柳江下游江段伍氏华吸鳅早期发育及其资源补充量评估
2021-09-13黄海博,高明慧,吴志强,黄亮亮,谭细畅,刘昊,陈建林,黄欣
黄海博,高明慧,吴志强,黄亮亮,谭细畅,刘昊,陈建林,黄欣


摘要:【目的】觀察伍氏华吸鳅早期发育并评估柳江下游江段其资源补充量,为柳江下游江段伍氏华吸鳅的渔业资源开发及可持续利用提供基础数据。【方法】2017年5月16日—8月31日于柳江下游石龙镇进行伍氏华吸鳅的鱼卵及鱼苗采集,对采集到的伍氏华吸鳅鱼卵进行人工培育,重点观察并详细描述其仔鱼时期的形态变化特征;并通过设置断面方法估算伍氏华吸鳅在柳江下游江段的资源补充量。【结果】伍氏华吸鳅卵呈球形,单层卵膜,淡黄色,卵径为4.50±0.10 mm;初孵仔鱼全长5.00±0.21 mm,肌节35对(3+20+12),眼色素和胸鳍原基出现,身体呈淡黄色,卵黄囊前部已有半球形点状色素分布,心跳约165次/min;出膜25 d,全长18.50±0.35 mm,身体呈淡红色,臀鳍、胸鳍和腹鳍均基本形成,完成仔鱼期发育,进入稚鱼阶段。调查期间柳江下游江段采样断面鱼卵和鱼苗的分布系数为0.416,柳江下游江段伍氏华吸鳅鱼卵的群体补充量为2.48×108粒。Spearman相关分析结果显示:伍氏华吸鳅鱼卵密度与流量和水位呈极显著负相关(r=-0.688,P<0.01;r=-0.663,P<0.01),与透明度和溶解氧呈极显著正相关(r=0.667,P<0.01; r=0.297,P<0.01)。【结论】伍氏华吸鳅体长相对其他平鳍鳅科鱼类偏小,其卵为无黏性漂流鱼卵,产卵量与流量和水位呈极显著负相关,与透明度和溶解氧呈极显著正相关,因此柳江下游江段伍氏华吸鳅群体资源补充量可能在大藤峡水库蓄水完成后明显下降。
关键词: 伍氏华吸鳅;早期发育;资源补充量;环境因子;柳江下游
中图分类号: S932.4 文献标志码: A 文章编号:2095-1191(2021)09-2590-09
Early development of Sinogastromyzon wui and its resource supplement in the lower reaches of Liujiang River
HUANG Hai-bo1, GAO Ming-hui2, WU Zhi-qiang1*, HUANG Liang-liang1,3,
TAN Xi-chang4, LIU Hao1, CHEN Jian-lin1, HUANG Xin1
(1College of Environmental Science and Engineering, Guilin University of Technology/Guangxi Collaborative Innovation Center for Water Pollution Control and Water Safety in Karst Areas, Guilin, Guangxi 541004, China; 2College of Life Science and Technology, Guangxi University, Nanning 530004, China; 3Guangxi Key Laboratory of Environmental Pollution Control Theory and Technology for Science and Education Combined with Science and Technology
Innovation Base, Guilin, Guangxi 541004, China; 4Bureau of Hydrology and Water Resources, Pearl River Conservancy Commission of Ministry of Water Resources, Guangzhou 510380, China)
Abstract:【Objective】In order to provide basic data for the resource assessment and sustainable utilization of Sinogastromyzon wui in the lower reaches of Liujiang River, the study observed early development of S. wui and evaluated its resource in the lower reaches of Liujiang River. 【Method】Fish larvaland fish eggs were collected in Shilong Town, lower reaches of the Liujiang River from May 16 to August 31, 2017. In this study, the collected eggs were artificially cultiva-ted, and the morphological changes of the larvae were observed and described in detail. The resource supplement of S. wui in the lower reaches of the Liujiang River was estimated by setting a cross section. 【Result】The shape of the fertilized eggs of the fish were ovate with monolayer and yellow colour, which was 4.50±0.10 mm in diameter. The total length of the newly hatched larvae was 5.00±0.21 mm, with 35 pairs of muscle segments 3+20+12. Meanwhile eye pigment and pectoral fin primordia appeared, the heart rate was about 165 times/min and the body was pale yellow with hemispherical spotty pigment distribution in the front part of the yolk sac.Anal fin, pectoral fin and abdominal fin were basically formed and the development of the larvae stage was completed in 25 d after hatching. The total length was 18.50±0.35 mm, and the body was light red. During the investigation period, the distribution coefficient in the sampling section of the lower reaches of the Liujiang River was 0.416 and the resources replenishment of S. wui was estimated to be 2.48×108 eggs du-ring the sampling period in Liujiang River. The Spearman correlation analysis showed that, the density of fish eggs was negatively related to the discharge, and water level significantly (r=-0.688,P<0.01;r=-0.663,P<0.01), while it had significant positive correlation with transparency and the dissolved oxygen (r=0.667,P<0.01;r=0.297,P<0.01). 【Conclusion】Compared with Balitoridae, the length of S. wui is small and its eggs are non-viscous drifting eggs. The spawning of S. wui is significantly negatively correlated with discharge and water level, and significantly positively correlated with transparency and dissolved oxygen. Therefore, the population resource supplement of S. wui in the lower reaches of Liu-jiang River may decrease after the completion of the Datengxia Reservoir.
