木林子大样地两个木姜子属物种空间分布格局及关联性
2021-09-13黄小姚兰艾训儒朱江朱强王进吴漫玲刘松伯
黄小 姚兰 艾训儒 朱江 朱强 王进 吴漫玲 刘松伯
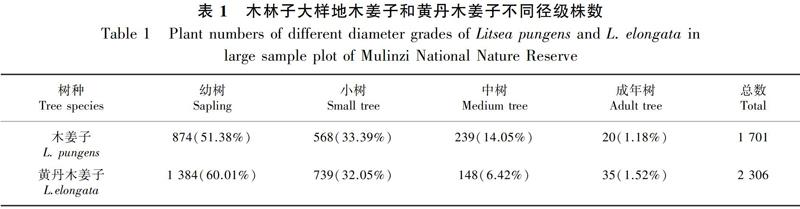

摘 要:該文以湖北木林子国家级自然保护区15 hm2大样地同属物种 [木姜子(Litsea pungens)和黄丹木姜子(L. elongata)]为研究对象,通过成对相关函数和不同零模型(完全空间随机模型、异质泊松模型和先决条件模型)的方法分析两个物种的空间分布格局、种内和种间空间关联性,从同属物种的空间分布格局角度探讨群落的构建机制。结果表明:(1)种群空间分布格局研究中,两个种的空间分布格局大致为聚集分布。基于CSR零模型,木姜子和不同径级(幼树、小树、中树和成年树)的聚集尺度分别为0~114 m、0~103 m、0~56 m、0~34 m和0~16 m,黄丹木姜子和不同径级的聚集尺度分别为0~150 m、0~150 m、0~59 m、0~56 m和0~14 m;基于HP零模型,木姜子整体的空间分布格局聚集尺度为0~23 m,其中幼树、小树和中树均在0~22 m呈现聚集分布,成年树仅在0~3 m尺度上表现为聚集分布,黄丹木姜子整体的空间分布格局聚集尺度为0~36 m、140~150 m,各径级的聚集尺度分别为0~35 m、0~35 m、0~12 m和0~6 m。(2)种内关联性研究中,木姜子和黄丹木姜子不同径级个体种内关联在小尺度上基本呈现正关联,随着尺度的增大变为微弱的无关联或负关联。(3)种间关联研究中,木姜子对黄丹木姜子和黄丹木姜子对木姜子的空间关系大致相同。总体上,两物种之间均体现为在30 m尺度内对另外一个物种呈现正关联,40~68 m尺度内呈现负关联;不同径级之间都表现为无关联,偶尔出现正关联或负关联。研究认为,种群空间分布格局基本上为聚集分布,其主要受到生境异质性和扩散限制的影响,木林子大样地中木姜子和黄丹木姜子空间分布格局存在相似性,而不同的生境偏好可能是导致两同属物种共存的原因。
关键词:同属物种,点格局,空间分布格局,空间关联,物种共存
中图分类号:Q948.15
文献标识码:A
文章编号:1000-3142(2021)08-1333-13
Abstract: In this paper, species of the same congeneric species (Litsea pungens and L. elongata ) in large sample plot of 15 hm2 in Mulinzi National Nature Reserve of Hubei Province were taken as the study object. The spatial distribution pattern, intraspecies and interspecific spatial correlations of the two species were analyzed by pairwise correlation function and different zero models(complete spatial randomness, heterogeneous Poisson and antecedent condition), and the construction mechanism of the community was discussed from the point of view of the spatial distribution pattern of the same congeneric species. The results were as follows: (1) In the study of population spatial distribution pattern, the spatial distribution pattern of the two species was generally aggregation distribution. The aggregation scales on the basis of the CSR zero model, the L. pungens and the different diameter stages(sapling, small tree, medium tree, adult tree) were 0-114 m, 0-103 m, 0-56 m, 0-34 m, and 0-16 m, respectively, and the aggregation scales of the L. elongata and the different diameter stages were 0-150 m, 0-150 m and 0-59 m, 0-56 m and 0-14 m; On the basis of the HP-zero model, the spatial distribution pattern of the whole L. pungens was 0-23 m, where in all of the sapling, small and medium trees were 0-22 m, and the aggregation distribution of adult trees was only on the scale of 0-3 m. The aggregation scale of the whole spatial distribution pattern of L. elongata was 0-36 m and 140-150 m, and the aggregation scale of each diameter grade was 0-35 m, 0-35 m, 0-12 m and 0-6 m. (2) In the study of intraspecific correlation, the intraspecies correlation of different diameter individuals showed positive correlation on a small scale, and became weakly unrelated or negative with the increase of scale. (3) In the study of interspecific correlation, the spatial relationship between the two species was about the same. On the whole, the two species showed positive correlation with another species within 30 m scale and negative correlation within 40-68 m scale, and there was no correlation between different diameter grades, and there was positive or negative correlation between the two species. It is considered that the spatial distribution pattern of the population is basically aggregation distribution, which is mainly affected by habitat heterogeneity and diffusion. The spatial distribution patterns of L. pungens and L. elongata in large sample plot are similar, and different habitat preferences may be the cause of the coexistence of the two species.
