中国不同生活型植物细根碳氮磷化学计量特征及其影响因子
2021-09-12孙佳祺熊维彬李永奇蔡天润余华
孙佳祺 熊维彬 李永奇 蔡天润 余华
摘要:[目的]為了探讨中国不同生活型植物细根碳(C)、氮(N)、磷(P)含量及其生态化学计量比对外界生活环境因子的响应趋势, 印证细根在“温度-植物生理假说”、“生长速率假说”“不同生活型植物种群的营养元素利用策略”等方面的规律。[方法]该研究通过收集已发表且可以获取的有关中国植物细根研究的100余篇文献, 从中提取出细根C、N、P及其比值等相关数据, 总结分析了不同生活型植物细根化学计量特征与年均降水量、年均温的关系。[结果](1)中国陆地不同生活型植物细根的C、N、P含量的平均值分别为415.8mg/g、9.1mg/g、0.9mg/g,细根的C:N、C:P和 N:P分别为63.5、903.3、15.3。(2)三种生活型植物细根在C含量及C∶P比上有显著性差异(P<0.05),均表现为草本显著低于乔木和灌木;而N、P含量及C∶N和N:P比无显著性相关性差异(P>0.05)。(3)不同生活型植物细根C含量与P含量极显著负相关(P<0.01),与C:N则显著正相关(P<0.05),与C:P、N:P则极显著正相关(P<0.01);N含量与P含量和N:P均极显著正相关(P<0.01),与C:N、C:P则均极显著负相关(P<0.01);P含量与C:N、C:P、N:P均极显著负相关(P<0.01),C:N与C:P则呈极显著正相关(P<0.01);C:P与N:P则极显著正相关(P<0.01)。(4)细根C、P含量随着年均温的升高而降低,细根C:P、N:P随着年均温的升高而升高;细根C、N、P含量随着年均降水量的升高而降低, 细根C:P、N:P随着年均降水量的升高而升高。[结论]不同生活型植物细根的生态化学计量特征载着植物生存环境的变化信息,从而为探索全球变化对植物内在机制的影响提供理论依据。
关键词:细根;碳氮磷生态化学计量特征; 生活型; 年均温, 年均降水量;
Abstract[objective]In order to investigate the responses and spatial patterns of carbon (C), nitrogen (N) and phosphorus (P) in fine roots of different plant life forms in China to environmental factors, and confirm the rules of "temperature-plant physiological hypothesis" and "growth rate hypothesis" and "nutrient utilization strategies of different plant groups" in fine roots. [method] This paper collection has been published and can get on Chinese land plants study of more than 100 articles, fine root extract content of C, N, P element fine root and its related data, The relationship between the content of C, N, P and their stoichiometric ratio in fine roots of different life forms and average annual precipitation and average annual temperature was summarized and analyzed.[result] The results showed as follows: (1) The mean contents of C, N and P in fine roots of different life forms in China were 415.8mg/g, 9.1mg/g and 0.9mg/g, respectively. The values of C:N, C:P and N:P in fine roots were 63.5, 903.3 and 15.3, respectively.(2) There were significant differences in C content and C∶P ratio in fine roots of three life forms (P<0.05), which were significantly higher in trees and shrubs than in herbs; There was no significant correlation between N and P contents and C∶N and N:P ratios (P>0.05).(3) The content of C in fine roots of different life forms was significantly negatively correlated with P content (P<0.01), significantly positively correlated with C:N content (P<0.05), and significantly positively correlated with C:P, N:P (P<0.01).N content was significantly positively correlated with P content and N:P content (P<0.01), and negatively correlated with C:N and C:P (P<0.01).P content was significantly negatively correlated with C:N, C:P and N:P (P<0.01), and C:N was significantly positively correlated with C:P (P<0.01).The correlation between C:P and N:P was significantly positive (P<0.01).(4) The content of C and P in fine roots decreased with the increase of annual mean temperature, and the content of C:P, N:P in fine roots increased with the increase of annual mean temperature. The contents of C, Nand P in fine roots decreased with the increase of mean annual precipitation, while C:P, N:P in fine roots increased with the increase of mean annual precipitation.[conclusion] The ecological stoichiometric characteristics of fine roots of different plant life forms carry information about the changes of plant living environment, thus providing a theoretical basis for exploring the effects of global changes on the internal mechanisms of plants.
