基于RSAP标记的10个白杨派无性系指纹图谱构建及遗传分析
2021-09-10程玮哲樊军锋高建社周永学
程玮哲,樊军锋,高建社,周永学
(西北农林科技大学 林学院,陕西 杨凌712100)
杨树为杨柳科(Salicaceae)杨属(PopulusL.)落叶乔木,其分布范围广,生长迅速,具有较强的抗逆性。既是良好的用材和绿化造林树种,又可作为林木遗传研究的优良材料[1-3]。杨树作为雌雄异株植物(极少雌雄同株),某些派间及种间可杂交,从而形成杂种[4]。同时育种研究中培育的优良杂种、无性系,单从形态学上很难区分,这为后续良种鉴定及进一步遗传研究带来困难[5]。随着现代分子生物学的发展,DNA分子标记技术在分析物种遗传关系、鉴别品种、构建遗传图谱等方面具有不可替代的作用[6],这也极大地推进了杨树遗传改良研究及良种生产推广产业的发展。
限制性位点扩增多态性(restriction site amplified polymorphism,RSAP)技术,是杜晓华[7]开发的基于限制性酶切位点的分子标记技术。该技术利用生物基因组上广泛分布的限制性酶切位点,无需限制性酶切及其他复杂步骤,仅一个简单的PCR反应即可实现对DNA限制性位点多态性的检测,具有引物设计简便、操作简单、产物特异性强、结果稳定、中等产率等特点,是RFLP和AFLP技术的简化[8-9]。该分子标记技术目前已运用于紫菜[10]、辣椒[11]、石斛[12]、红花檵木[13]等植物的研究中,但尚未见运用于杨树的研究报道。吕志华等[14]以马铃薯为试验材料,比较了3种分子标记RSAP、SSR和SRAP的分析效力,证明RSAP标记能较好地分析马铃薯的遗传关系,3种标记均有各自的优缺点,在研究植物遗传多样性时可综合3种标记结果,能使分析更全面、客观。王津果等[15]采用RSAP标记的15对引物组合分析龙须菜3个种群的遗传多样性,3个种群的14个个体共扩增出146个多态位点,平均每对引物扩增近10个多态位点,多态性比率为22%,种群间的遗传差异大。中国农业科学院的邹自征[16]利用正交试验对RSAP-PCR反应体系进行了优化,确定了反应最佳体系及退火温度,并分别采用3种分子标记RSAP、SRAP、SSR和田间性状对16份苎麻种质进行了亲缘关系聚类,证明RSAP标记用于苎麻是可行的,并用55个RSAP标记、40个SSR标记和8个SRAP标记成功构建了苎麻连锁遗传图谱。
在运用RSAP分子标记进行植物遗传关系等的研究中,前人大多以琼脂糖电泳和聚丙烯酰胺电泳为检测手段,位点读取存在人为误差,而高灵敏度、高精度的毛细管电泳检测法可自动检测位点信息,使试验结果更加精准[17]。在杨属植物遗传研究中,试验对象大多以种群或一定区域的杨属植物为主[18-20],极少见对优良无性系或近缘种的相关研究。本试验以白杨派10个优良无性系为材料,运用RSAP分子标记结合高精度毛细管电泳检测法,分析白杨派无性系间的遗传多样性及亲缘关系,构建10个无性系的指纹图谱,以期证明RSAP在杨树上的适用性,为杨树种质资源保护与品系鉴定提供一定的理论依据。
1 材料与方法
1.1 试验材料
试验所用杨树材料共10个,均采自西北农林科技大学渭河试验站种质资源库(表1)。包括秦白杨系列中的秦白杨1号、3号、5号无性系及其父母本84K、I-101,新疆杨以及4个白杨派无性系毛白杨30号、07-17-18、07-23-23、07-30-11。2020年5月于试验站苗圃采集无病虫害的健康新生嫩叶,每个杨树材料分别采集4~5片,置于液氮中充分研磨后保存于-80 ℃冰箱。