Key words: Sinogastromyzon wui; development of early phase; resource replenishment; environmental factor; lower reaches of Liujiang River
Foundation item: National Natural Science Foundation of China(32060830); Guangxi Natural Science Foundation(2018GXNSFAA281022);Chunhui Program of Ministry of Education(Jiaowaisirenwen〔2020〕703); Guangxi Bagui Scho-lars Special Project(2016A10)
0 引言
【研究意義】华吸鳅属(Sinogastromyzon)目前共记载有22个种(杨骏和郭延蜀,2013),主要栖息于我国南部水系及越南北部水系(Liu et al.,2010)。其中,伍氏华吸鳅(S. wui)主要分布在左江、红水河、融江、柳江及珠江等水系(广西水产研究所和中国科学院动物研究所,2006),是柳江鱼类的重要组成部分,但国内外针对于伍氏华吸鳅的研究资料甚少。大藤峡水利枢纽工程坝址位于珠江水系西江流域黔江干流大藤峡出口(王丽等,2016),柳江在其上游,水利水电工程蓄水后大坝上游河段“湖泊”化,河流形态和径流特征发生明显变化,对鱼类的产卵、生活及其种群结构等方面均存在严重影响(梁晓华等,2013)。因此,迫切需要开展对伍氏华吸鳅的早期资源调查,为柳江鱼类资源开发及渔业管理提供理论依据。【前人研究进展】鱼类早期资源调查以早期生活史阶段的鱼类为对象进行调查研究,是研究鱼类生态学及渔业生物学的重要手段,对鱼类资源开发和渔业管理具有重要意义(曹文宣等,2007;余为等,2013;王永杰等,2018)。近年来,有关西江流域鱼类早期资源调查的研究主要集中在中下游江段。谭细畅等(2007,2009a,2009b)对肇庆江段鱼类早期资源的时空分布特征进行研究,并系统性描述了鲤(Cyprinus carpio)和赤眼鳟(Squaliobarbus curriculus)的早期发育特征。帅方敏等(2016)对位于桂平江段东塔产卵场鳙(Hypophthalmichthys nobilis)繁殖的生态水文需求进行分析,并采用交互小波光谱分析法分析径流量与鳙仔鱼多度的关系。高明慧(2018)对来宾江段鱼类早期资源及其与环境因子的关系进行调查,并研究鱼类与环境因子间的相关性,为来宾江段鱼类早期发育特征的种间比较和资源现状评估提供了基础资料。匡天旭(2018)基于COI基因对珠江鮊亚科早期资源从种类鉴定到群落分析进行系统探索。李策等(2019)通过形态学识别和DNA条形码鉴定方法对西江肇庆江段的鱼类进行种类鉴定,并评估该江段优势种类的资源现状。【本研究切入点】鱼类早期发育是其形态、生态及生理行为等方面动态变化的过程,也是鱼类生活史中死亡率较高的时期,直接影响鱼类群体的补充量(胡飞飞和陈新军,2015),但目前国内外有关伍氏华吸鳅早期发育形态及其资源量的调查研究鲜见报道。【拟解决的关键问题】对从柳江下游江段采集到的伍氏华吸鳅鱼卵进行培养和观察,重点描述处于仔鱼时期的形态发育情况,并与同科种类进行种间比较,以补充其早期形态学方面的资料;同时通过设置断面方法估算伍氏华吸鳅在柳江下游江段的资源补充量,以期为其渔业资源开发及可持续利用提供基础数据。
1 材料与方法
1. 1 调查方法
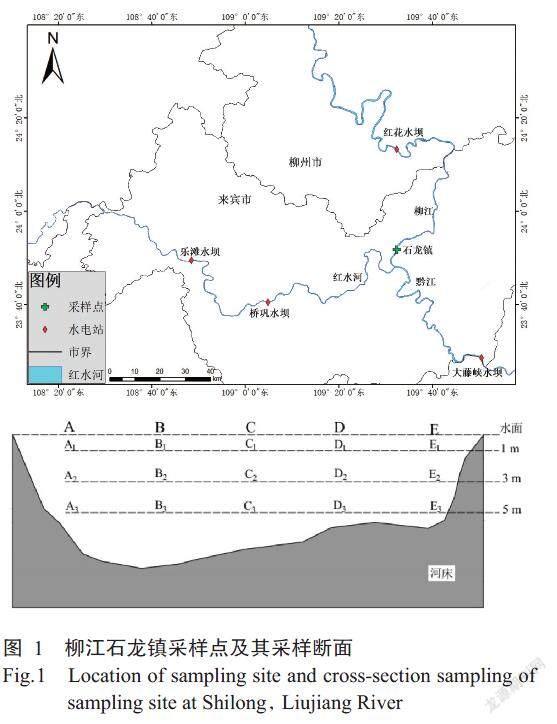
2017年5月16日—8月31日,于柳江下游石龙镇(东经109°31'30″,北纬23°52'21″)设置断面开展鱼类早期资源的常规采集工作(图1),期间每日上午7:00—8:00于柳江左岸近岸表层使用弶网(网口1.0 m×1.5 m,网目0.5 mm,网长6.0 m)和集苗箱(0.8 m×0.4 m×0.4 m)进行常规采集。断面采样开展在2017年7月29日上午6:00—11:00进行,表层采集使用弶网完成,中层和底层使用圆锥网(网口面积0.196 m2,网长1.5 m,网目0.5 mm),各采样点的采集时间为15~20 min。
1. 2 种类鉴定与培育