Key words: same congeneric species, point pattern, spatial distribution pattern, spatial correlation, species coexistence
一般来说,同属物种拥有相同祖先,在形态学和生理特征上都具有一定的相似性,所需的资源可能相似,从生态位分化来看,同属物种可能因为这些相似性而产生竞争,而不能稳定共存(Ackerly & Donoghue, 1998; Webb & Ackerly, 2002),同属物种共存现象多发现于热带和温带植物群落中(张健, 2009)。因此,生态学家一直对同属植物的共存进行激烈的探讨。在生态研究中对同属物种进行比较,可以最大限度地减少系统差异造成的混乱影响,从同属物种的空间结构和相关性的角度进行探讨(Swenson et al., 2006; 姚兰, 2016)。
植物种群的空间分布格局是指某一种群在一定水平内所有个体的分布位置或配置(曲仲湘, 1984; 张金屯, 1995),到现在为止是生态学研究的热点之一(Wiegleb, 1989; Nathan, 2006)。点格局方法由Ripley提出,它能够很好地解决传统格局分析方法的缺点(Condit, 2000; 张金屯和孟东平, 2004)。近些年,点格局分析越来越广泛,主要侧重于单物种的空间分布格局(范繁荣等, 2008; 黄小波等, 2018)、优势树种的空间分布格局(赵中华等, 2011; 张玉双等, 2016)、两种或两种以上物种的空间相关性(姚良锦等, 2018)以及空间分布格局与环境之间关系(刘旻霞等, 2018; 闫海冰, 2018)等研究较多,对于种群同科(何增丽等, 2017)或同属(陈俊等, 2018)研究较少。
樟科(Lauraceae)木姜子属(Litsea Lam) 的植物分布非常广泛,在中国大约有72种(陈幼竹, 2004)。亚热带常绿落叶阔叶混交林保存最为完好的地区包括木林子国家级自然保护区(黄永涛, 2016),其中木姜子属植物仅有2种:黄丹木姜子和木姜子,其数量占样地内物种总数量的4.8%。木姜子的药用价值很高,黄丹木姜子具有较高的商业价值(陈幼竹, 2004)。木姜子和黄丹木姜子的研究仅仅只限于药用价值等方面的研究,黄丹木姜子还见于叶凋落量及养分特征研究(董舒, 2016),对于更多的生态学方面的研究未见有报道,黄丹木姜子和木姜子虽不是建群种和先锋种,这类小乔木在种群空间分布格局的研究同样具有重要意义。本研究以湖北木林子大样地中同属的木姜子和黄丹木姜子为研究对象,采用g(r)函数对木姜子属两个种进行空间格局及空间关联分析,探讨其成因,探讨两物种是否存在相似性,同属物种共存机制,并为研究亚热带地区物种共存机制提供参考。
1 研究区概况
研究区(109°59′30″—110°17′58″ E、29°55′59″—30°10′47″ N)位于湖北省鹤峰县,面积为20 838 hm2,属于云贵高原的东北延伸区。该地区最低海拔为1 100 m,最高海拔为2 095.6 m,大陆性季风气候,冬季无严寒,夏季无极端炎热,雨热同季,日照充足,山地性气候明显,年均温15.5 ℃,年均降水量1 700~1 900 mm,年均相对湿度82%,无霜期270~279 d。土壤为黄棕壤、棕壤和黄壤。
木林子15 hm2大样地位于保护区核心区内,海拔为1 588~1 780 m,地势相对缓慢,属于亚热带山地常绿落叶阔叶混交林,主要树种有翅柃(Eurya alata)、多脉青冈(Cyclobalanopsis multinervis)、川陕鹅耳枥(C. fargesiana)、小叶青冈(C. myrsinifolia)、石灰花楸(Sorbus folgneri)、四照花(Dendrobenthamia japonica var. chinensis)、锥栗(Castanea henryi)、白栎(Quercus fabri)、山矾(Symplocos sumuntia)和光叶山矾(S. lancifolia)等(姚兰, 2016)。