Key words Fine root,Ecological stoichiometry of carbon, nitrogen and phosphorus,The life,annual average temperature, annual average precipitation;
引言:
生态化学计量学(ecological stoichiometry)的含义:通过分析生态系统中各物质元素供需平衡关系,主要包括碳(C)、 氮(N)、磷(P)含量及其比率等化学计量特征,来统一不同尺度、不同生物种群和不同研究领域的综合方法[1]。C是生命的骨架元素[2]与结构性物质[3],通过光合作用同化的C是植物生理生化过程的能量来源。N 是植物生长的重要限制元素,参与光合作用、凋落物降解等生态过程[4]。P 是遗传物质核酸、能量储存物质ATP(adenosine triphosphate)和各种蛋白质的重要组成元素,在一些生境中为植物生长的限制元素[5]。C:N和C:P反映植物对养分的利用效率,N:P表征生态系统的元素限制。C、N、P含量及其计量比之间存在复杂的耦合作用,不仅参与地球生物化学循环,而且为植物的生长发育提供必须的营养元素,在生态系统物质流动和能量循环以及多元素平衡过程中共同发挥着重要作用[6,7]。
细根(Fineroot)通常是指植物根系中直径≤2mm的部分[8,9],是根系系统中最活跃和最敏感的部分,是植物获取水分和营养元素的主要器官之一[10]。在生态系统的物质循环、能量流动和元素供需平衡中扮演着重要的角色[11]。探究细根C、N、P生态化学计量特征为揭露生态系统养分循环对全球气候变化的响应具有重要的理论意义和应用价值[12]。
目前,对于生态化学计量学的研究大多集中于森林[13]与草原系统[14],或者是生态系统的单一群落层次[15],对大尺度生态化学计量学的认识比较短缺。然而,不同种群植物往往由于地理区域、气候条件(年均温、年均降水量)的差异,导致植物的营养器官在结构和功能上存在一定差异,植物C、N、P的分配利用也呈现多元化。同时,中国幅员辽阔,南北跨纬度、东西跨经度近50度,各地年均温、年均降水量差异较大,具有世界独一无二的生物群区。因此,研究不同生活型植物细根C、N、P及其比值与年均温、年均降水量的响应趋势, 进一步完善有关中国植物细根元素动态平衡的研究。
1 材料和方法
1.1数据来源
通过查阅《中国植被》并借助“中国知网”、“维普网”、“万方”等中文期刊数据库,以“细根”、“化学计量”、“养分”、“碳、氮、磷”为关键词检索, 搜索已发表的相关中国植物根系研究的100余篇文献。通过筛选整理得到不同生活型植物细根的C、N、P含量及其化学计量比。同时记录研究地的经纬度、海拔、年均温、年均降水量及其生活型等相关数据。共收集不同生活型植物细根C、N、P数据144个(其中乔木68个, 灌木52个,草本24个) 数据来自50个研究站点, 研究区覆盖了中国的大部分地区(84°20′-128°53′E,19°31′–47°10′N), 年平均气温-0.3-24℃, 年均降水量65.5–2583mm, 涉及森林、草原、荒漠、滩涂、沼泽等生态系统。
1.2数据分析
采用单因素方差分析(one-way ANOVA)分析三种生活型植物细根C、N、P含量及其计量比的差异;在分析前先进行方差齐性检验,方差齐性时,运用最小显著法(LSD)进行多重比较检验;方差非齐性时,进行Tamhane,A.C.检验。利用皮尔逊相关分析(Pearson)对细根各指标进行相关分析。以年均温、年均降水量为自变量, 细根 C、N、P含量及其计量比作为因变量,进行一元线性回归拟合分析。运用Excel 2019进行数据整理及绘图,SPSS 24.0進行统计分析。