将采集到的鱼卵带回实验室,于体视解剖镜下依据卵膜颜色、性质、直径和肌节等特征进行种类鉴定(曹文宣等,2007)。对不能确定种类的鱼卵进行分类及编号,放入50 mL的培养杯中单独培养,待发育至稚鱼期再进一步鉴别(易伯鲁等,1988;乔晔,2005)。水温维持在22~25 ℃,每日换采集的河水1次,保证溶解氧充足及水中成分稳定。胚胎孵出2 d后开始投喂熟蛋黄直至稚鱼期,此后投喂河水中的丰年虾;在投喂熟蛋黄30 min后将多余的蛋黄吸出以维持水质良好。
1. 3 观察方法
对单独培养的鱼卵在体视显微镜下以尼康相机(COOLPIX 5400)进行观察及拍照,其中,卵期每30 min观察拍照1次,卵黄囊期仔鱼每8 h观察拍照1次,之后根据发育情况逐步延长观察拍照的时间间隔。记录及拍照结束后将仔鱼继续单独培养,对难以活体拍照的时期则以5%福尔马林固定后进行拍照,同时从单独培养的鱼卵中选取相同种类作为下一发育期的描述对象。以此方法对每日采集的伍氏华吸鳅鱼卵进行多样本的形态学描述和统计。
1. 4 数据分析
鱼卵及鱼苗总径流量统计参考Mu等(2014)的方法,计算公式为:
A=M+M'
M=[i=1n(CTiQiDi)]
M'=[i=1nCT′(i,i+1)×(Qi+Qi+1)/2×(Di+Di+1)/2]
Di=Ni/(SViTi)
C=[j=1mDi/m]/DA1
式中,A为调查期间通过设置断面的总鱼卵及鱼苗径流量(ind),M为常规采集过程中通过设置断面的鱼卵和鱼苗总量(ind),M'为2次采集间隔期间通过设置断面的鱼卵和鱼苗总量(ind),n为常规采集样本的数量(n=107),i是指特定的一次常规采集(“i”、“i+1”即第i次和下一次采集),Ti是第i次采集的持续时间(s),Qi是第i次采集过程中的径流量(m3/s),Di是第i次采集中鱼卵和鱼苗的漂流密度(ind/m3),C为采样断面鱼卵和鱼苗的分布系数,T'(i,i+1)第i次和i+1次采集的时间间隔(s),Qi+1是第i+1采集过程中的径流量(m3/s),Di+1是第i+1次采集中鱼卵和鱼苗的漂流密度(ind/m3),Ni是第i次断面采集收集到的鱼卵和鱼苗数量(ind),S是网口面积(m2),Vi是第i次断面采集时的网口流速(m/s),Dj是断面采集过程中采样点的鱼卵和鱼苗密度(ind/m3),j是指断面采集过程中的采样点,m是指断面采样点的数量,[ DA1]是指断面采样中采样点A1的鱼卵和鱼苗密度(ind/m3)。
采集过程中,使用LJD-10A打印式流速仪测量网口流速以计算滤水量,水银温度计测量气温和水温,萨氏盘测量水体透明度,并通过查询全国水雨情信息网(http://xxfb.hydroinfo.gov.cn)获取水位和流量数据。
2 结果与分析
2. 1 伍氏华吸鳅早期形态特征
2. 1. 1 伍氏华吸鳅鱼卵特征 伍氏华吸鳅鱼卵呈球形,单层卵膜,表面光滑无黏性,属漂流性卵;卵径为4.50±0.10 mm;卵为淡黄色,卵膜透光性好。所采集的鱼卵多数已发育至孵出期,胚体在卵膜内抖动剧烈,经3~5 h后孵出。
2. 1. 2 仔鱼阶段 初孵仔鱼(图2-A):全长5.00±0.21 mm,肌节35对(2+20+13);胸鳍原基明显;眼睛已有色素沉着,头部尚未与身体分离;卵黄囊呈锥形;尾鳍褶呈圆形;肛门原基形成;身体呈淡黄色、卵黄囊前部已有半球形点状色素分布;心脏位于卵黄囊前方,由心房、心室和静脉窦组成,内部血液流动明显,心跳约165次/min;仔鱼活动能力较强。
出膜1 d(图2-B):全长5.20±0.21 mm,肌节36对(3+19+14);头部与身体分离,口裂基本形成;卵黄囊前部色素增多;卵黄囊体积减小;眼睛黑色素沉着基本完成;仔鱼鳃丝长出,鳃盖显现;仔鱼尾端呈圆形,会不时向上冲游。
出膜2 d(图2-C):全长5.50±0.22 mm,肌节36对(3+19+14);颌须1对,口周围和口须上出现刺状感觉刚毛;卵黄囊体积进一步收缩呈长条形,前端形成空泡;心脏分化完成;部分鱼类开始平游。
出膜3 d(图2-D):全长5.80±0.23 mm,肌节36对(3+19+14);鳔雏形;口角须2对;体色加深,背部开始有黑色素沉着;鳃丝收缩;卵黄囊几乎消耗殆尽;口裂形成,口下位;尾鳍褶开始分化,其下叶出现放射状鳍条;仔鱼可平游,但时常静卧水底不动。
出膜4 d(图2-E):全长6.03±0.26 mm,肌节37对(3+19+15);鳔一室形成;卵黄囊消耗殆尽,肠道中有食物堆积;须4对,口角须与吻须各2对;鳃丝消失,内鳃形成,呈红色;尾椎开始上翘。