2 研究方法
2.1 样地建设及野外调查
2013年,按照CTFS大样地建设与检测技术规范,建立15 hm2固定监测样地,分成375块20 m × 20 m小樣地,用防腐处理的不锈钢管将4角作为永久标识,每个小样地再分为4个10 m × 10 m样方,每个样方再分为4个5 m × 5 m的小样方,对DBH≥1.0 cm的木本植物进行挂牌,并用红色油漆标记(树高1.3 m),对DBH≥1 cm的所有木本植物进行每木检尺,识别物种(科、属、种),调查所有木本植物在样地内的坐标值(以大样地单元西南角为原点,测定x轴和y轴的坐标值)。
2.2 种群径级划分
运用径级代替龄级的方法,结合湖北木林子样地内木姜子和黄丹木姜子的径级特征,将胸径DBH划分为4个径级:Ⅰ级(1 cm≤DBH<2.5 cm),Ⅱ级(2.5 cm≤DBH<5 cm),Ⅲ级(5 cm≤DBH<10 cm),Ⅳ级(DBH≥10 cm),分别为幼树、小树、中树和成年树。
2.3 点格局分析方法
本研究采用点格局分析中成对相关函数g(r)进行分析。g(r)函数是基于对所有定位的成对个体之间距离的格局分析,消除了Ripieys K(r)函数在大尺度上产生的累积效应(Ripley, 1976; Thorsten et al., 2007)。分析时,选取距离尺度为较短边长的1/2(150 m),圆环宽度为1 m,通过199次Monte Carlo随机模拟,利用模拟的最大值和最小值绘制上下包迹线,计算99%的置信区间(Wiegand et al., 2006, 2009)。
2.4 零模型选择
采用点格局分析,需要检测生境异质性在物种分布中是否存在影响。一般来说,如果在大于10 m尺度上呈聚集分布,那就可以解释为受到生境异质性的影响;采用异质泊松模型和先决条件模型目的在于消除大尺度上生境异质性对空间分布格局的影响,在排除生境异质性后,如果小尺度的格局还是聚集,则说明扩散限制存在作用。所以在进行点格局分析时,不同的分析需要选择不同的零假设模型(null model),选取正确合适的零模型十分重要(梁爽等, 2014)。在本研究中单变量g(r)函数采用完全空间随机模型(complete spatial randomness, CSR)和异质泊松模型(heterogeneous Poisson, HP),双变量g(r)函数选择先决条件模型(antecedent condition, AC)。
文中数据的处理与分析使用R 3.4.1软件中的“spatstat”包中完成点格局分析,使用Sigmaplot 13软件绘制空间分布点,Origin 2018软件绘制点格局分析图。
3 结果与分析
3.1 两个物种空间分布格局
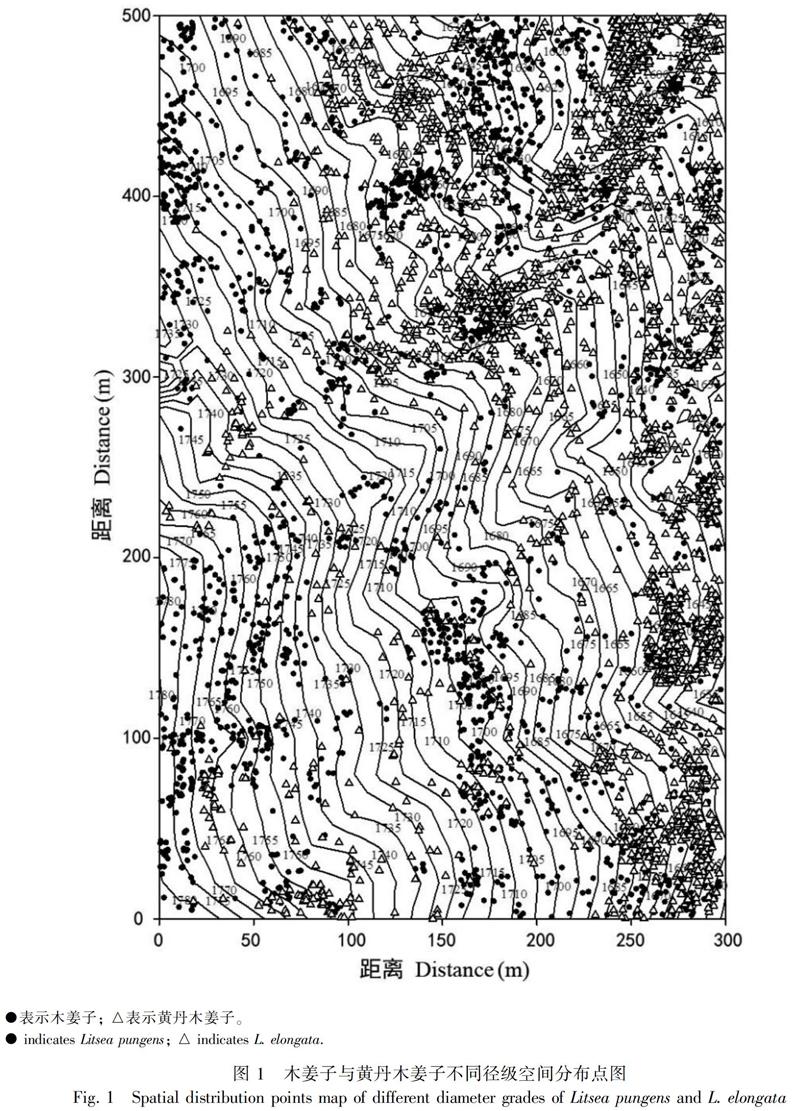
整个样地中(表1),木姜子1 701株,黄丹木姜子2 306株。由图1可知,两个物种的空间分布总体差异较大。木姜子主要分布于低海拔、山脊地区;黄丹木姜子主要分布于高海拔和山谷两侧区域,小部分零星分布于样地内。两个物种的整体空间分布格局的研究(图2)表明,基于CSR零模型:木姜子和黄丹木姜子的聚集尺度分别为0~114 m和0~150 m。基于HP零模型:木姜子的聚集尺度为0~23 m,其他尺度上为微弱的随机或均匀分布;黄丹木姜子的聚集尺度为0~36 m、140~150 m,随机分布尺度为48~139 m。
