内麦系列小麦品种(系)的染色体结构变异分析
2021-09-23关淑仙黄辉跃汪仁全王相权王仕林荣飞雪杨杰智钟光跃周海燕陈新媛黄书盈
关淑仙,黄辉跃,汪仁全,王相权,王仕林,荣飞雪,杨杰智, 钟光跃,周海燕,陈新媛,李 明,黄书盈
(四川省内江市农业科学院,四川内江 641000)
由于大多数育种单位多年的定向育种,导致小麦的遗传基础日益狭窄,抵御风险的能力逐渐下降[1-2]。评价材料的遗传多样性有利于小麦育种亲本的选择,从而避免使用同一骨干亲本。在遗传分析中,分子标记是常用的工具。这些分子标记能方便和快捷地跟踪一些优良性状/基因,但是分子标记不能反映染色体区段的具体结构变异。而植物染色体核型分析是研究植物染色体数量及结构变异、形态结构特征和物种起源与进化关系的重要方法,在植物育种中具有重要应用价值[3-6]。近年来,在植物染色体核型分析中越来越多的研究采用某些克隆重复序列或寡核苷酸序列为探针,利用荧光原位杂交技术(FISH)技术建立FISH核型图。FISH技术是一种根据碱基互补配对原则,使带有荧光物质的探针与目标DNA接合,最后用荧光显微镜可直接观察目标DNA所在的位置。近几年利用FISH技术,在小麦染色体结构变异和外源染色体识别方面已经有很多报道[7-8]。利用寡核苷酸探针和FISH技术,可以反映不同小麦品种(系)的染色体结构差异[9]。Tang等[10]利用寡核苷酸序列探针pTa535和pSc119.2,清晰地识别了小麦A、B和D基因组染色体,并建立了中国春的标准FISH核型图。Huang等[9 ]利用一系列寡核苷酸序列探针,对373个栽培品种及骨干亲本进行FISH分析,发现了大量的多态类型和结构变异,并根据FISH核型对其进行分类。蒽 玮等[11]通过对南麦号系列小麦品种进行FISH核型分析,明确了南麦号系列小麦品种及其亲本的染色体结构特点。因此,利用基于寡核苷酸探针的FISH技术,可以快速、方便地分析小麦亲本及其衍生系的染色体结构差异,进而跟踪亲本染色体结构在衍生系中的变异情况。内麦系列品种(系)是由四川省内江市农业科学院选育的高产且高抗条锈病品种(系),连续多年遴选为四川省及全国主导品种,在长江上游区广泛种植,取得显著的社会经济效益。本研究利用内麦号系列品种(系)进行染色体核型分析,研究这些材料的染色体结构变异情况,以期为小麦育种提供更为直观的遗传变异参考。
1 材料与方法
1.1 材 料
21份普通小麦(AABBDD,2n=42)材料(表1),均由四川省内江市农业科学院选育。中国春(CS)作为FISH核型的对照。
1.2 方 法
每份材料随机选取5粒种子,置于湿润滤纸的培养皿中,4 ℃过夜(约24 h),露白后转移到 23 ℃恒温培养箱中。待根长达到1~2 cm时剪取,用一氧化二氮密封处理4 h,之后用乙酸固定5~10 min,70%乙醇保存[13]。取根尖2~3 mm部分,在纤维素酶/果胶酶溶液(2∶1,w/w)中酶解,得到的悬浮液滴到载玻片上,于显微镜下镜检并拍照[12]。
荧光原位杂交过程参照Hao等[13]的方法进行,所有材料的FISH核型分析参照Tang等[9]的标准。利用TAMRA(6-carboxytetramethylrhodamine)和6-FAM(6-carboxy-fluorescein)分别标记的寡核苷酸pTa535和pSc119.2作为探针(pTa535和pSc119.2探针能清晰识别普通小麦A、B和D基因组所有染色体),对所有材料进行FISH核型分析。pTa535和pSc119.2两种重复序列最初分别来源于普通小麦和黑麦。pTa535和pSc119.2两种寡核苷酸探针均由上海生工生物工程股份有限公司合成。