出膜6 d(图2-F):全长6.12±0.19 mm,肌节37对(3+19+15);尾椎完全上翘;胸鳍呈扇形,垂直伸展于身体两侧;头呈三角形,身体细长;背鳍褶前部稍隆起,开始有放射状鳍条出现;尾鳍继续分化,鳍条迅速增多,基部出现上、下2朵色素花。
出膜8 d(图2-G):全长7.30±0.26 mm,肌节37对(3+19+15);头背及眼后色素增多,体背色素由脑后一直延申至尾鳍基部,形成2条明显的纵向色素带;背鳍继续分化,臀鳍鳍条出现3~4根,胸鳍开始平铺于身体两侧。
出膜10 d(图2-H):全长8.70±0.25 mm,肌节37对;鳔二室;身体色素进一步增多,头背色素呈两头连接的三角形,2个鼻孔外侧各出现1条黑色纵纹;背鳍形成,腹鳍继续分化,并开始平铺。
出膜14 d(图2-I):全长11.0±0.31 mm,臀鳍形成,背鳍鳍条10根,臀鳍鳍条7根,臀鳍与臀鳍褶分离;头部及身体前部变得扁平,背缘呈弧形;上唇增厚,且出现乳突,口须延申。
出膜20 d(图2-J):全长13.50±0.33 mm,腹鳍形成,鳍条10~12根;胸鳍鳍条16根,呈椭圆形,较大平铺;尾截性,边缘开始内凹,基部色素增多,上下连接成线。
出膜25 d(图2-K):全长18.50±0.35 mm,身体呈淡红色,鳃部及胸鳍间呈鲜红色,尾鳍分叉,各鳍条发育完成,進入稚鱼期。
2. 2 伍氏华吸鳅产卵日期及其群体补充状况
柳江下游伍氏华吸鳅的主要产卵周期为5月下旬—7月底,期间出现2次明显的产卵高峰,分别为5月18日—6月10日和7月20日—8月1日。鱼卵密度为0~22粒/100 m3,平均为3.2粒/100 m3。断面鱼卵采集量如表1所示,结合断面系数进行估算,2017年5月16日—8月30日柳江下游江段采样断面鱼卵和鱼苗的分布系数为0.416,柳江下游江段伍氏华吸鳅鱼卵的群体补充量为2.48×108粒。
2. 3 伍氏华吸鳅鱼卵密度与环境因子的关系
采样期间,位于采样点上游24 km的象州水文站提供数据显示该江段水位变化为77.12~83.10 m,平均为78.41 m,流量变化范围为738~17800 m3/s。水位、流量和水温低值均出现在5月,高值则出现在7月,而溶解氧和透明度与之相反。现场实测数据(图3)显示,水温变化为22.6~30.0 ℃,溶解氧变化为3.04~7.32 mg/L,透明度变化为15~115 mm。Spearman相关分析结果(图4~图7)显示:伍氏华吸鳅鱼卵密度与流量和水位呈极显著负相关(r=-0.688,P<0.01;r=-0.663,P<0.01),与透明度和溶解氧呈极显著正相关(r=0.667,P<0.01;r=0.297,P<0.01)。从水文条件来看,透明度与流量和水位呈显著负相关(r=-0.699,P<0.05;r=-0.757,P<0.05),溶解氧与流量和水温也呈显著负相关(r=-0.436,P<0.05;r=-0.439,P<0.05)。
3 讨论
3. 1 伍氏华吸鳅早期发育形态特征
分布于广西的平鳍鳅科(Homalopteridae)鱼类相对较原始,因此有研究认为广西可能是平鳍鳅科鱼类的起源中心。广西山溪支流中广泛分布有平鳍鳅科鱼类16种,其中仅有3个种为广西特有种,其他13个种已扩散至珠江中下游江段(陈旻等,2002)。至今,有关平鳍鳅科鱼类早期形态发育的研究相对较少,仅熊玉宇等(2008)、吴金明等(2011)分别对金沙江犁头鳅(Lepturichthys fimbriata)和赤水河四川华吸鳅(S. szechuanensis)进行胚胎早期及仔稚鱼期的系统性描述。伍氏华吸鳅与四川华吸鳅和犁头鳅同属于平鳍鳅科,3种仔鱼在早期发育过程中表现出一定的趋同性:(1)初孵仔鱼形态相似,背鳍褶起点接近头部与第3对肌节对应,眼径小,小于全长的1/30,卵黄囊呈锥形,头部紧贴卵黄囊前部,未与身体分离。(2)体形与体表色素的发育过程相似,初孵仔鱼体形相似,体表透明无色素,伴随着卵黄消耗,仔鱼头部变得扁平且呈三角形,身体变细长,体表色素逐渐增多,头背形成三角形色素斑,体背纵向色素和肌节横向色素也逐渐显现。(3)器官形成顺序及时间相似,3种初孵仔鱼胸鳍均发育快速,胸鳍从垂直立于身体两侧至卵黄囊吸尽时期已发育成较大扇形;孵出2~3 d颌须出现,此时口型由亚下位发育为下位。