大豆鲜荚籽粒上位性QTL及其互作效应分析
2021-09-11陈士亮孙亚倩邵振启李文龙孔佑宾杜汇李喜焕张彩英
陈士亮, 孙亚倩, 邵振启, 李文龙, 孔佑宾,杜汇, 李喜焕*, 张彩英
(1.河北农业大学农学院, 华北作物改良与调控国家重点实验室, 河北 保定 071001;2.河北农业大学生命科学学院, 河北 保定 071001)
大豆[Glycinemax(L.) Merr.]是重要的粮食与油料作物,籽粒富含蛋白、脂肪、异黄酮、氨基酸等多种营养物质,在人们膳食结构中占有不可替代的重要地位[1-2]。同时,大豆还可进行鲜荚采摘,作为鲜食毛豆丰富人们餐桌[3-4]。据统计,鲜食毛豆单位面积经济效益约为普通粒用大豆的2倍以上,因而对于提高大豆经济效益具有重要意义。然而,鲜食大豆产量与鲜荚、鲜粒性状关系密切。大豆籽粒产量与其荚长性状呈现极显著正相关[4];改良大豆荚长和荚宽性状不仅有利于增大籽粒,并且有利于提高其单位面积产量[5]。并且,鲜荚和籽粒性状是鲜食大豆品种审定和出口的重要参考依据。因此,改良大豆鲜荚和籽粒相关性状,对于鲜食大豆品种审定、产品出口和经济效益提升均具有重要意义[6]。
目前,已有少数学者针对大豆荚粒相关性状开展研究。王珍[7]利用大豆重组自交系(recombinant inbred line, RIL)获得多个控制二粒荚长、荚宽、粒长和粒宽加性数量性状位点(quantitative trait loci,QTL),分别位于9条染色体(4、5、6、7、8、10号染色体等)上。赵晋铭等[8]分析大豆RIL群体百粒鲜重,获得1个位于18号染色体,可解释7.64%~12.7%表型变异加性QTLs。牛远[9]鉴定了257份大豆资源籽粒相关性状,获得多个控制粒长、粒宽、粒厚加性QTLs。Kenichiro等[10]鉴定大豆RIL群体单株荚数、每荚粒数和单粒重等,获得4个稳定QTLs。由此可见,尽管目前已有少数关于大豆荚粒相关性状遗传位点研究报道[11-16],但多是利用成熟籽粒针对单一加性QTLs进行研究,很少有利用鲜荚和鲜粒、针对上位性QTLs及其互作效应开展研究的报道。
然而,上位性QTLs及其互作效应作为数量性状遗传结构重要组成部分,其对复杂数量性状的影响与简单性状相比更为重要,因而在育种工作中的作用不容忽视[17-18]。由于自花授粉作物较异花授粉作物具有较强的非等位基因之间的互作,因而若检测到加性×加性上位性互作效应,则可以通过人工选择实现其稳定遗传,进而产生额外的遗传收益[19];并且,在上位性效应存在时,仅考虑加性效应将会严重影响高世代材料的选择效率[19]。有学者在研究水稻种子活力相关性状时发现,对于水稻种子发芽率、苗长和干物重而言,上位性效应的作用高于加性效应[20]。另外,对于异花授粉作物而言,上位性效应还是其杂种优势形成的重要遗传基础。可见,在研究数量性状单个加性QTL的同时,挖掘控制其上位性QTL及其互作效应对于今后实现数量性状精准遗传改良具有重要意义。