表1 10个供试白杨派无性系及其遗传背景Table 1 Ten clones of Sect. Leuce and their genetic background
1.2 DNA提取
称取适量(0.1~0.2 g)供试杨树嫩叶,采用改进的CTAB法[21]提取各无性系新生嫩叶基因组DNA。用超微量分光光度计(Nanodrop2000,美国)检测DNA的浓度及纯度,将检测后的DNA稀释至20 ng/μL,并于-20 ℃保存备用,使用时取出保存于4 ℃。
1.3 引物筛选及RSAP-PCR反应
9条RSAP引物的选择参考张明科等[22]的研究,两两组成36对引物组合,由上海生工生物工程股份有限公司合成。首先用8%聚丙烯酰胺凝胶电泳对36对引物组合进行初筛,对扩增清晰、多态性高的引物组合重复电泳3次,最终选出6对扩增状况较好且重复性强的引物组合(表2)。将6对RSAP引物对供试杨树无性系的荧光PCR产物送至上海生工生物工程股份有限公司进行STR分型检测。RSAP反应总体系为20 μL:5 U/μLTaq酶 0.2 μL,10 μmol/L dNTP 1.5 μL,10×Taq buffer(MgCl2) 2.5 μL,10 μmol/L正、反引物各1 μL,20 ng/μL模板DNA 2 μL,ddH2O补足20 μL。反应程序为:94 ℃预变性5 min;94 ℃变性1 min,35 ℃退火1 min,72 ℃延伸1 min,5个循环;94 ℃变性1 min,退火温度(表2)下退火1 min,72 ℃延伸1 min,35个循环;72 ℃延伸10 min;4 ℃保存。
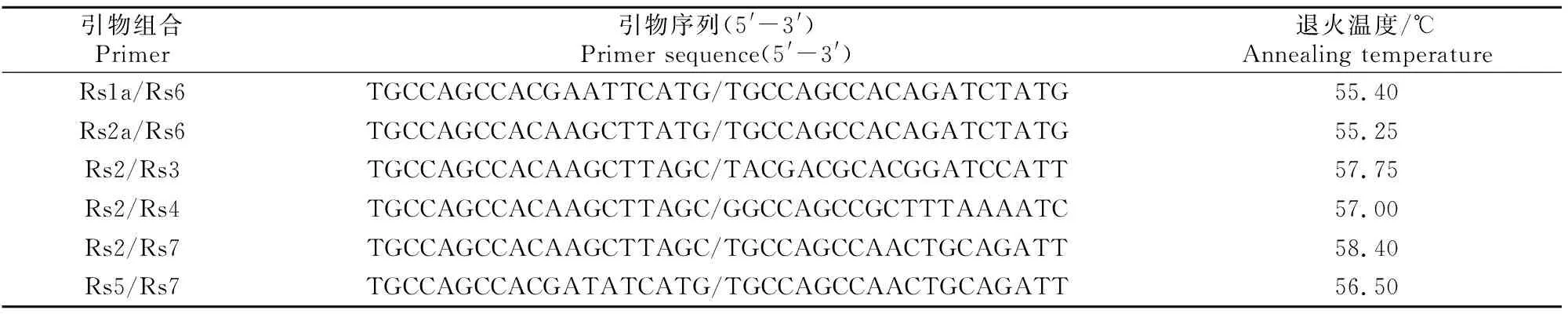
表2 筛选的RSAP引物序列Table 2 Sequences of selected RSAP primers
1.4 10个白杨派无性系扩增结果的多态性分析
利用Excel 2010将试验数据统计为“0/1”格式(有带记为“1”,无带记为“0”),并运用POPGENE 32软件分析电泳结果,统计各引物组合对10个供试白杨派无性系扩增的位点总数、多态性位点数,计算多态性位点百分率、观测等位基因数、有效等位基因数、Shannon信息指数及Nei’s基因多样性指数等遗传多样性相关指标[23-24]。
1.5 10个白杨派无性系指纹图谱的构建
结合扩增结果中多态性和遗传多样性指标,选取多态性高且能区分所有无性系的引物对Rs2/Rs4和Rs2/Rs7,构建10个白杨派无性系的DNA指纹图谱。
1.6 10个白杨派无性系间的遗传相似系数及聚类分析
利用NTSYS 2.1软件Similarity程序组中的SimQual计算各无性系间的遗传相似系数,构建遗传相似系数矩阵;基于UPGMA法进行聚类分析,构建树状聚类图。
2 结果与分析
2.1 10个白杨派无性系的RSAP扩增结果
由表3可知,6对引物组合对10个白杨派无性系的扩增总位点数为143个,平均每个引物组合扩增23.83个,其中Rs2/Rs7引物组合扩增位点数最多,为30个;引物组合Rs1a/Rs6扩增位点数最少,为16个。多态性位点总数为133个,平均每个引物组合扩增22.17个,具有较高的多态性。平均多态性位点百分率为93.01%,引物组合Rs1a/Rs6和Rs5/Rs7的位点多态率为100%,引物组合Rs2/Rs3位点多态率最低。观测等位基因数和有效等位基因数的平均值分别为1.930和1.559;Shannon信息指数为0.212~0.426,平均为0.326;Nei’s基因多样性指数为0.348~0.611,平均为0.488。上述结果表明,10个白杨派无性系间遗传变异性强,遗传多样性丰富。

表3 10个白杨派无性系的RSAP多态引物扩增结果Table 3 Amplification of 10 clones of Sect. Leuce by RSAP polymorphic primers
2.2 10个白杨派无性系的DNA数字化指纹图谱
综合分析6对引物组合扩增位点的多态性,选取2个引物组合的6个多态性位点,即可将10个白杨派无性系有效鉴别。基于2个多态性引物组合构建的10个白杨派无性系指纹图谱见表4。由表4可知,引物组合Rs2/Rs4可对秦白杨1号、秦白杨3号、秦白杨5号、84K、新疆杨、07-23-23进行单独鉴别,结合引物组合Rs2/Rs7可对其余无性系I-101、毛白杨30号、07-17-18、07-30-11进行鉴别。