3种仔鱼发育过程中也存在趋异性:(1)卵粒性质不同,漂流性卵吸水膨胀,随时间增长鱼卵直径增大,密度减少,平均卵径约在4.80 mm,相对比密度为1.0014~1.0031,略高于水(Dudley and Platania,1999);而黏沉性卵粒径较大,比重大,且有黏性,易附着(刘雪飞,2019)。伍氏华吸鳅和犁头鳅的鱼卵吸水后膨胀,卵径分别为4.50和6.50 mm,卵膜径约为1.85 mm,因此判断为无黏性漂流鱼卵;而四川华吸鳅鱼卵为黏沉性鱼卵。(2)初孵仔鱼的体长与总肌节数不同,伍氏华吸鳅和四川华吸鳅的体长相近,约4.50 mm,肌节数分别为35和37对;而犁头鳅的体长可达6.50 mm,肌节数为39对。3种仔鱼发育过程的形态特征及部分可数形状比较见表2。
总体来看,鱼类在发育过程中表现出的趋同性和趋异性均能反映其在不同生境下所采取的适应性生存策略。多数鱼类选择夏季产卵,是由于水流量增大有利于漂流性卵鱼的扩散,水温升高有利于鱼卵的快速发育,且饵料丰富能有效提高仔鱼的存活率(广西水产研究所和中国科学院动物研究所,2006)。对于平鳍鳅科鱼类而言,胸鳍和腹鳍的优先发育促使其更快地由漂流向底栖生活转变(熊玉宇等,2008),而口须长和眼径小是鱼类生活在水流湍急江河中靠化学气味觅食的典型适应特征(Wang and Liu,2005)。
3. 2 柳江下游江段伍氏华吸鳅补充群体资源量预测
平鳍鳅科鱼类好流水环境,喜生活在山涧石滩和江河急流中,营底栖生活,常吸附于岩石壁或底部,以刮食岩石表面苔藓为生。广西伍氏华吸鳅集中分布于在漓江、柳江及红水河等西江水系的支流中(陈旻等,2002)。这些江段地势高,水位变化明显,水流速度快,且水体中溶解氧含量较高,为伍氏华吸鳅提供了适宜的产卵环境及鱼卵发育条件,鱼卵产出后迅速吸水膨胀,形成较大的卵周隙,顺水漂流在流水中迅速分散,既能防止被泥沙淤埋,又降低被捕食的风险。河道疏浚、航运、挖沙及护岸等水利工程对仔鱼生存和繁殖均有一定影响(封文利等,2017),大库区水位升高可导致以沙洲、滩涂和卵石为产卵环境的鱼类产卵场所遭到破坏(曹文宣等,2007);水流变缓,水中溶解氧含量下降易导致漂流性卵下沉或因缺氧而无法正常发育(周春生等,1980)。本研究结果显示,伍氏华吸鳅的产卵受流量、水位及溶解氧等多种因素的影响,与流量和水位呈极显著负相关,与透明度和溶解氧呈极显著正相关,与刘孟凯等(2017)的研究结论一致。虽然水温和鱼类产卵的相关性不显著,但许多研究发现水温对鱼类的产卵时间有明显影响,陶雨薇等(2018)研究发现三峡水库运行导致的水温变化致使中华鲟繁殖时间约推迟29 d。综上所述,在产卵场所受到破坏及鱼卵无法完成正常发育的双重威胁下,柳江下游江段伍氏华吸鳅群体资源补充量可能在大藤峡水库蓄水完成后明显下降。
3. 3 伍氏华吸鳅的资源保护措施
水利工程在发电、防洪及蓄水等方面能对社会产生巨大的经济效应,但在不同程度上也改变了生态环境,影响其连通性而导致一系列的生态问题(Rand and Fukushima,2014)。长江水利工程的修建致使长江渔业资源严重下降,甚至导致部分鱼类物种濒临灭绝(Zhang et al.,2020)。针对伍氏华吸鳅的资源保护,本研究提出以下3个保护措施:(1)通过调查、评估和分析渔业资源情况,建立统计模型,设定捕捞限额,积极管理渔业,以期有效改善渔业情况及保护鱼类资源(Hilborn and Ovando,2014;Roa-Ureta et al.,2020)。(2)实时监控,科学管理,尤其是加强水域生态环境健康监测,建立覆盖全流域和所有环境要素的监测网络体系(刘飞等,2020)。(3)综合考虑柳江江段其他鱼类物种与环境因子的关系,科学设定蓄水和排放量,同时结合气象水文资料,人为制造合理的洪峰以满足气象水文规律。
4 结论
伍氏华吸鳅体长相对其他平鳍鳅科鱼类偏小,其卵为无黏性漂流鱼卵,产卵量与流量和水位呈极显著负相关,与透明度和溶解氧呈极显著正相关,因此柳江下游江段伍氏華吸鳅群体资源补充量可能在大藤峡水库蓄水完成后明显下降。
参考文献:
曹文宣,常剑波,乔晔,段中华. 2007. 长江鱼类早期资源[M]. 北京:中国水利水电出版社. [Cao W X,Chang J B,Qiao Y,Duan Z H. 2007. Fish resources of early life history stages in Yangtze River[M]. Beijing:China Water & Power Press.]