2 结果与分析
2.1 21份小麦材料和中国春的FISH分析
利用pSc119.2和pTa535探针对21份材料和中国春根尖细胞进行原位杂交分析,建立了FISH标准核型图(图1)。从pSc119.2的FISH带型看,有5份材料(内麦482、内麦0821、内麦919、内麦071和内麦854)含1RS/1BL易位染色体。这5份1RS/1BL易位系材料在之间的试验中高抗条锈病,为今后培育高抗条锈病材料提供了新的育种资源。
所有材料的B基因组染色体以及2D和4D染色体上都有pSc119.2信号。A基因组染色体上的pSc119.2信号有7种类型:1、1A、2A、5A和6A染色体上均有pSc119.2信号(内麦366); 2、4A、5A和6A染色体上均有pSc119.2信号(内麦9号、杏麦2号、靖麦19、内麦2889、内麦836和内麦316);3、5A和6A染色体上均有pSc119.2信号(内麦5348); 4、4A和6A染色体上均有pSc119.2信号(内麦482);5、4A和5A染色体上均有pSc119.2信号(中国春、内麦561、内麦919、内麦071和内麦673); 6、仅5A染色体上有pSc119.2信号(内麦416、内麦866和内麦101);7、仅4A染色体上有pSc119.2信号(内麦0821、内麦538、内麦7538、内麦854和内麦5348)。以上结果说明,A基因组染色体遗传变异较丰富,尤其是在5A染色体上。在D基因组中,pSc119.2信号还出现在1D(内麦673和内麦101) 和3D(内麦482、内麦0821、内麦538、内麦7538、内麦854、内麦510、内麦561、内麦919、内麦071、内麦673、内麦416和内麦866)染色体上。pSc119.2信号在1D、2D、3D和4D染色体的变异类型只有1~2种(图2),说明D基因组染色体遗传变异低。所有B染色体上都有pSc119.2信号,但变异类型也只有1~2种(5B和6B染色体除外)(图2),说明B基因组染色体遗传变异较低。pTa535在所有材料的D基因组染色体、除5A外的其他A基因组染色体以及3B、6B和7B染色体上分别有强、较强和弱信号。同时,在中国春和内麦101的5A染色体长臂上有较强的pTa535信号。pSc119.2和pTa535探针信号表明,A基因组染色体有较高的变异,而B和D基因组染色体有较低的变异。所以在今后的育种过程中,选择杂交亲本时要注意B和D基因组染色体的遗传变异,这样才能丰富育成品种的遗传多样性。

表1 21份材料的来源及审定情况Table 1 Source and approval status of the 21 materials
2.2 21份小麦材料的FISH核型多样性分析
在21份小麦材料中共鉴定出49种染色体多态类型(图2)。A基因组染色体多态类型数量最多(22种),其次是B基因组(14种)和D基因组(13种)。所有材料每条染色体上多态类型出现频率最高的被认定为类型1(5A除外),其频率范围为29%~100%,平均约83%。类型1是每条染色体上最占优势的类型。每条染色体上多态类型频率次于类型1的被认定为类型2,频率为 5%~38%,平均约19%。类型2是每个染色体上第二常见的类型,有一定的选择优势。每条染色体除类型1和类型2外,其他多态类型频率为5%~19%,平均约8%。这些较低频率的染色体多态类型可能在适应特定环境的品种中具有 优势。