2 结果
2.1 不同生活型植物细根化学计量特征差异
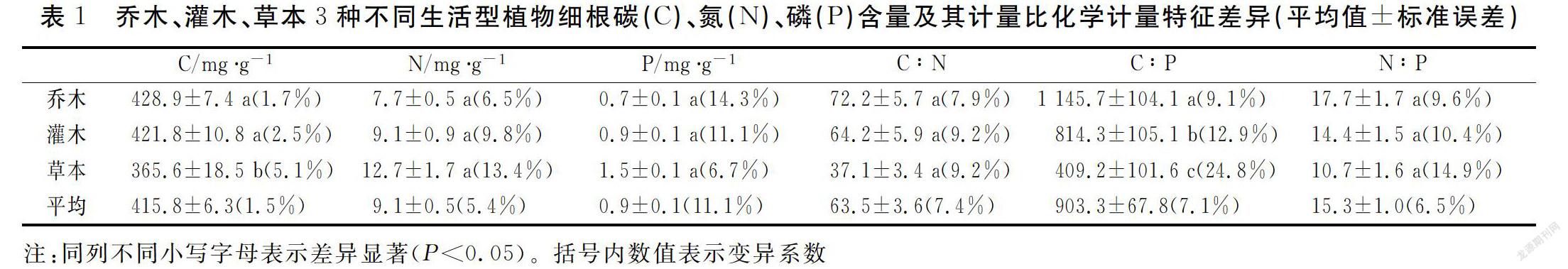
本研究中,三类生活型植物细根的C、N、P含量的平均值分别为415.8mg/g、9.1mg/g、0.9 mg/g,细根的C:N、C:P和 N:P分别为63.5、903.3、15.3。三类生活型植物细根在C含量及C∶P化学计量比值上有显著性差异(P<0.05),在N、P含量及C∶N和N;P化学计量比值上无显著相关性差异(P>0.05)
通过对不同生活型植物细根C、N、P含量及其化学计量比特征差异进行分析,结果如图(1):三种生活型植物细根在C含量以及C∶N、C∶P和N∶P化学计量比值上存在乔木>灌木>草本的规律,三种生活型植物细根在N、P含量上存在草本>灌木>乔木的规律。总体而言,乔木的C含量、C:N、C:P和N:P均大于灌木和草本, 草本植物的N、P含量大于乔木和灌木。
2.2.不同生活型植物细根化学计量特征间相关性
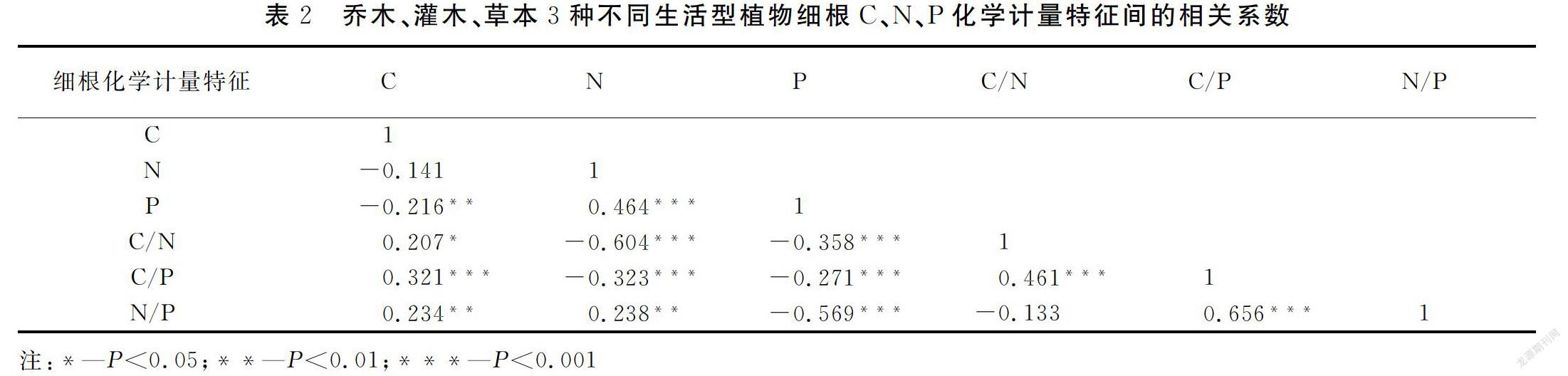
乔木、灌木、草本三种不同生活型植物细根C含量与P含量极显著负相关(P<0.01),与C:N则显著正相关(P<0.05),与C:P、N:P则极显著正相关(P<0.01);N含量与P含量和N:P均极显著正相关(P<0.01),与C:N、C:P则均极显著负相关(P<0.01);P含量与C:N、C:P、N:P均极显著负相关(P<0.01),C:N与C:P则呈极显著正相关(P<0.01);C:P与N:P则极显著正相关(P<0.01)。
2.3细根化学计量特征的影响因子
细根C含量总体随着年均温的升高而降低,从回归斜率可以看出,灌木细根的C含量比草本细根的C含量对年均温的响应更敏感(如图1-a);细根P含量总体随着年均温的升高而降低,从回归斜率可以看出,草本细根的N含量比乔木细根的N含量对年均温的响应更敏感(如图1-c)。细根C含量总体随着年均降水量的升高而降低,从回归斜率可以看出,草本细根的C含量比灌木细根的C含量对年均降水量的响应更敏感(如图2-g);细根N含量总体随着年均降水量的升高而降低(如图2-h);细根P含量总体随着年均降水量的升高而降低(如图2-i)。
细根C:P总体随着年均温的升高而升高(如图1-e);细根N:P随着年均温的升高而升高(如图1-f)。细根C:P随着年均降水量的升高而升高(如图2-k);细根N:P随着年均降水量升高而升高(如图2-i)。
3讨论
3.1不同生活型植物细根化学计量比特征及其差异
本研究中,细根的C、N、P含量平均值分别为 415.8mg/g、9.1mg/g和0.9mg/g。植物细根C含量明显低于全球(436.9 mg/g)和中国(473.9mg/g)植物的平均值,其原因可能是由于前人统计的大量样本中用于细根C含量测定的样本量较少;且本研究中,温带森林研究对象较多,而缺少亚热带森林。 Yuan等[16]对全球细根的研究表明,细根的N含量在9.9-11.2mg/g之间, P含量在0.55-0.85mg/g之间。本研究发现,细根N含量小于全球平均水平,P含量大于全球平均水平。经分析发现,本文研究对象中缺少N含量高、P含量低的热带森林,而多N含量低、P含量高的温带森林和草原,致使本研究结果中N、P含量与全球对比存在出入[17]与马玉珠等人研究结果类似。