根据两个物种不同径级的空间分布格局研究(图3,图4)表明,基于CSR零模型:木姜子幼树、小树和中树的聚集尺度分别为0~103 m、0~56 m和0~34 m,成年树在0~16 m尺度表现为聚集分布;黄丹木姜子幼树、小树、中树和成年树分别在0~150 m、0~59 m、0~56 m和0~14 m尺度上均表现为聚集分布。基于HP零模型:木姜子幼树、小树和中树的聚集尺度均为0~22 m,而成年树仅在0~3 m尺度上表现为聚集分布;黄丹木姜子各径级的聚集尺度分别为0~35 m、0~35 m、0~12 m和0~6 m。由此可见,随着径级的增长,木姜子与黄丹木姜子均呈现聚集尺度逐渐减小的现象,特别是成年树仅在小尺度上表现为聚集分布,而随机分布格局现象比较明显。
3.2 两个物种不同径级种内关联性
木姜子不同径级种内关联见图5。基于AC零模型,成年树对幼树在所有尺度上均表现为无关联,而其他不同径级之间在较小尺度(r≤20 m)上表现为正相关,其中小树对幼树、中树对幼树、中树对小树、成年树对小树、成年树对中树正关联尺度分别为0~20 m、0~18 m、0~20 m、0~14 m和0~17 m。
黄丹木姜子不同径级种内关联见图6。基于AC零模型,黄丹木姜子成年树对中树在所有尺度上表现为无相关,小树对幼树、中树对幼树、成年树对幼树、中树对小树、成年树对小树分别在0~36 m、0~25 m、8~15 m、0~23 m和0~6 m尺度上呈现正关联。
3.3 两个物种种间关联性
用先决条件模型排除30 m以上生境异质性,结果显示:整体上,木姜子对黄丹木姜子在0~30 m呈现正关联,黄丹木姜子对木姜子在7~29 m呈现正关联(图7)。不同径级之间,木姜子不同径级对黄丹木姜子不同径级空间关联结果显示(图8):木姜子幼树对黄丹木姜子幼树在4~34 m尺度表现为正关联;木姜子小树对黄丹木姜子幼树在13~29 m尺度表现为正关联,42~67 m尺度上为负关联;木姜子中树对黄丹木姜子幼树在40~50 m尺度上为负关联;木姜子小树对黄丹木姜子小树在0~25 m尺度上为正关联,44~60 m尺度上为负关联;木姜子中树对黄丹木姜子中树在0~5 m尺度上为正关联,在47~54 m和119~128 m尺度上表现为负关联;其余径级之间均呈现无关联。黄丹木姜子不同径级对木姜子不同径级空间结果显示(图9):黄丹木姜子幼树对木姜子幼树在13~32 m尺度呈现正关联,42~76 m尺度呈现负关联;黄丹木姜子小树对木姜子幼树在0~13 m尺度呈现正关联;黄丹木姜子小树对木姜子小树在0~15 m呈现正关联,46~59 m尺度呈现负关联;其余径级之间均呈现无关联。
由此可见,木姜子对黄丹木姜子空间关系和黄丹木姜子对木姜子空间关系大致相同,仅有部分细微差别,其中两物种成年树与另一物种所有径级均没有显著的生态学关系。
4 讨论
4.1 两个物种空间分布格局形成的原因
植物自身生物学特性和环境因素共同作用于种群的空间分布格局,同一树种在不同的发育阶段将呈現不同的空间分布模式(Frost & Rydin, 2000)。本研究中基于CSR和HP零模型两个物种在研究尺度上主要表现为聚集分布,随着尺度增加表现为随机分布或均匀分布,结果与大部分研究一致(祝燕等, 2011; Guo et al., 2013)。在很大程度上,生境的异质性被认为是植被空间格局的主要原因。通常,如果它的聚集尺度大于10 m,则可以解释为受到栖息地异质性的影响(Takyu et al., 2002; Getzin et al., 2008)。基于CSR零模型,在整体上木姜子和黄丹木姜子分别在0~114 m和0~150 m尺度范围内呈聚集分布,并使用HP零模型排除了30 m以上的生境异质性,聚集尺度减小成为随机分布,可以推测,两个种群空间分布格局受到生境异质性影响。从幼树、小树、中树、成年树时间序列来看,分布格局从聚集分布向随机分布发展趋势,符合一般规律(Vargas et al., 2010; 张玉双等, 2011)。其中木姜子和黄丹木姜子不同径级之间均体现为幼树聚集尺度>小树聚集尺度>中树聚集尺度>成年树聚集尺度,可见生境异质性对两个物种的幼树影响最强,可能是在此阶段,种群的个体较小,生存力较低,需要通过聚集分布形成相互庇护,能够更好地维持种群自身的稳定性(张金屯, 1998; Wang et al., 2014)。生境异质性对两个物种的成年树影响较小,两个物种负密度制约超过了生境异质性对聚集分布的影响,随着径级的增大,对资源、对空间的需求也增加,物种内部的竞争日益激烈,物种内部的稀疏性越来越强,这将导致一些个体死亡从而形成均匀或随机分布的状态(舒兰等, 2019)。