鉴于此,本研究利用大豆重组自交系群体,2年4种不同环境条件下分别鉴定其鲜荚、鲜粒共8个相关性状,在前期已明确其在鲜荚、鲜粒等相关性状存在广泛遗传变异,并已揭示其在不同环境条件下存在许多一致性加性主效QTLs基础上,继续挖掘控制其鲜荚籽粒性状上位性QTLs,并分析其互作效应,对今后结合利用加性和上位性QTLs,实现大豆鲜荚籽粒性状分子遗传改良以及进一步解析大豆荚粒性状分子遗传机制具有重要意义。
1 材料与方法
1.1 供试材料
以大豆重组自交系群体(C813×垦农7号,193个家系)为材料,由河北农业大学大豆遗传育种课题组构建并提供。
1.2 供试大豆RIL群体田间种植与管理
供试大豆重组自交系群体分别于2019、2020年种植于河北省4种环境条件下(E1~E4),以鉴定供试群体在不同年度、不同地点的遗传变异,其中E1为2019年保定环境(河北农业大学作物育种中心)、E2为2020年保定环境(河北农业大学作物育种中心)、E3为2020年文安环境(廊坊文安)、E4为2020年鹿泉环境(石家庄鹿泉)。从地理位置看,鹿泉试点较保定试点偏西偏南、文安试点较保定试点偏东,4个试点在温度、水分和土壤地力等方面存在一定差异。4个试点均采用完全随机区组试验设计,行长3 m,行距0.5 m,3次重复,田间管理同一般大田管理方法。
1.3 供试大豆RIL群体鲜荚籽粒相关性状测定
供试大豆RIL群体生长至R6~R7期,参照国家鲜食大豆区域试验调查方法并结合前人研究进展[7, 11],分别检测供试群体鲜荚与籽粒相关性状,具体包括标准二粒荚长(fresh pod length, FPL)、二粒荚宽(fresh pod width, FPW)、鲜荚重量(fresh pod weight, FPWT)、鲜粒重量(fresh seed weight, FSWT)、鲜粒长度(fresh seed length, FSL)、鲜粒宽度(fresh seed width, FSW)、长宽比(fresh seed length/width, FSLW)、鲜粒面积(fresh seed area, FSA)共8个性状。
1.4 供试RIL群体上位性QTLs及其互作效应分析
利用供试大豆RIL群体上述8个鲜荚籽粒相关性状,结合前期已构建的群体单核苷酸多态性(single nucleotide polymorphisms, SNP)标记遗传连锁图谱(含2 234个SNP标记),采用IciMapping V4.2软件MET模块[21],将不同环境设为因子,通过完备区间作图法,对8个性状进行上位性QTLs及其互作效应分析,其中检测步长设为5.0 cM,若临近QTL间的遗传距离小于5.0 cM,则将其认定为相同QTL;随后,参照McCouch等[22]提出的QTL命名方法进行命名。
