莱州湾鱼卵、仔稚鱼群落结构及多样性的季节变化
2021-09-09杨艳艳朱明明徐炳庆李凡高彦洁孙春晓魏潇王斌
杨艳艳,朱明明,徐炳庆,李凡*,高彦洁,孙春晓,魏潇,王斌
( 1.山东省海洋资源与环境研究院 山东省海洋生态修复重点实验室,山东 烟台 264006;2.鲁东大学 滨海生态高等研究院,山东 烟台 264025)
1 引言
鱼卵和仔稚鱼在海洋生态系统能量传递中起重要作用[1],是鱼类发育过程中的重要阶段。同时,鱼卵和仔稚鱼发育过程是鱼类生命周期中两个非常短暂,但形态学、生理学和生态学等特性变化明显的过渡时期,也是整个鱼类发育周期中最为脆弱的阶段[2-3]。这一发育阶段成活率的高低、剩存量的多寡将决定鱼类补充群体资源量的丰歉,是鱼类资源可持续利用研究中不可缺少的内容[4]。从20世纪50代开始,我国开展了多次鱼卵、仔稚鱼的监测调查工作,1964-1981年在开展渤海中国明对虾(Fenneropenaeus chinensis)产卵场调查时,就收集过鱼卵、仔稚鱼的资料[5]。
莱州湾位于渤海南部,平均水深为10 m[6-7],是多种经济种类产卵和育肥的场所[8],在渔业经济中占重要地位[9-10]。从20世纪50年代至21世纪初,我国生态学者、环境学者对该海域渔业资源的状况进行过广泛的研究[11-13]。近60 a来,受过度捕捞和环境变化的影响,该海域渔业资源结构发生重大变迁[14],生态环境质量呈现明显下降的趋势[7,15-16]。有报道显示[17],2000年以来,莱州湾鱼卵、仔鱼数量和浮游生物的种数及多样性等均呈下降趋势。在这种背景下,摸清莱州湾海域鱼卵、仔稚鱼的种类组成、数量分布以及群落结构和多样性的季节变化对了解渔业资源的补充状况、保护渤海生物资源有很重要的现实意义。近年来,国内有关莱州湾海域鱼卵、仔稚鱼的研究很多,但是大多数研究集中在春季和夏季,例如王爱勇等[18]和高彦洁等[19]对莱州湾春季的鱼卵、仔稚鱼进行了研究;还有一些研究是集中在莱州湾的某一个特定海域,例如卞晓东等[20]对黄河口海域的鱼卵和仔稚鱼进行了研究;高东奎等[21]对莱州湾人工鱼礁区的鱼卵和仔稚鱼进行了研究等。有关莱州湾海域不同季节的鱼卵、仔稚鱼群落结构和生物多样性变化的研究较少。