陈旻,黄宁,李红敬. 2002. 广西平鳍鳅科鱼类及其地理分布研究[J]. 信阳师范学院学报(自然科学版),15(2):204-207. [Chen M,Huang N,Li H J. 2002. Study on the fishes of Homalopteridae and their distribution from Guangxi[J]. Journal of Xinyang Teachers College(Natural Science Edition),15(2):204-207.] doi:10.3969/j.issn.1003-0972. 2002.02.022.
封文利,吴志强,黄亮亮,胡祎祥,师瑞丹,丁洋,常显志. 2017. 漓江中游16种常见鱼仔稚鱼形态特征初步研究[J]. 水生态学杂志,38(2):94-100. [Feng W L,Wu Z Q,Huang L L,Hu Y X,Shi R D,Ding Y,Chang X Z. 2017. Morphological characters of 16 larval and juvenile fish species in the middle reaches of Lijiang River[J]. Journal of Hydroecology,38(2):94-100.] doi:10.15928/j.1674-3075.2017.02.015.
广西水产研究所,中国科学院动物研究所. 2006. 广西淡水鱼类志[M]. 第2版. 南宁:广西人民出版社. [Guangxi Fisheries Research Institute,Institute of Zoology,Chinese Academy of Sciences. 2006. Freshwater fishes of Guangxi,China[M]. The 2nd Edition. Nanning:Guangxi Peoples Press.]
胡飞飞,陈新军. 2015. 太平洋褶柔鱼秋生群资源补充量预报模型研究[J]. 广东海洋大学学报,35(6):64-69. [Hu F F,Chen X J. 2015. Study on forecasting model of recruitment for the autumn-spawning group of Todarodes pacificus[J]. Journal of Guangdong Ocean University,35(6):64-69.] doi:10.3969/j.issn.1673-9159.2015.06.012.
高明慧. 2018. 西江来宾段鱼类早期资源及其与环境因子的关系研究[D]. 桂林:桂林理工大学. [Gao M H. 2018. Fish resource in the early life stages and its relation to environmental factors in Laibin section of Xijiang River[D]. Guilin:Guilin University of Technology.] doi:10.27050/d.cnki.gglgc.2018.000072.
匡天旭. 2018. 珠江鮊亚科仔鱼分子鉴定与群落分析[D]. 上海:上海海洋大学. [Kuang T X. 2018. Molecular identification and community structure of Cultrinae larvae in the Pearl River[D]. Shanghai:Shanghai Ocean University.]
李策. 2018. 西江仔鱼种类识别及优势种资源现状研究[D]. 上海:上海海洋大学. [Li C. 2018. Identification of larvae and the status of dominant species resources in the Xi-jiang River[D]. Shanghai:Shanghai Ocean University.] doi: 10.27314/d.cnki.gsscu.2019.000051.