2 结果与分析
2.1 大豆鲜荚籽粒上位性QTLs及其染色体分布
通过分析供试大豆RIL群体8个鲜荚籽粒相关性状上位性QTLs(表1)发现,共获得321对鲜荚籽粒“加性×加性”上位性QTLs,涉及大豆所有染色体,其中以13号染色体分布的上位性QTLs数量最多,其次为6号染色体,而以1号和16号染色体分布数量最少。由此可见,控制大豆鲜荚籽粒的上位性QTLs数量较多,且在染色体上的分布范围较广,除以往报道的单个加性QTLs外,“加性×加性”上位性QTLs在大豆鲜荚籽粒形成和发育中也发挥着重要作用。进一步分析上述测试性状检测到的上位性QTLs还发现,以鲜粒长宽比定位到的上位性QTLs最多,其次为鲜粒宽度,而以鲜荚宽度定位到的上位性QTLs最少。

表1 大豆鲜荚籽粒上位性QTLs染色体分布Table 1 Distribution of epistatic QTLs of fresh pod and seed related traits on soybean chromosomes
另外,上述上位性QTLs互作关系如图1所示,可以看出,大豆鲜荚籽粒性状存在复杂的QTLs互作网络,并以7号、13号染色体与其他染色体间的互作关系较多,其中7号染色体与其余18条染色体间存在互作关系(除3号和17号染色体外),而13号染色体则与除1号和19号染色体外的18条染色体存在互作关系。由此推测,7号和13号染色体可能在大豆鲜荚和籽粒形成与发育过程中处于较为重要的“核心”位置,其通过与较多的染色体间的互作来影响鲜荚和籽粒形成。

图1 大豆鲜荚籽粒性状上位性QTLs染色体互作网络Fig.1 Interaction network of epistatic QTLs associated with soybean fresh pod and seed related traits
2.2 大豆鲜荚籽粒上位性QTLs效应分析
通过分析供试大豆群体鲜荚籽粒性状上位性QTLs遗传效应(表2)发现,在定位到的321对上位性QTLs中,有144对上位性QTLs为正向效应(加性效应为正值),表明两基因座位间的互作基因型与具有正向效应的加性基因型作用方向相同;有177对上位性QTLs为负向效应(加性效应为负值),说明两基因座位间的互作基因型与具有正向效应的加性基因型作用方向相反。在144对正向效应QTLs中,有37对QTLs效应为“++”型,70对效应为“+-”型,另有37对效应表现为“--”型;而在177对负向效应QTLs中,有45对效应为“++”型,99对效应为“+-”型,33对效应为“--”型。