通过研究发现,不同生活型植物细根的N、P含量以及C:N、C:P和N:P均存在较大的差异,表明了不同植物细根受到的养分限制不同,这不但与生境提供的N和P养分的情况有关,还与植物自身的结构特点、生长节律以及摄取养分的方式息息相关[18]。本研究中,3种生活型植物细根在C:N和C:P上存在乔木>灌木>草本的规律,且乔木的C:N和C:P的比值均远高于灌木和草本,这可被认为乔木植物在生长发育过程中相比于灌木和草本有更加高效的N、P利用效率,生长速度快。其次,乔木和灌木的N:P较为接近,一定程度上表征了乔木和灌木在养分利用策略上存在一定的相似性。其原因可能是乔木和灌木均为木本植物,具有相似的生态学特性,支持朱亮等[19]的研究结果。本研究中还发现,乔木和灌木的N、P含量低于草本,可能是由于草本植物寿命相对较短,需要在相对较短的时间内汲取充足的养分利于植物的生长,而根系作为主要的吸收器官就会在短时间内从土壤中大量吸收N和P提供给植物,保证植物的正常生长发育[20]。
在植物的营养器官上 ,与叶片相比,本研究细根的N、P含量均小于中国陆地植物叶片N、P含量的平均水平 (分别为:18mg/g及 23mg/g),同时也低于澳大利亚植物叶片 N、P含量 (分别为:10mg/g及 0.8mg/g),也验证了Yuan等[16]的研究结果。这是因为植物叶片在进行光合作用的生态过程中,需要投入大量的 N元素来构建光合器官 ,而细根则需要更多富P的rRNA来支持蛋白质的合成,从而也会展示出更低的N:P,这也是细根与叶片的营养元素含量差异较大的因素之一。尽管植物叶片和细根对N、P的吸收和分配具有明显的保守性,不同生活型植物叶片和细根之间N:P仍然存在显著差异(P<0.001),反映了不同生活型植物对N、P元素的利用存在较大的差异。
3.2不同生活型植物细根化学计量特征间相关关系
C作为植物的结构基础,N、P通常被认为是植物功能的限制性元素,三者相辅相成、共同作用以促进植物生长[21]。本研究中显示,细根N、P含量间呈极显著(P<0.01)的正相关关系,在一定程度上表明功能相关的细根N、P元素之间具有相似的生化途径[22] ,同时在相同生境条件下二者对外界环境因子(年均温、年均降水量)也具有相似的响应特征。
细根的C:N和C:P表征着植物的物质养分循环及其碳同化效率,其N:P体现植物生长对营养元素的供需平衡关系[23]。本研究结果显示,细根N含量与其C:N和C:P极显著负相关(P<0.01),P含量与C:N、C:P和N:P极显著负相关(P<0.01),C:P与C:N和N:P极显著正相关(P<0.01)。产生这一结果主要是因为C作为植物的结构基础,亦是相对稳定的结构性元素, 其稳定性要远超于N、P元素,而N、P则是一对具有显著正相关关系的限制性因子,二者在变化上存在P含量要大于N含量的规律,这也支持了其他学者的研究结论[24-29]。
3.3细根化学计量对外界环境因子的响应模式
在全球气候变化背景下,年均温、年均降水量的变化对植物群落动态演化将扮演重要角色,中国从最南端的曾母暗沙往北到黑龙江漠河,南北跨纬度近50度, 从乌苏里江到最西端的帕米尔高原,东西跨经度近60度,幅员辽阔,各地的气候状况和干湿度条件不尽相同。植物的细根C、N、P含量和细根形态对外部环境因子变化的响应程度也有所差别,具体表现为:不同类型植物细根根据各自的营养元素利用策略分别做相应的形态调整。