本研究基于2014-2015年莱州湾海域的采样数据,分析了该海域鱼卵、仔稚鱼的种类组成、数量分布、优势种等,探讨了该海域鱼卵和仔稚鱼的群落结构和生物多样性的季节变动规律,以期为莱州湾海域渔业资源修复和保护提供理论依据。
2 材料方法
2.1 调查时间与站位分布
于2014-2015年在莱州湾海域进行4个航次的鱼类浮游生物拖网调查。调查时间分别为2014年5月(春季)、8月(夏季)、10月(秋季)和 2015年 2月(冬季)。在莱州湾海域的调查范围(37°10′~37°45′N,119°00′~120°10′E)内共布设 20 个站位(图1a),由于2015年2月(冬季)受天气影响,实际完成调查12 个站位(图1b)。
图1 莱州湾调查站位Fig.1 Sample stations in the Laizhou Bay
2.2 样品采集
样品采集及分析方法严格按照《海洋调查规范》[22]进行,租用渔业生产船只,使用口径为80 cm,长为280 cm的大型浮游生物网在海域表层进行水平拖拽采集。每站拖拽10 min,拖速为2 n mile/h,采集到的样品用中性甲醛溶液固定保存,带回实验室,根据鱼卵、仔稚鱼的形态特征(包括鱼卵和仔稚鱼的外部形态以及不同发育时期的个体形态和组织、器官、系统发育特征等)利用解剖镜进行种类鉴定[23-28],并计数。
2.3 数据分析方法
2.3.1 优势度
利用相对重要性指数(Index of Relative Importance, IRI)对鱼卵和仔稚鱼的优势种进行计算[20],公式为
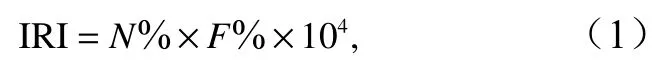
式中,N%为某一种类个体数量占采获总量的百分比;F%为某一种类出现的站位数占总调查站位数的百分比,IRI不小于1 000的种类为优势种。
2.3.2 生物多样性
丰富度指数(D)计算公式为