同时分析不同荚粒相关性状连锁QTLs(表2)发现,控制鲜荚长度的36对上位性QTLs中,有19对表现为正向效应,17对表现为负向效应,其中8对为“++”型,22对为“+-”型,6对为“--”型;控制鲜荚宽度的8对上位性QTLs中,2对表现为正向效应,均为“--”型,6对表现为负向效应。控制鲜荚重量的16对QTLs中有6对表现为正向效应,10对表现为负向效应;控制鲜粒长度的52对QTL中有21对正向效应,31对负向效应;鲜粒宽度的68对QTLs中,29对表现为正向效应,39对表现为负向效应。上述上位性QTLs及其遗传效应分析可为今后改良大豆荚粒相关性状提供一定参考。
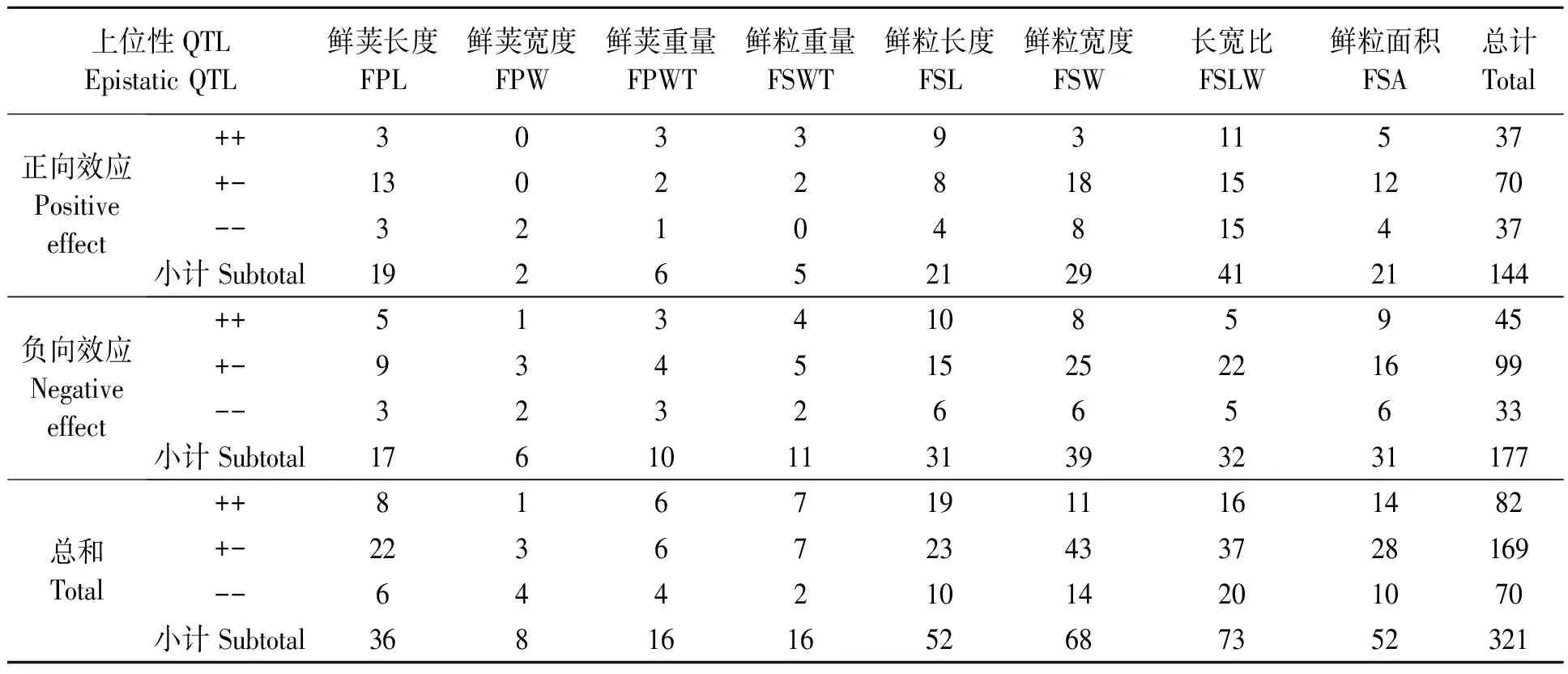
表2 大豆鲜荚籽粒性状上位性QTLs遗传效应分析Table 2 Genetic effect analysis of epistatic QTLs controlling soybean fresh pod and seed related traits
2.3 大豆鲜荚籽粒性状“一因多效”上位性QTLs分析
2.3.1“一因多效”上位性QTLs染色体分布与效应分析 通过分析不同鲜荚籽粒性状定位到的“加性×加性”上位性QTLs(表3)发现,检测到34对“一因多效”上位性QTLs,其贡献率分布在1.98%~6.58%之间,且“加性×加性”上位性QTLs间互作贡献率为1.89%~4.85%(PVE-AA),明显高于其与环境互作间的贡献率(PVE-AAE),说明这些上位性QTLs主要由遗传因素决定,而环境因素影响较小。
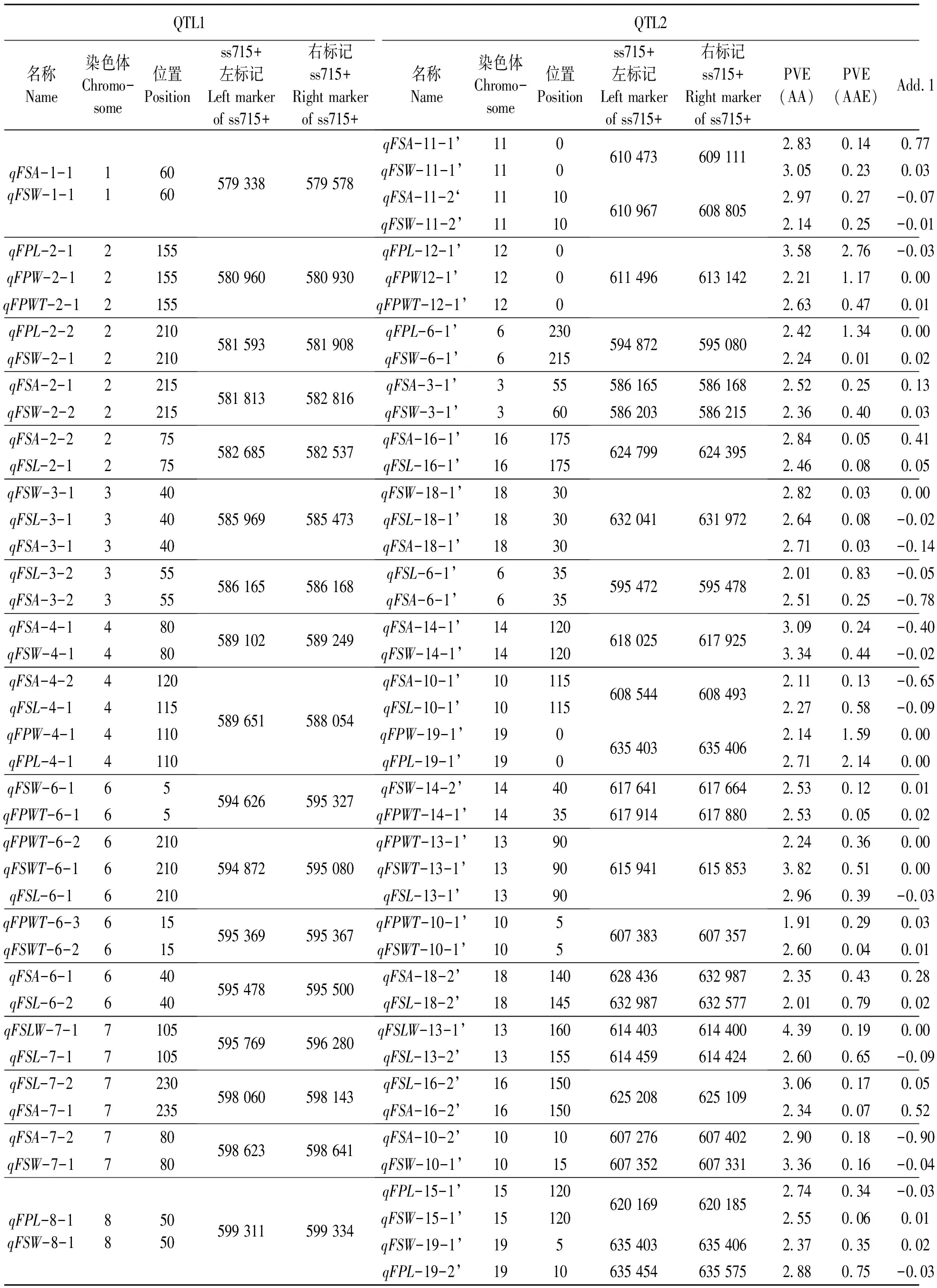
表3 大豆鲜荚籽粒性状一因多效上位性QTLsTable 3 Pleiotropism epistatic QTLs associated with soybean fresh pod and seed related traits
同时发现,在上述34对“一因多效”QTLs中,有23对QTLs定位区间完全一致,11对QTLs定位区间距离较近(< 5 cM),涉及18条染色体,并以13号染色体数量最多,该染色体分别与6号、7号、8号、11号、12号、13号、14号、15号和16号染色体发生互作;6号染色体存在6对“一因多效”QTLs,其分别与2号、3号、10号、13号、14号和18号染色体发生互作。可见,由于大豆鲜荚籽粒相关性状间存在一定相关性,这些相关性不仅与单个加性QTLs的一因多效有关,而且与上位性QTLs的一因多效密切相关。上述一因多效位点的发现可为今后进一步揭示大豆荚粒性状间的相关关系提供分子依据。