Reich和Oleksyn[30]的研究发现,在全球范围内,植物叶片的N、P含量随着纬度降低和年平均温度升高而降低。Han等[31]和任书杰等[32]在中国的研究也有相同的发现,同时,在本研究中细根的N、P含量随年均温的升高而降低的规律也与前人研究结果一致,该结果与“温度-植物生理假说”相符。“溫度-植物生理假说”认为: 植物在生长旺季光合作用旺盛,需要大量的N元素用于叶绿素和光合蛋白等光合器官的构建以及P元素参与合成的ATP和酶 [33] 。植物的代谢效率和生长速度与N、P含量成正比,因此需要更高的N、P含量来支持植物的光合作用与所需养分的合成,以此抵消了高纬、高寒地区的低温对植物体生物化学反应速率降低的效应[34,35], 这是植物对外界环境变化的一种适应策略。
通过对比本研究中年均温和年均降水量两个气候因子对细根C、N、P含量的影响,得出温度比降水对细根营养元素的影响更大,但不同生活型植物细根对降水变化的响应幅度也存在一定的差异[36],如在干旱少雨的戈壁荒漠地区,大气降水是主要的水分来源,降水的变化幅度对植物种群多样性、生产力、植被动态变化以及一系列重要生态系统功能将产生深远的影响[37]。此外,降水的波动会通过土壤水分的变化在不同程度上影响植物的营养元素利用策略和对外界生活环境的适应策略[38],例如,乔木、灌木细根由于能从更深的土层中获取水分和养分,对降水变化响应的敏感性较弱;而草本植物对年际降水变化的响应比木本科植物更敏感,有关不同降水条件下不同生活型植物生长响应的研究较少。因此,探究不同生活型植物细根如何响应降水变化,对全球不同植物类型细根动态和植被的管理保护与可持续发展具有重要意义。
结论:
综上所述,(1)中国不同生活型植物细根的C、N、P含量的平均值分别为415.8mg/g、9.1mg/g、0.9mg/g,细根的C:N、C:P和 N:P分别为63.5、903.3、15.3。 (2)三种生活型植物细根在C含量及C∶P比上有显著性差异(P<0.05),均表现为草本显著低于乔木和灌木;而N、P含量及C∶N和N:P比无显著性相关性差异(P>0.05)。(3)不同生活型植物细根C含量与P含量极显著负相关(P<0.01),与C:N则显著正相关(P<0.05),与C:P、N:P则极显著正相关(P<0.01);N含量与P含量和N:P均极显著正相关(P<0.01),与C:N、C:P则均极显著负相关(P<0.01);P含量与C:N、C:P、N:P均极显著负相关(P<0.01),C:N与C:P则呈极显著正相关(P<0.01);C:P与N:P则极显著正相关(P<0.01)。(4)细根C、P含量随着年均温的升高而降低,细根C:P、N:P随着年均温的升高而升高;细根C、N、P含量随着年均降水量的升高而降低, 细根C:P、N:P随着年均降水量的升高而升高。本文通过对中国陆地不同生活型植物细根C、N、P元素含量及其比值等相关数据的收集和分析, 印证了细根在“温度-植物生理假说”及“生长速率假说”和“不同植物类群的营养元素利用策略”等方面的规律, 这对了解中国植物细根营养元素的利用策略和对外界生活环境的适应策略提供了数据支持。
参考文献
[1]. Michaels, Anthony F . The Ratios of Life.[J]. Science, 2003.
[2]. 邓斌.高寒草地不同演替阶段植被变化和土壤碳氮磷的生态化学计量研究[D].兰州:兰州大学草地农业科技学院,2012.