香农-威纳多样性指数(H′)计算公式为

均匀度指数(J′)计算公式为

式中,S为种类数;N为总个体数;Pi为第i种个体数占总个体数的比例。
2.3.3 季节间种类更替率
季节间种类更替率(E)计算公式为

式中,A为季节间种类增加与减少数之和;B为季节间相同的种类数[20]。
2.3.4 群落结构分析
利用R软件的vegan、cluster等程序包计算2014-2015年不同季节间种类Bray-Curtis相似性系数矩阵,并利用非加权组平均法进行聚类。
3 结果与分析
3.1 种类组成
调查期间采集鱼卵135 275粒,仔稚鱼2 456尾,共22种(表1)。其中1种仔稚鱼未鉴定出种,1种鰕虎鱼科仔稚鱼鉴定到科,其余20种隶属于9目16科20属。22种鱼卵、仔稚鱼中,鱼卵13种,仔稚鱼15种。鲈形目种类最多,为7种,鲱形目4种,鲽形目3种,鲻形目2种,颌针鱼目、银汉鱼目、鼠鱚目、刺鱼目、鲉形目和胡瓜鱼目各1种。

表1 莱州湾鱼卵、仔稚鱼种类组成Table 1 Species composition of ichthyoplankton in the Laizhou Bay
3.2 优势种及其季节变化
5月(春季)和8月(夏季)调查中,鱼卵的优势种分别为鳀(Engraulis japonicus)和赤鼻棱鳀(Thryssa kammalensis),仔稚鱼的优势种分别为鮻(Liza haematocheilus)、布氏银汉鱼(Allanetta bleekeri),仔稚鱼冬季的优势种为长绵鳚(Zoarces elongatus)。10月(秋季)和2月(冬季)未采获到鱼卵所以无法计算优势种,10月(秋季)调查所采集的仔稚鱼样品中无优势种存在,仅大银鱼(Protosalanx hyalocranius)、中颌棱鳀(Thryssa mystax)和青鳞小沙丁(Sardinella zunasi)构成重要种(表2)。
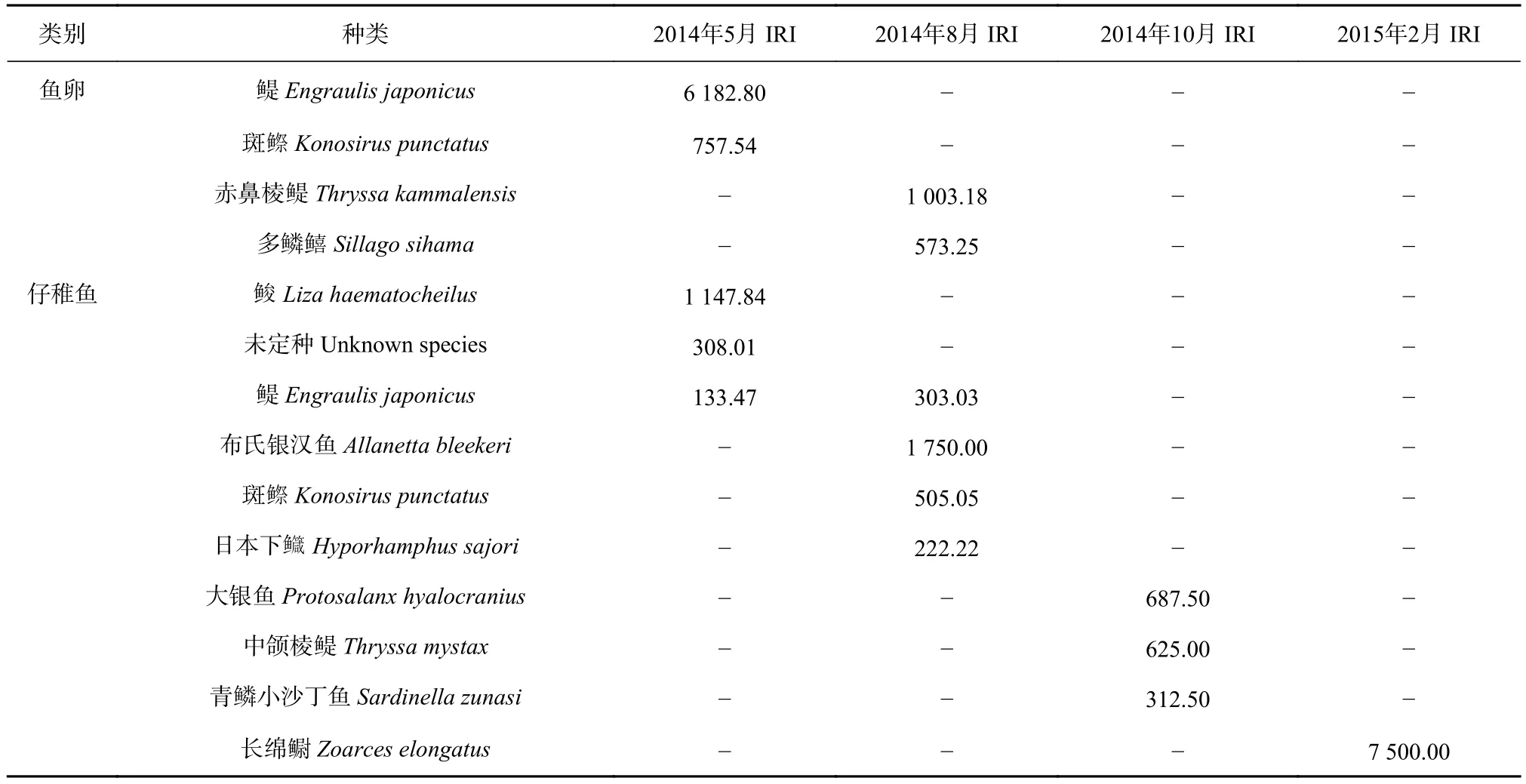