另外还发现,上述34对“一因多效”上位性QTLs中,有12对QTLs效应值为正值,另有22对QTLs效应值为负值;并且,上述“一因多效”上位性QTLs在不同性状间的效应值同时为正值或负值,一方面揭示了上述性状间的正相关关系,同时为今后实现上述性状同步遗传改良提供了依据。
2.3.2“一因多效”上位性QTLs互作方式分析
在上述34对“一因多效”QTLs中,包含6组“一对多”上位性QTLs互作方式(图2)和16组“一对一”上位性QTLs互作方式。在6组“一对多”上位性QTLs中,包括2组“一对三”上位性QTLs和4组“一对二”上位性QTLs,其中8号染色体(ss715599311~ss715599334)与15号染色体(ss715620169~ss715620185)和19号染色体2个区间(ss715635403~ss715635406、ss715635454~ss715635575)存在互作;11号染色体(ss715610473~ss715609111)与1号染色体(ss715579338~ss715579578)、15号染色体2个区间(ss715620881~ss715620795、ss715621359~ss715621034)存在互作。
同时,1号染色体(ss715579338~ss715579578)与11号染色体2个区间(ss715610473~ss715609111、ss715610967~ss715608805)存在互作;4号染色体(ss715589651~ss715588054)与10号(ss715608544~ss715608493)、19号染色体(ss715635403~ss715635406)存在互作;6号染色体(ss715594872~ss715595080)与2号(ss715581593~ss715581908)、13号染色体(ss715615941~ss715615853)存在互作;19号染色体(ss715635403~ss715635406)与4号(ss715589651~ss715588054)、18号染色体(ss715632987~ss715632577)存在互作关系。
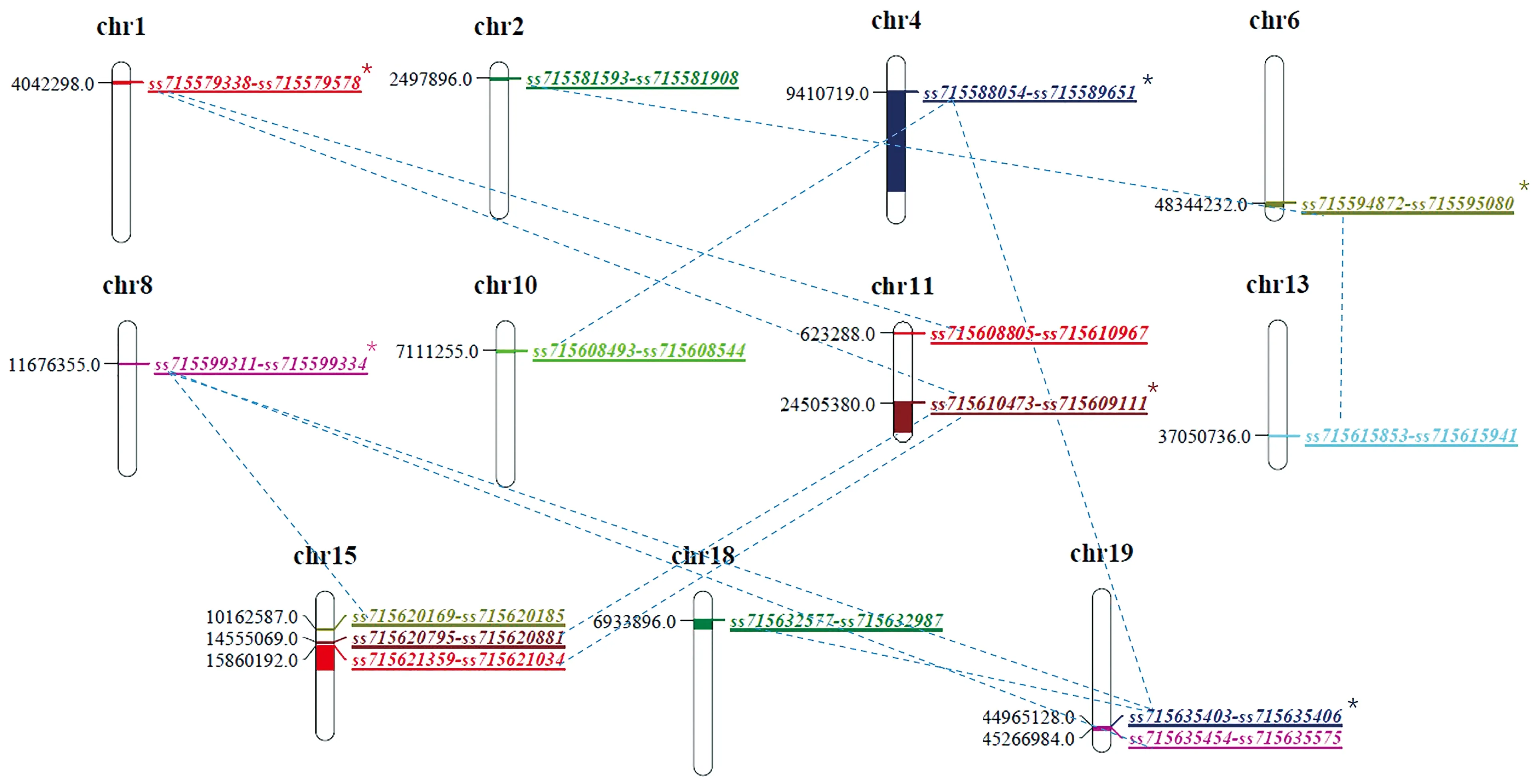
注:*为“一对多”类型上位性QTL。Note: * indicates the “single to multiple” QTL interaction mode.图2 大豆鲜荚籽粒性状“一因多效”上位性QTLs互作分析Fig.2 Interaction analysis of pleiotropism epistatic QTLs associated with soybean fresh pod and seed related traits
3 讨论