梁晓华,刘杰,雷卫东. 2013. 广西水利水电建设项目鱼类保护措施落实情况及效果分析[J]. 红水河,32(3):35-39. [Liao X H,Liu J,Lei W D. 2013. Analysis on implementation and effectiveness of fish conservation measures for water conservancy and hydropower projects in Guangxi[J]. Hongshui River,32(3):35-39.] doi:10.3969/j.issn. 1001-408X.2013.03.010.
刘飞,林鹏程,黎明政,高欣,王春伶,刘焕章. 2020. 长江流域鱼类资源现状与保护对策[J]. 水生生物学报,43(S1):144-156. [Liu F,Lin P C,Li M Z,Gao X,Wang C L,Liu H Z. 2020. Situations and conservation strategies of fish resources in the Yangtze River basin[J]. Acta Hydrobiologica Sinica,43(S1):144-156.] doi:10.7541/2019. 177.
劉孟凯,潘保柱,黄明海,尹正杰. 2017. 长江上游铜鱼产卵流速特性分析[J]. 水力发电,43(7):5-9. [Liu M K,Pan B Z,Huang M H,Yin Z J. 2017. Flow velocity characteristic analysis for Coreius heterodon spawning in upper reaches of Changjiang River[J]. Water Power,43(7):5-9.]
刘雪飞. 2019. 漂流性鱼卵运动特性研究[D]. 北京:中国水利水电科学研究院. [Liu X F. 2019. Study on the movement characteristics of drifting fish eggs[D]. Beijing:China Institute of Water Resources and Hydropower Research.]
乔晔. 2005. 长江鱼类早期形态发育与种类鉴别[D]. 北京:中国科学院研究生院. [Qiao Y. 2005. Early morphogenesis and species identification of fishes in Yangtze River[D]. Beijing:University of Chinese Academy of Scien-ces.]
帅方敏,李新辉,李跃飞,杨计平,李捷. 2016. 珠江东塔产卵场鳙繁殖的生态水文需求[J]. 生态学报,36(19):6071-6078. [Shuai F M,Li X H,Li Y F,Yang J P,Li J. 2016. Analysis of the ecohydrological demands of bighead carp breeding in the Dongta spawning grounds of the Pearl River,via cross-wavelet analysis[J]. Acta Ecologica Sinica,36(19):6071-6078.] doi:10.5846/stxb201501290244.
谭细畅,李新辉,林建志,李跃飞,毕晔,李捷,王超. 2009a. 珠江肇庆江段鲤早期发育形态及其补充群体状况[J]. 大连水产学院学报,24(2):125-129. [Tan X C,Li X H,Lin J Z,Li Y F,Bi Y,Li J,Wang C. 2009a. Early morphogenesis and larval resources of common carp at Zhaoqing section in the Pearl River[J]. Journal of Dalian Fishe-ries University,24(2):125-129.] doi:10.3969/j.issn.1000- 9957.2009.02.007.
谭细畅,李新辉,陶江平,李捷,王超,罗建仁,赖子尼. 2007. 西江肇庆江段鱼类早期资源时空分布特征研究[J]. 淡水渔业,37(4):37-40. [Tan X C,Li X H,Tao J P,Li J,Wang C,Luo J R,Lai Z N. 2007. Spatial and temporal distribution of larval resources of fishes in Xijiang River[J]. Freshwater Fisheries,37(4):37-40.] doi:10.3969/j.issn.1000-6907.2007.04.008.
谭细畅,李跃飞,王超,李捷,庞世勋,李新辉. 2009b. 珠江江段赤眼鳟早期发育形态及其补充群体状况[J]. 华中农业大学学報,28(5):609-613. [Tan X C,Li Y F,Wang C,Li J,Pang S X,Li X H. 2009b. Early morphogenesis and larval resources of Squaliobarbus curriculus in the Pearl River[J]. Journal of Huazhong Agricultural University,28(5):609-613.] doi:10.3321/j.issn:1000-2421. 2009.05.021.
陶雨薇,王远坤,王栋,吴吉春,倪玲玲. 2018. 三峡水库坝下水温变化及其对鱼类产卵影响[J]. 水力发电学报,37(10):48-55. [Tao Y W,Wang Y K,Wang D,Wu J C,Ni L L. 2018. Assessing water temperature variations and impacts on fish spawning downstream of Three Gorges Dam[J]. Journal of Hydroelectric Engineering,37(10):48-55.] doi:10.11660/slfdxb.20181006.
王丽,朱远生,杨晓灵,梁晓华. 2016. 大藤峡水利枢纽工程设计中的水生态优化措施[J]. 水资源保护,32(3):74-78. [Wang L,Zhu Y S,Yang X L,Liao X H. 2016. Measures for aquatic ecological optimization in design of Datengxia Water Project[J]. Water Resources Protection,32(3):74-78.] doi:10.3880/j.issn.1004-6933.2016.03.014.