[3]. AGREN G 1. Stoichiometry and nutrition of plant growth in natural communities [J]. Annual Review of Ecology,Evolution,and Systematics ,2008,39:153-170.
[4]. Lebauer D S, Treseder K K. NITROGEN LIMITATION OF NET PRIMARY PRODUCTIVITY IN TERRESTRIAL ECOSYSTEMS IS GLOBALLY DISTRIBUTED[J]. Ecology, 2008, 89(2):371-379.
[5]. Reich P B, Wright O I J . Leaf Phosphorus Influences the Photosynthesis-Nitrogen Relation: A Cross-Biome Analysis of 314 Species[J]. Oecologia, 2009, 160(2):207-212.
[6]. 邬畏. 生态系统氮磷比化学计量特征研究进展[J]. 中国沙漠, 2010, 30(2):296-302.
[7]. Reed S , Cleveland C C , Davidson E A , et al. Patterns in foliar nutrient resorption stoichiometry at multiple scales: controlling factors and ecosystem consequences (Invited)[C]// Agu Fall Meeting. AGU Fall Meeting Abstracts, 2013.
[8]. 张小全,吴可红,Murach D. 树木细根生产与周转研究方法评述. 生态学报,2000,20(5) : 875-883.
[9].Geng Y, Wang L, Jin DM, Liu HY, He JS (2014). Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits. Oecologia, 175, 445–455.
[10].王维奇, 仝川, 贾瑞霞,等. 不同淹水频率下湿地土壤碳氮磷生态化学计量学特征[J]. 水土保持学报, 2010(03):240-244.
[11].Li M , Zheng-Quan W , Yun-Huan C , et al. A review: Factors influencing fine root longevity in forest ecosystems[J]. Chinese Journal of Plant Ecology, 2004, 28(5):704-710.
[12].張鑫, 邢亚娟, 闫国永,等. 细根对降水变化响应的meta分析[J]. 植物生态学报, 2018, 42(002):164-172.
[13].王晶苑, 王绍强, 李纫兰,等. 中国四种森林类型主要优势植物的C:N:P化学计量学特征[J]. 植物生态学报, 2011, 35(006):587-595.
[14].周鹏, 耿燕, 马文红,等. 温带草地主要优势植物不同器官间功能性状的关联[J]. 植物生态学报, 2010.
[15].车应弟, 刘旻霞, 李俐蓉,等. 基于功能性状及系统发育的亚高寒草甸群落构建[J]. 植物生态学报, 2017, 41(11):1157-1167.