荚粒性状对于鲜食大豆品种审定、产品出口以及产量和品质形成等至关重要,而且是普通粒用大豆产量性状重要构成因素,因而已成为大豆育种重要目标性状。大豆荚长、荚宽、荚重、籽粒长度、宽度等性状与产量关系密切[23];并且,荚长、荚宽、荚重和鲜百粒重等性状间存在显著相关[24]。然而,大豆荚粒性状属于多基因控制数量性状,因而发掘控制其遗传位点成为实现性状分子遗传改良重要前提。
据统计,有关大豆不同荚粒相关性状遗传位点发掘研究最早可追溯到20世纪初,到目前为止,已报道的10余个荚粒相关性状加性QTLs约400多个,分布范围涉及大豆所有染色体。截至2018年,国内外已报道的154个大豆荚粒性状加性QTLs,经元分析(meta-analysis)等方法最终获得13个有效加性QTLs,分别位于2号、3号、4号、5号、6号、7号和14号染色体,其中5个QTLs区间缩小到0.5 Mb以内,同时在上述QTLs区间寻找到11个与荚粒性状相关候选基因[25]。
与上述荚粒相关加性QTLs相比,目前有关荚粒性状上位性QTLs及其互作效应分析研究报道甚少,发掘到的上位性遗传位点数量也屈指可数。梁慧珍等[26]对大豆RIL群体6个籽粒性状进行分析,检测到3对影响粒宽和宽厚比的上位性QTLs。杨振等[17]利用大豆RIL群体定位到9对控制二粒荚长的加性×加性上位性QTLs,涉及1号、2号、3号、5号、6号、11号、14号、16号和20号染色体,可解释荚长性状9.02%表型变异,并提出上位性QTLs在大豆荚长、荚宽性状遗传中起着至关重要的作用。由此可见,在已获得加性QTLs基础上,进一步分析控制大豆荚粒性状上位性QTLs才能更充分地揭示荚粒性状分子遗传基础,也才能更准确地实现性状分子遗传改良。
鉴于此,本研究利用前期获得的大豆RIL群体,在2年4种不同环境条件下鉴定其8种鲜荚籽粒相关性状,获得321对性状连锁上位性QTLs,分布于所有染色体;进一步分析发现,其中包含34对“一因多效”上位性QTLs,涉及18条染色体,且有6组“一对多”类型的上位性QTLs,说明大豆荚粒性状除与已报道的单个加性QTLs有关外,还受多个上位性QTLs调控,且这些上位性QTLs形成了复杂的互作网络,进一步揭示这些QTLs互作网络是实现性状分子遗传改良的重要前提。
进一步分析本研究定位到的上位性QTLs可以发现,与前人报道具有一定程度的一致性。本研究在3号染色体ss715585969~ss715585473区间和18号染色体ss715632041~ss715631972区间检测到大豆粒长、粒宽和籽粒面积“一因多效”上位性QTLs,其中3号染色体ss715585969~ss715585473区间与Cui等[27]获得的粒长与粒宽加性QTLs以及Li等[28]获得的荚重加性QTL重叠。同时,本研究在10号染色体和7号染色体检测到控制籽粒面积的上位性QTLs,其中10号染色体的定位区间与梁慧珍等[26]获得的籽粒长宽比QTL距离较近。并且,本研究在13号染色体ss715614097~ss715614125区间与ss715614050 ~ss715614188区间定位到粒长和籽粒面积的“一因多效”上位性QTLs,该QTLs与Hina等[29]定位的籽粒长宽比加性QTL重叠。