在小尺度上,由于扩散限制可能导致聚集分布。当使用HP零模型分析,在较小尺度上两个物种均呈现聚集分布,这说明扩散限制对两物种的空间分布格局存在一定的作用。扩散限制使种子成熟后会在重力和风等因素的共同作用下落在母树周围,随着与母树距离越远,种子数量逐渐减少,从而在小尺度上形成聚集分布,木姜子和黄丹木姜子的果实均为球果,主要依靠重力传播为受距离限制。从两个物种来看,木姜子的聚集尺度范围和聚集程度均小于黄丹木姜子,可能是木姜子属于小乔木,一般高3~10 m,而黄丹木姜子属于中乔木,一般高达12 m,黄丹木姜子比木姜子种子的扩散范围更大,可能是黄丹木姜子在小尺度上的聚集尺度大于木姜子的原因。
種内和种间竞争也是影响种群聚集分布的一部分原因,种内关联是用来表示同一物种不同发育阶段在空间分布上的关联性(王磊等, 2010)。本研究发现两个物种之间存在较小种内竞争和种间竞争,其中种内竞争中仅发现黄丹木姜子的小树与幼树在一定的尺度上呈现负相关,说明这两个径级都属于演替前期,对资源和环境的需求趋于相同,种群不稳定,处于相互排除的阶段,幼树的生长受到了小树的抑制。种间竞争中,两个物种不同径级之间大致都趋于无关联的现象,仅在某些小尺度上呈现微弱的正关联和负关联的现象,说明两个物种种间竞争较弱,这可能是因为两物种不同的生境偏好导致的,研究发现木姜子个体更多偏向分布于高海拔和山脊地区,而黄丹木姜子更喜低海拔和山谷两侧。
4.2 两个同属物种相似性及共存机制
同一个生境中,同属物种的空间分布格局应该存在相似性。本研究结果显示,木姜子和黄丹木姜子总体上和不同径级之间的空间分布格局、两个物种的种内关联以及两个物种相互的种间关系均存在一定的相似性,两个物种总体上和不同径级之间均表现为聚集分布;在种内关联中除了木姜子的成年树对幼树和黄丹木姜子的成年树对中树表现出不同的空间关联,其余的均表现出相似的种内空间关系;在种间关联中木姜子对黄丹木姜子和黄丹木姜子对木姜子的种间关系均表现为幼树对幼树、小树对幼树、小树对小树存在较为强烈的生态学关系,其余径级之间生态学关系不显著。本研究显示,木姜子和黄丹木姜子在空间分布上存在一定的差异,可能是因为两物种不同的生长环境导致。
在一个区域内,多个物种肯定存在相互关系才会产生共存(宗国等, 2018)。本研究表明,总体上木姜子对黄丹木姜子和黄丹木姜子对木姜子的空间关系均表现为仅在40~68 m尺度内为负关联,其余尺度为正关联和无关联,而不同径级的木姜子和黄丹木姜子之间在大部分尺度上表现为正关联和无关联,仅在极小尺度上为负关联。可能是因为两个物种不同的生境偏好导致,从而减少两个物种对资源的竞争。同时,韦博良(2015)研究宝天曼栎属的空间分布格局时,发现栎属物种具有较小种间竞争关系,这样更有利于种群较小的物种能够共存。本研究显示,木姜子属两个物种之间存在较小的竞争关系,而是相互促进生长,这有利于两个物种共存,并同时抵御其他物种产生竞争关系。
5 结论
综上所述,木姜子和黄丹木姜子两个同属物种的空间分布存在一定的相似性,生境异质性、扩散限制和种内种间竞争都影响两个物种空间格局的形成,其中生境异质性对两个物种的幼树影响最大,扩散限制导致黄丹木姜子在排除生境异质性后扩散范围大于木姜子,两物种在种内关系中均体现为在小尺度上呈现正关联,同时两个物种之间存在较小的竞争关系;不同的生境偏好是两个物种在样地中实现共存的重要基础,同时也可能是因为同属物种容易形成相互庇护而抵御其他物种,则需要进一步的研究。
参考文献:
ACKERLY DD, DONOGHUE MJ, 1998. Leaf size, saplingallometry, and Corners rules: phylogeny and correlated evolution in maples(Acer) [J]. Am Nat, 152(6): 767-791.
CHEN J, AI XR, YAO L, et al., 2018. Point pattern analysis of two species of Cyclobalan opsis in large plot in Mulinzi Nature Reserve [J]. Sci Silv Sin, 54(10): 1-10. [陈俊, 艾训儒, 姚兰, 等, 2018. 木林子大样地两个青冈属优势种的点格局对比 [J]. 林业科学,
54(10): 1-10.]
CHEN YZ, 2004. Studies on the botanical origins and qualities of medicinal Litsea Lam [D]. Chengdu: Chengdu University of TCM. [陈幼竹, 2004. 木姜子属主要药用植物的品种品质研究 [D]. 成都: 成都中医药大学.]
CONDIT R, 2000. Spatial patterns in the distribution of tropical tree species [J]. Science, 288(5470): 1414-1418.
DIGGLE PJ, 2003. Statistical analysis of spatial point patterns [M]. 2nd ed. London: Hodder Education Publishers.
DONG S, 2016. Study on leaf litterfall production and nutrient characteristcs of six dominant species in an evergreen broad-leaved forest in Tiantong [D]. Shanghai: East China Normal University. [董舒, 2016. 天童常绿阔叶林6个优势种凋落量及养分特征研究 [D]. 上海: 华东师范大学.]
FAN FR, PAN BZ, MA XQ, et al., 2008. A study on structure and spatial distribution pattern of Artocarpus hypargyreus population [J]. For Res,(2): 176-181. [范繁荣, 潘标志, 马祥庆, 等, 2008. 白桂木的种群结构和空间分布格局研究 [J]. 林业科学研究, (2): 176-181.]
FROSTI, RYDIN H, 2000. Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests [J]. coscience, 7(1): 38-44.
GETZIN S, WIEGAND T, WIEGAND K, et al., 2008. Heterogeneity influences spatial patterns and demographics in forest stands [J]. J Ecol, 96(4): 807-820.
GUO YL, LU JM, FRANKLIN SB, et al., 2013. Spatial distribution of tree species in a species-rich subtropical mountain forest in central China [J]. Can J For Res, 43(9): 826-835.