在A基因组染色体中,5A和6A染色体分别有7种和4种变异类型,而其余染色体都不超过3种。在B基因组染色体中,5B染色体有4种变异类型,其余染色体都不超过3种。在D基因组染色体中,变异类型最高的是3D染色体(3种),其余染色体的变异类型为1~2种(图2)。以上结果表明,不同染色体的变异类型数量不一致,推测在育种过程中变异类型数量多的染色体在不断进化,而没有变异或者变异少的染色体比较保守。5B、6B、3D和7D染色体的同源染色体之间存在不对称核型(图2)。pSc119.2信号分别在一条5B染色体的长臂和一条6B染色体的短臂上消失,pTa535信号分别在一条3D染色体的长臂和一条7D染色体的短臂上消失。以上染色体的不对称核型可能是由染色体结构变异引起。
2.3 21份小麦材料的FISH核型分类
根据A、B和D基因组染色体的FISH核型类型,把21份材料分为16类(表2)。其中内麦538、内麦7538和内麦510的FISH核型一致;靖麦19、内麦2889和内麦836的FISH核型一致;内麦919和内麦071的FISH核型一致。其他材料的FISH核型都不一致。内麦538、内麦7538与内麦510的亲本不同,但其FISH核型一致,这说明它们的杂交亲本的遗传背景可能相似。内麦5348与内麦101的亲本相同,但其FISH核型却不一致,说明亲本一样的衍生系后代在染色体结构上可能会发生变异(表1和表2)。
3 讨 论
3.1 染色体FISH类型的多态性
构建植物染色体FISH核型图,在植物的分类、进化以及育种研究中具有重要意义[14-15]。Huang等[9]根据373个小麦栽培品种及骨干亲本的高清FISH核型,鉴定出167种染色体多态类型。同时在148个品种中发现14种染色体结构变异类型,包括易位、倒位和缺失等。在染色体多态类型方面,本研究结果与Huang等[9]结果一致,多态类型的数量都是A基因组>B基因组>D基因组。本研究只发现49种染色体多态类型,可能是研究材料比较少的原因造成的。本研究只发现5B、6B、3D和7D染色体上均出现核型不对称的现象。原因可能是所用探针只有pSc119.2和pTa535,而这两个探针不能识别出更多的染色体的结构变异。
3.2 FISH核型分类与遗传背景的关系
FISH核型对小麦的分类以及亲本的追溯起到重要作用[9,11]。本研究选取21份内麦系列小麦品种(系)材料,根据它们的杂交亲本了解杂交后代染色体结构变异,结果发现,杂交组合亲本相同的后代品种FISH核型不一致,可能是由于染色体结构变异造成的[11];同时也发现,杂交组合亲本不同的后代品种FISH核型一致,可能是亲本FISH核型一致或者是遗传背景相似[9]。21份材料根据A、B和D基因组染色体的FISH核型将这些材料分成了16类,说明这些材料遗传多样性较高,并且一些品种(系)有自己独特的FISH核型[7]。本研究结果发现,在这16类中很多只是在少数染色体(5A、6A、5B、6B和1D等)上有信号差异,同时每条染色体多态类型出现频率最高的占比29%~100%,其他类型占比5%~38%(图 2),在育种过程中,一些染色体有选择优势,而其他染色体较保守[9]。本研究结果表明,FISH技术可以推测小麦品种系谱,反映染色体之间的交换重组关系。因此,可以建立内麦系列小麦品种(系)染色体FISH核型,从染色体结构水平来反映品种间的遗传多样性,从而推断小麦品种(系)的演变过程。
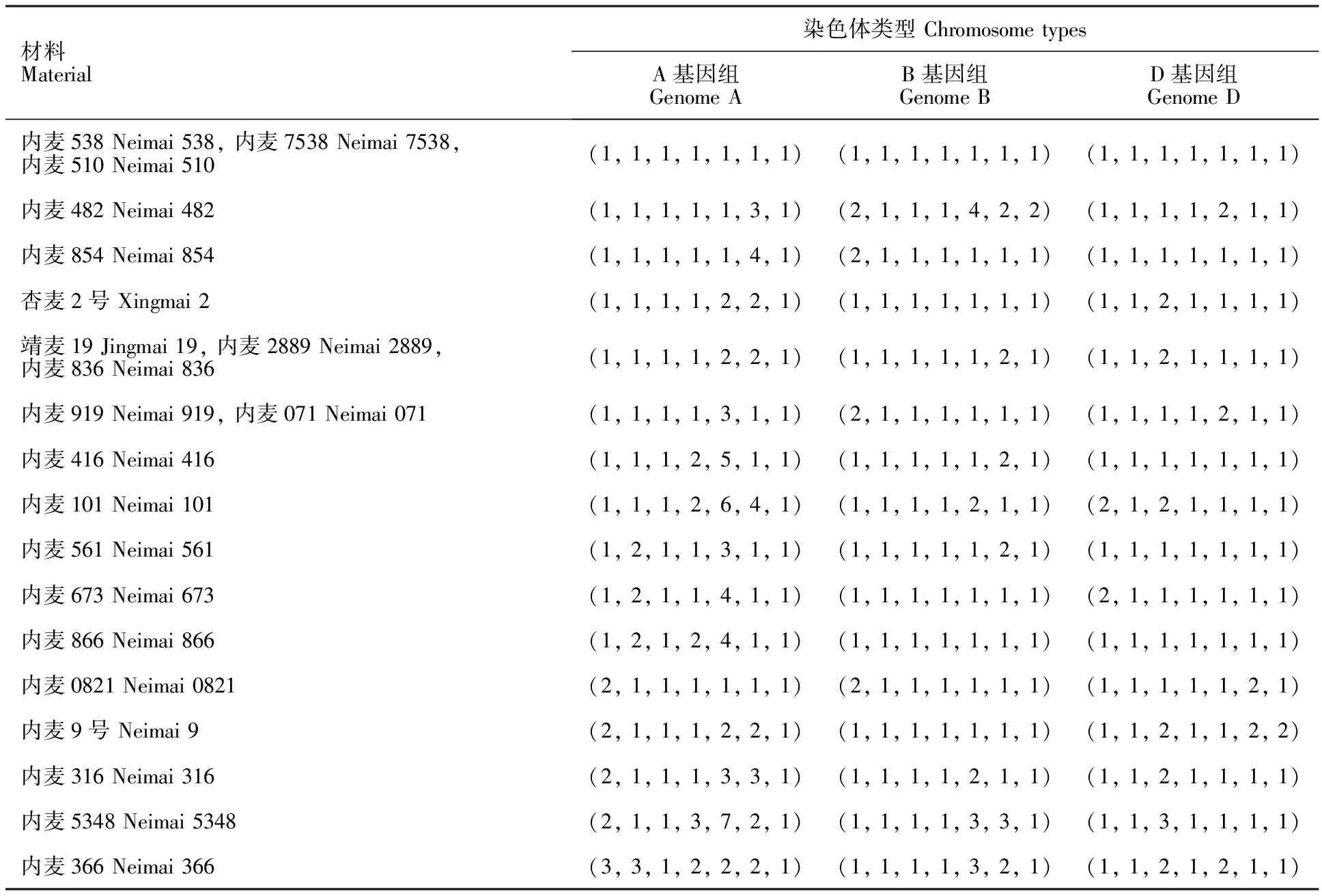
表2 21份小麦材料A、B、D基因组的FISH核型类型Table 2 FISH karyotypes of A,B and D genomes of the 21 wheat materials
综上所述,利用FISH技术可以反映小麦品种(系)的染色体结构差异,进行染色体跟踪,可以明确小麦染色体结构组成及遗传背景,为下一步的分子标记育种提供可靠的参考。