[16].Yuan ZY, Chen HYH, Reich PB (2011). Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus. Nature Communications, 2, 344.
[17].马玉珠, 钟全林, 靳冰洁,等. 中国植物细根碳、氮、磷化学计量学的空间变化及其影响因子[J]. 植物生态学报, 2015, 039(002):159-166.
[18].Chen F S, Niklas K J, Zeng D H. Important foliar traits depend on species-grouping: analysis of a remnant temperate forest at the Keerqin Sandy Lands, China[J]. Plant & Soil, 2011, 340(s1-2):337-345.
[19].朱亮, 顾国军, 徐振锋,等. 北亚热带常绿阔叶林乔灌草物种的碳氮磷生态化学计量比[J]. 应用与环境生物学报, 2019, 25(06):1277-1287.
[20].宁志英, 李玉霖, 杨红玲,等. 科尔沁沙地主要植物细根和叶片碳、氮、磷化学计量特征[J]. 植物生态学报, 2017, 041(010):1069-1080.
[21].孙美美, 关晋宏, 岳军伟,等. 黄土高原西部针叶林植物器官与土壤碳氮磷化学计量特征[J]. 水土保持学报, 2017, 31(003):202-208.
[22].GARTEN, Charles T . Correlations between concentrations of elements in plants[J]. Nature, 1976, 261(5562):686-688.
[23].SabineGüsewell. N : P ratios in terrestrial plants: variation and functional significance[J]. 2004, 164(2):243-266.
[24].Wright I J , Reich P B , Cornelissen J H C , et al. Assessing the generality of global leaf trait relationships[J]. New Phytologist, 2005, 166(2).
[25].Reich, PB, Oleksyn, et al. Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes[J]. PROC R SOC B, 2010, 2010,277(1683)(-):877-883.
[26].戚德輝,温仲明,王红霞,等. 2016. 黄土丘陵区不同功能群植物碳氮磷生态化学计量特征及其对微地形的响应. 生态学报, 36(20): 6420-6430.
[27].张慧, 郭卫红,杨秀清,等. 2016. 麻栎种源林叶片碳、氮、磷化学计量特征的变异. 应用生态学报,27(7): 2225-2230.
[28].马任甜, 安韶山, 黄懿梅. 黄土高原不同林龄刺槐林碳、氮、磷化学计量特征[J]. 应用生态学报, 2017, 28(009):2787-2793.
[29].余华, 钟全林, 程栋梁,等. 不同种源刨花楠林下幼苗叶片碳氮磷化学计量特征[J]. 林业科学, 2018, 54(12):22-32.
[30].Reich P B , Oleksyn J . Global Patterns of Plant Leaf N and P in Relation to Temperature and Latitude[J]. Proceedings of the National Academy of ences, 2004, 101(30):11001-11006.
[31].Han W , Fang J , Guo D , et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2).
[32].任书杰, 于贵瑞, 陶波,等. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J]. 环境科学, 2007, 28(012):2665-2673.
[33].Reich P B , Oleksyn J . Global Patterns of Plant Leaf N and P in Relation to Temperature and Latitude[J]. Proceedings of the National Academy of ences, 2004, 101(30):11001-11006.
[34].Weih M, Karlsson PS (2001). Growth response of mountain birch to air and soil temperature: Is increasing leaf-nitrogen content an acclimation to lower air temperature? New Phytologist, 150, 147–155.
[35].Woods HA, Makino W, Cotner JB, Hobbie SE, Harrison JF, Acharya K, Elser JJ (2003). Temperature and the chemical composition of poikilothermic organisms. Functional Ecology, 17, 237–245.
[36].张鑫, 邢亚娟, 闫国永,等. 细根对降水变化响应的meta分析[J]. 植物生态学报, 2018, 42(002):164-172.
[37].Loarie, Scott, R, et al. The velocity of climate change.[J]. Nature, 2009.
[38].王军锋, 张丽华, 赵锐锋,等. 荒漠草原区不同生活型植物生长对降水变化的响应[J]. 应用生态学报 2020年31卷3期, 778-786页, MEDLINE ISTIC PKU CSCD CA BP, 2020.