HE ZL, XU H, QIN XS, et al., 2017. Spatial distribution patterns and association of two Apocynaceae plants in the tropical mountain rainforests of Jianfengling, Hainan Island, China [J]. Biodivers Sci, 25(10): 1065-1074. [何增麗, 许涵, 秦新生, 等, 2017. 海南尖峰岭热带山地雨林2种夹竹桃科植物的空间分布格局与关联性 [J]. 生物多样性, 25(10): 1065-1074.]
HUANG YT, 2016. The functional community structure and its dynamics of the subtropical evergreen-deciduous broadleaved mixed forest in Southwest Hubei of Central China [D]. Beijing: Chinese Academy of Forestry. [黄永涛, 2016. 亚热带常绿落叶阔叶混交林的功能性群落结构及其动态研究 [D]. 北京: 中国林业科学研究院.]
HUANG XB, LI SF, SU JR, et al., 2018. Distribution of Pinus yunnanensis natural population in Yunlong Tianchi National Nature Reserve [J]. For Res, 31(4): 47-52. [黄小波, 李帅锋, 苏建荣, 等, 2018. 云龙天池云南松自然种群分布格局分析 [J]. 林业科学研究, 31(4): 47-52.]
LIANG S, XU H, LIN JY, et al., 2014. Spatial distribution pattern of the dominant species Gironniera subaequalis in tropical montane rainforest of Jianfengling, Hainan Island, China [J]. Chin J Plant Ecol, 38(12): 1273-1282. [梁爽, 许涵, 林家怡, 等 , 2014. 尖峰岭热带山地雨林优势树种白颜树空间分布格局 [J]. 植物生态学报, 38(12): 1273-1282.]
LIU WX, LIAN YM, LI W, 2018. Effects of microtopography on the pattern and association of dominant population [J]. Chin J Appl Ecol, 29(5): 1569-1575. [刘旻霞, 连依明, 李文, 2018. 微地形对优势种群点格局和关联性的影响 [J]. 应用生态学报, 29(5): 1569-1575.]
NATHAN R, 2006. Long-distance dispersal of plants [J].
Science, 5788(3): 786-788.
QU ZX, 1983. Plant ecology [M]. Beijing: Higher education press. [曲仲湘, 1983. 植物生态学 [M]. 北京: 高等教育出版社.]
RIPLEY DB, 1976. The second-order analysis of stationary point processes [J]. J Appl Prob, 13(2): 255-266.
SHU L, LIU ZG, DONG LB, 2019. Spatial pattern and regeneration characteristics of main woody species in natural secondary forest in Maoershan, Northeast China [J]. Chin J Appl Ecol, 30(6): 1945-1955. [舒兰, 刘兆刚, 董灵波, 2019. 帽儿山天然次生林内主要木本植物空间格局及更新概况 [J]. 应用生态学报, 30(6): 1945-1955.]
SWENSON NG, ENQUIST BJ, PITHER J, et al., 2006. The problem and promise of scale dependency in community phylogenetics [J]. Ecology, 87(10): 2418-2424.
TAKYU M, AIBA SI, KITAYAMA K, 2002. Effects of topography on tropical lowermontane forests under different geological conditions on Mount Kinabalu, Borneo [J]. Plant Ecol, 159(1): 35-49.
THORSTEN W, SAVITRI G, NIMAL G, et al., 2007. Analyzing the spatial structure of a Sri Lankan tree species with multiple scales of clustering [J]. Ecology, 88(12): 3088-3102.
VARGAS R, CUEVAS JG, QUESNE CL, et al., 2010. Spatial distribution and regeneration strategies of the main forest species on Robinson Crusoe Island Distribución espacial y estrategias de regeneración de las principales especies forestales de la Isla Robinson Crusoe [J]. Rev Chil De Hist Nat, 83(3): 349-363.
WANG L, SUN QW, HAO ZY, et al., 2010. Point pattern analysis of different age-class Taxus chinensis var. mairei individuals in mountainous area of southern Anhui Province [J]. Chin J Appl Ecol, 21(2): 272-278. [王磊, 孙启武, 郝朝运, 等, 2010. 皖南山区南方红豆杉种群不同龄级立木的点格局分析 [J]. 应用生态学报, 21(2): 272-278.]
WANG XT, LIANG CZ, WANG W, 2014. Balance between facilitation and competition determines spatial patterns in a plant population [J]. Chin Sci Bull, 59(13): 1405-1415.
WEBB CO, ACKERLY DD, MCPEEK MA, et al., 2002. Phylogenies and community ecology [J]. Ann Rev Ecol Syst, 475-505.
