湾,造成胡寨夏、秋季浮游植物丰度最高.
4 结论
丹江口水库丹库浮游植物群落组成以硅藻、绿藻、蓝藻和隐藻为主,春、冬季物种多样性高,群落结构复杂,夏、秋季绿藻和蓝藻比例高. 动态调水过程,秋季库区高水位、调水口高流速和高流量的扰动,会引起浮游植物多样性下降. 水文因子和理化因子共同影响了浮游植物群落组成动态变化,水温和营养盐仍然是决定浮游植物群落结构的主控因素,同时水位和流速也是影响丹江口水库浮游植物群落结构的主要驱动因子. 人类干扰强度变化会改变库湾水质理化性质,强干扰有利于提高蓝藻门的相对丰度,增加丹江口水库发生富营养化的风险. 本文关于水质、藻类和水文的复杂关系研究为丹江口水库的保护和管理提供了理论支持.