表4 基于2个多态性引物组合的10个白杨派无性系的指纹图谱Table 4 Fingerprinting of 10 clones of Sect. Leuce based on 2 polymorphic primer combinations
2.3 10个白杨派无性系的遗传相似系数与聚类结果
基于RSAP分子标记的10个白杨派无性系的遗传相似系数矩阵见表5。由表5可以看出,10个白杨派无性系间的遗传相似系数为0.400~0.817,平均遗传相似系数为0.594,变幅为0.417,表明无性系间具有遗传多样性,遗传变异丰富。秦白杨系列的秦白杨1号、3号和5号遗传关系较近,遗传相似系数为0.764~0.817,平均遗传相似系数为0.794;07系列的07-17-18、07-23-23和07-30-11的遗传相似系数为0.695~0.765,平均遗传相似系数为0.732。84K、I-101与秦白杨系列的平均遗传相似系数分别为0.624和0.585,I-101与07系列的平均遗传相似系数为0.699,表明父母本与子代的亲缘关系相近。新疆杨、毛白杨30号无性系与其余无性系遗传背景差异相对较大,因此遗传相似系数也偏低。

表5 10个白杨派无性系间的遗传相似系数Table 5 Genetic similarity coefficient among 10 clones of Sect. Leuce
由图1可见,依据遗传相似系数构建的树状聚类图与实际生物学分类大致相符。在遗传相似系数0.510处,10个白杨派无性系被分为两大类,第一大类为秦白杨系列、84K、I-101及07系列,第二大类为新疆杨和毛白杨30号。第一大类中秦白杨系列与其父本84K聚为一类,07系列与其母本I-101聚为一类,表明秦白杨系列与其父本84K亲缘关系较近。秦白杨1号、秦白杨3号和秦白杨5号聚在一起,07-17-18、07-30-11和07-23-23聚在一起,表明秦白杨系列与07系列分别具有相似的遗传背景,遗传差异较小。

图1 基于遗传相似系数的10个白杨派无性系的树状聚类结果Fig.1 Dendrogram clustering of 10 clones of Sect. Leuce based on genetic similarity coefficient
3 结论与讨论
袁佳秋等[25]利用EST-SSR分子标记技术对12个黑杨无性系进行了遗传结构和亲缘关系分析,从30对引物中筛选出18对多态性引物,多态性比率为70.5%,无性系间的遗传相似系数为0.602~0.904,遗传距离较近。樊蓉等[26]对9个白杨品种进行SRAP分析,筛选的14对多态性引物能较好地反映白杨品种遗传多样性水平,且多态性比率为85.63%,白杨品种间的遗传相似系数为0.400~0.760。藕丹[24]以美洲黑杨青杨派杂种无性系为材料,建立了稳定高效的SCoT-PCR反应体系,并运用SCoT标记分析无性系间的遗传关系,平均每个引物扩增9.07条多态性条带,多态性比率为74.8%。本试验将RSAP分子标记应用于杨树资源,且运用高效、灵敏的毛细管电泳检测扩增产物,平均多态性位点百分率为93.01%,遗传相似系数变幅为0.417,相较于前人研究的其他分子标记[25-27],兼具较高的引物多态性和遗传多样性。
遗传相似系数是评价亲缘关系的常用指标,无性系间的遗传相似系数在一定程度上反应其亲缘关系的远近[28-30]。本试验遗传相似系数表明10个无性系间具有一定的遗传差异,毛白杨30号、新疆杨与秦白杨系列和07系列具有相对较大的遗传背景差异,秦白杨系列无性系中秦白杨1号、3号和5号遗传相似性大,07系列中的07-17-18、07-23-23和07-30-11无性系间遗传相似系数高,遗传差异小。聚类分析结果显示,84K较母本I-101优先与秦白杨系列聚类,而07系列与其母本I-101聚类,这与张海燕等[31]研究结果存在差异,因此推测在基因基础上秦白杨系列与84K亲缘关系较近,在实际生长中则可能受环境、表型性状基因调控等因素影响,秦白杨系列的表型遗传了母本I-101的优良干形特征。秦白杨系列的3个无性系聚为一类,07系列的3个无性系聚为一类,说明其遗传基础相近或相同,这与实际亲缘关系相符。
本试验通过基于限制性位点的RSAP分子标记,分析了10个白杨派无性系的遗传多样性水平,并采用2个引物组合的6个多态位点构建了指纹图谱,为杨树新品种保护与选育奠定了基础。从本试验针对杨树开展的RSAP分子标记的运用结果来看,RSAP扩增位点数量多且扩增位点多态性比率高,能用较少的引物对无性系(品种)进行鉴别,具有一定可行性,可应用于杨树分子生物学研究。