WEI BL, 2015. Point pattern analysis and similarity study of the Quercus species at Baotianman, Henan, China [D]. Zhengzhou: Henan Agricultural University. [韋博良, 2015. 宝天曼栎属物种的空间格局及其相似性分析 [D]. 郑州: 河南农业大学.]
WIEGAND T, KISSLING W, CIPRIOTTI PM, 2006. Extending point pattern analysis for objects of finite size and irregularshape [J]. J Ecol, 94(4): 825-837.
WIEGAND T, MARTNEZ I, HUTH A, 2009. Recruitment in tropical tree species: Revealing complex spatial patterns [J]. Am Nat, 174(4): 106-140.
WIEGLEB G, 1989. Explanation and prediction in vegetation science [J]. Vegetatio, 83(1/2): 17-34.
YAN HB, 2017. Spatial distribution patterns of tree species and their correlation with habitat in secondary cold-warm Picea forest in Guandi Mountain [D]. Jinzhong: Gradute College of Shanxi Agricultural University. [闫海冰, 2018. 关帝山寒温性云杉次生林树种空间分布格局及其与生境相关性 [D]. 晋中: 山西农业大学.]
YAO L, 2016. The foresr community structure and diversity of 15 hm2 dynamics plot in Hubei Mulinzi Nature Reserve [D]. Beijing: Beijing Forestry University. [姚兰, 2016. 湖北木林子保护区15 hm-2大样地森林群落结构及多样性 [D]. 北京: 北京林业大学.]
YAO LJ, YAO L, YI YM, et al., 2018. Spatial patterns of dominant species Carpinus fargesiana and Cyclobalanopsis multinervis in a subtropical evergreen and deciduous broad-leaved mixed forest [J]. Sci Silv Sin, 54(12): 1-11. [姚良锦, 姚兰, 易咏梅, 等, 2018. 亚热带常绿落叶阔叶混交林优势种川陕鹅耳枥和多脉青冈的空间格局 [J]. 林业科学, 54(12): 1-11.]
ZHANG JT, 1995.Quantitative plant ecology [M]. Beijing: Science and Technology of China Press. [张金屯, 1995. 植被数量生态学方法 [M]. 北京: 中国科学技术出版社.]
ZHANG JT, 1998. Analysis of spatial point pattern for plant species [J]. Chin J Plant Ecol, 22(4): 344-349. [张金屯, 1998. 植物种群空间分布的点格局分析 [J]. 植物生态学报, 22(4): 344-349.]
ZHANG JT, MENG DP, 2004. Spatial pattern analysis of individuals in different age-classes of Larix principis-rupprechtii in Luya mountain reserve, Shanxi, China [J]. Acta Ecol Sin,(1): 35-40. [张金屯, 孟东平, 2004. 芦芽山华北落叶松林不同龄级立木的点格局分析 [J]. 生态学报, (1): 35-40.]
ZHANG YS, DUAN WB, DU S, et al., 2016. Analysis on spatial distribution pattern for main populations and gap makers in Korean pine broad-leaved forest in Xiaoxinganling Mountains of Northeast China [J]. For Res, 29(3): 395-401. [張玉双, 段文标, 杜珊, 等, 2016. 阔叶红松林主要种群及林隙形成木的空间格局分析 [J]. 林业科学研究, 29(3): 395-401.]
ZHAO ZH, HUI GY, HU YB, et al., 2011. Spatial distribution patterns and associations of dominate population in two types of broadleaved Korean pine mixed forest [J]. For Res, 24(5): 554-562. [赵中华, 惠刚盈, 胡艳波, 等, 2011. 2种类型阔叶红松林优势种群空间分布格局及其关联性 [J]. 林业科学研究, 24(5): 554-562.]
ZHU Y, BAI F, LIU HF, et al., 2001. Population distribution patterns and interspecific spatial associations in warm temperate secondary forests, Beijing [M]. Biodivers Sci, 19(2): 252-259. [祝燕, 白帆, 刘海丰, 等, 2011. 北京暖温带次生林种群分布格局与种间空间关联性 [J]. 生物多样性, 19(2): 252-259.]
ZONG G, BAI XJ, ZHANG SY, et al., 2018. Spatial pattern and interspecific spatial association of tree seedlings in a secondary forest in montane region of eastern Liaoning Province, China [J]. Chin J Appl Ecol, 29(1): 18-24. [宗国, 白雪娇, 张淑媛, 等, 2018. 辽东山区次生林乔木幼苗分布格局与种间空间关联性 [J]. 应用生态学报, 29(1): 18-24.]
(责任编辑 周翠鸣)