王永杰,吕文军,王静,梁春银,王雷,刘哲. 2018. 甘肃康县大鲵自然保护区大鲵资源调查与分析[J]. 甘肃农业大学学报,53(2):23-30. [Wang Y J,Lü W J,Wang J,Liang C Y,Wang L,Liu Z. 2018. Investigation and analysis on resources of the Chinese giant salamander in Kang County Nature Reserve of Gansu Province[J]. Journal of Gansu Agricultural University,53(2):23-30.] doi:10.3969/j.issn.1003-4315.2018.02.004.
吴金明,王芊芊,刘飞,刘春池,王剑伟. 2011. 赤水河四川华吸鳅的早期发育[J]. 四川动物,30(4):527-529. [Wu J M,Wang Q Q,Liu F,Liu C C,Wang J W. 2011. Early development of Sinogastromyzon szechuanensis in the Chishui River[J]. Sichuan Journal of Zoology,30(4):527-529.] doi:10.3969/j.issn.1000-7083.2011.04.002.
熊玉宇,乔晔,刘焕章,谭德清. 2008. 犁头鳅早期发育[J]. 水生生物学报,32(3):421-433. [Xiong Y Y,Qiao Y,Liu H Z,Tan D Q. 2008. Early development of Lepturichthys fimbriata[J]. Acta Hydrobiologica Sinica,32(3):421-433.] doi:10.3321/j.issn:1000-3207.2008.03.023.
杨骏,郭延蜀. 2013. 中国四川省华吸鳅属鱼类一新种(鲤形目,爬鳅科)[J]. 动物分类学报,38(4):895-900. [Yang J,Guo Y S. 2013. A new species of the genus Sinogastromyzon Fang from Sichuan Province,China(Cypriniformes,Balitoridae)[J]. Acta Zootaxonomica Sinica,38(4):895-900.]
易伯鲁,梁秩燊,余志堂. 1988. 葛洲坝水利枢纽与长江四大家鱼[M]. 武汉:湖北科学技术出版社. [Yi B L,Liang Z C,Yu Z T. 1988. Gezhouba water control project and four famous fishes in Yangtze River[M]. Wuhan:Hubei Science and Technology Press.]
余为,陈新军,易倩,李日嵩. 2013. 北太平洋柔鱼早期生活史研究进展[J]. 上海海洋大学学报,22(5):755-762. [Yu W,Chen X J,Yi Q,Li R S. 2013. Review on the early life history of neon flying squid Ommastrephes bartramii in the North Pacific[J]. Journal of Shanghai Ocean University,22(5):755-762.]
周春生,梁秩燊,黄鹤年. 1980. 兴修水利枢纽后汉江产漂流性卵鱼类的繁殖生态[J]. 水生生物学集刊,(2):175-188. [Zhou C S,Liang Z S,Huang H N. 1980. Ecological features of the spawning of certain fishes in the Hanjiang River after the donstruction of dams[J]. Acta Hydrobiologica Sinica,(2):175-188.]
Dudley R K,Platania S P. 1999. Imitating the physical properties of drifting semibuoyant fish(Cyprinidae) eggs with artificial eggs[J]. Journal of Freshwater Ecology,14(4):423-430. doi:10.1080/02705060.1999.9663700.
Hilborn R,Ovando D A. 2014. Reflections on the success of traditional fisheries management[J]. ICES Journal of Marine Science,71(5):1040-1046. doi:10.1093/icesjms/fsu034.
Liu S W,Chen X Y,Yang J X. 2010. Two new species and a new record of the genus Sinogastromyzon(Teleostei:Balitoridae) from Yunnan,China[J]. Environmental Biology of Fishes,87:25-37. doi:10.1007/s10641-009-9554-8.
Mu H X,Li M Z,Liu H Z,Cao W X. 2014. Analysis of fish eggs and larvae flowing into the Three Gorges Reservoir on the Yangtze River,China[J]. Fisheries Science,80:505-515. doi:10.1007/s12562-014-0729-7.
Rand P S,Fukushima M. 2014. Estimating the size of the spawning population and evaluating environmental controls on migration for a critically endangered Asian salmonid,Sakhalin taimen[J]. Global Ecology and Conservation,2:214-225. doi:10.1016/j.gecco.2014.09.007.
Roa-Ureta R H,Henríquez J,Molinet C. 2020. Achieving sustainable exploitation through co-management in three Chilean small-scale fisheries[J]. Fisherise Research,230:105674. doi:10.1016/j.fishres.2020.105674.
Wang X Z,Liu H Z. 2005. Phylogenetic relationships of the Chinese cyprinid genus Rhinogobio Bleeker(Teleostei:Cyprinidae) based on sequences of the mitochondrial DNA control region,with comments on character adaptations[J]. Hydrobiologia,532:215-220. doi:10.1007/s10750-004-8773-4.
Zhang H,Jari? I,Roberts D,He Y F,Du H,Wu J M,Wang C Y,Wei Q W. 2020. Extinction of one of the world?s lar-gest freshwater fishes:Lessons for conserving the endangered Yangtze fauna[J]. The Science of the Total Environment,710:136242. doi:10.1016/j.scitotenv.2019.136 242.
(責任编辑 兰宗宝)