同时,本研究还发现,11号染色体ss715610795~ss715610804区间与13号染色体ss715614663~ss715614511区间存在一个粒长、粒宽和籽粒面积“一因多效”上位性QTLs,该QTL的11号染色体区间与杨振等[17]定位的荚长上位性QTLs距离较近,且与Soybase中公布的大豆粒重遗传位点Seedweight49-2[30]一致。同时,6号染色体ss715594872~ss715595080区间与13号染色体ss715615941~ss715615853区间检测到的荚重、粒重和粒长“一因多效”上位性QTLs,与杨振等[17]定位的荚长上位性QTL重叠,且与Soybase中公布的荚宽遗传位点Podwallwidth1-2[31]、Podwallwidth1-3[31],粒重位点Seedweight36-2[32]、Seedweight49-4[30]和Seedweight49-5[30]重叠。这些一致性QTLs一方面验证了本研究结果可靠性,另一方面也说明大豆荚粒加性QTLs除自身主效应外,还可通过与其他位点的互作来调控荚粒形成与发育过程。
除上述与前人报道的一致性加性或上位性位点外,本研究还发现了一些未报道的上位性遗传位点;同时,通过对比本课题组前期已发现的大豆荚粒相关性状加性QTLs可以发现,有很多上位性QTLs并未在加性QTLs中检测到,比如本研究在大豆3号、4号、12号、17号和18号染色体检测到多对上位性QTLs,而在上述染色体中并未检测到有加性QTLs存在。由此可见,并未引起显著加性效应的单个QTL有时可以通过与其他位点的互作来形成显著的上位性效应,进而影响大豆荚粒相关性状。因此,在大豆荚粒性状分子遗传改良中,一方面要考虑加性QTLs,另一方面还应充分考虑上位性QTLs作用。
另外,上位性QTL应用价值还体现在能够揭示已有功能基因分子作用机制方面。艾丽娟等[33]在大豆2号染色体(qHS-2-1)和6号染色体(qHS-6-1)之间检测到一对控制籽粒硬实现象的上位性QTL,通过查阅文献得知,2号染色体存在2个控制种皮渗透性的相关基因,而关于这2个基因的分子作用机制尚不清楚,因而认为可以将这2个功能基因作为出发点,通过寻找与其相关的调控因子或酶类,进而确定6号染色体qHS-6-1区间与其互作的基因,实现解析上述功能基因分子作用机制的目的。因此,上位性QTL的挖掘还可为揭示已有功能基因分子作用机制提供参考。