表2 莱州湾海域鱼卵、仔稚鱼优势种和重要种组成Table 2 Dominant and important species composition of ichthyoplankton in the Laizhou Bay
3.3 丰度时间和空间变化
调查中共采集鱼卵135 275粒,仔稚鱼2 456尾。鱼卵、仔稚鱼平均密度分别为1 878.82粒/站和34.11 尾/站。
5月(春季)采获鱼卵135 118粒,平均密度为6 755.90粒/站,波动范围为8~112 340粒/网。最高值出现在3号站,最低值出现在1号和7号站位(图2a)。其中,鳀卵数量最多,占本航次调查鱼卵数量的88.32%。8月(夏季)采获鱼卵157粒,平均密度7.85粒/站,波动范围为0~95粒/网,最高值出现在18号站位,为95粒/站,赤鼻棱鳀鱼卵数量最多,占本航次调查鱼卵数量的66.88%(图2b)。秋季和冬季没有采获鱼卵(图2c,图2d)。
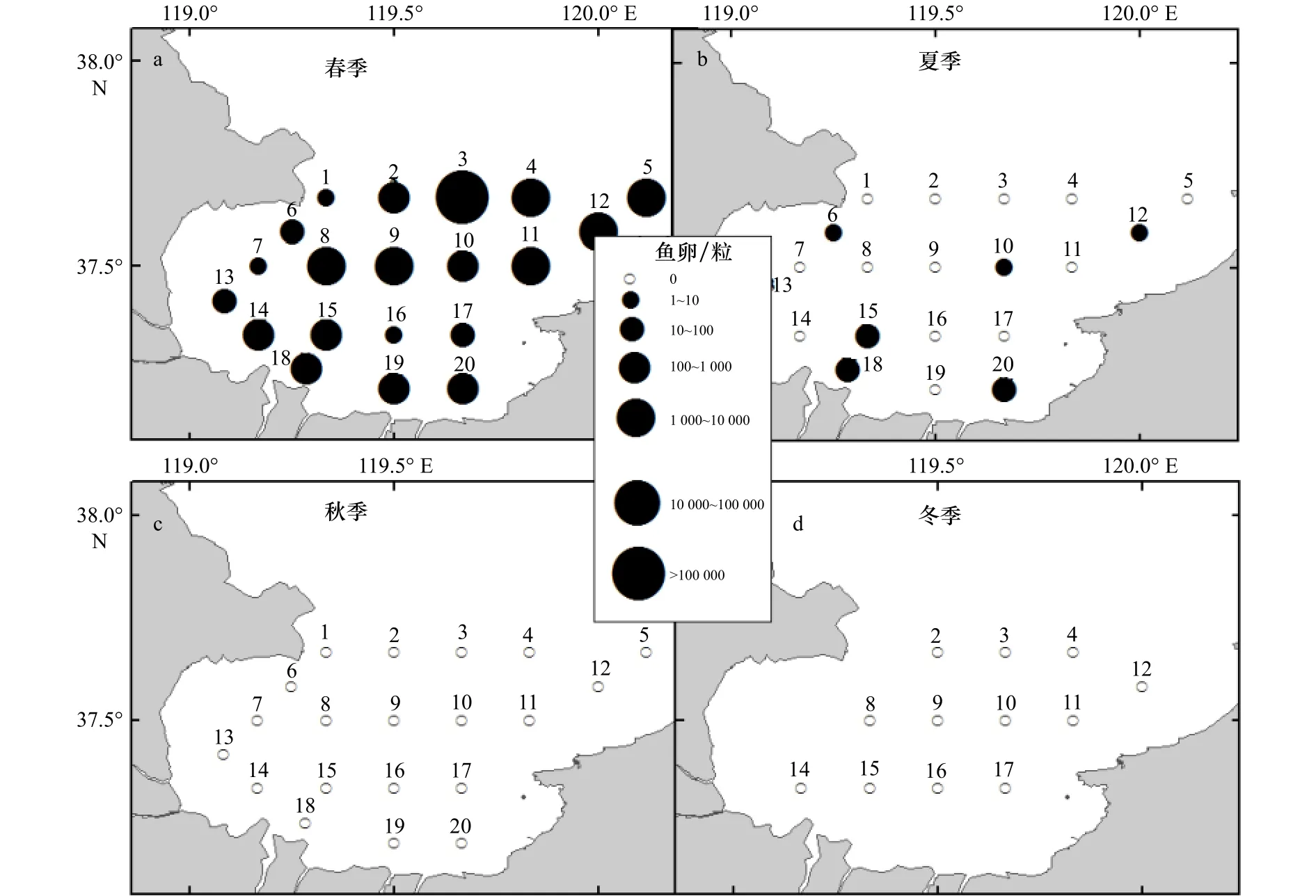
图2 莱州湾海域鱼卵空间分布Fig.2 Distribution of fish eggs in the Laizhou Bay
5月(春季)采获仔稚鱼1 948尾,平均密度为97.4尾/站,波动范围为0~760尾/网,最高值出现在13号站位,为760尾/站(图3a)。其中,鮻仔稚鱼的数量最多,占本航次调查仔稚鱼数量的57.39%。8月(夏季)采获仔稚鱼198尾,平均密度为9.9尾/站,波动范围为0~35尾/网,最高值出现在16号站位(图3b)。其中,布氏银汉鱼数量最多,占本航次调查仔稚鱼数量的50%。10月(秋季)采获仔稚鱼24尾,平均密度为1.2尾/站,波动范围为0~13尾/网,最高值出现在1号站位(图3c)。其中大银鱼仔稚鱼数量最多,占本航次调查仔稚鱼数量的84.62%。2月(冬季)采获仔稚鱼286尾,平均密度为23.83尾/站,最高值出现在4号站位(图3d)。全部为长绵鳚的仔稚鱼。从全年变化范围来看,鱼卵、仔稚鱼分布不均匀,资源密度总体呈现春夏高,秋冬低的趋势。
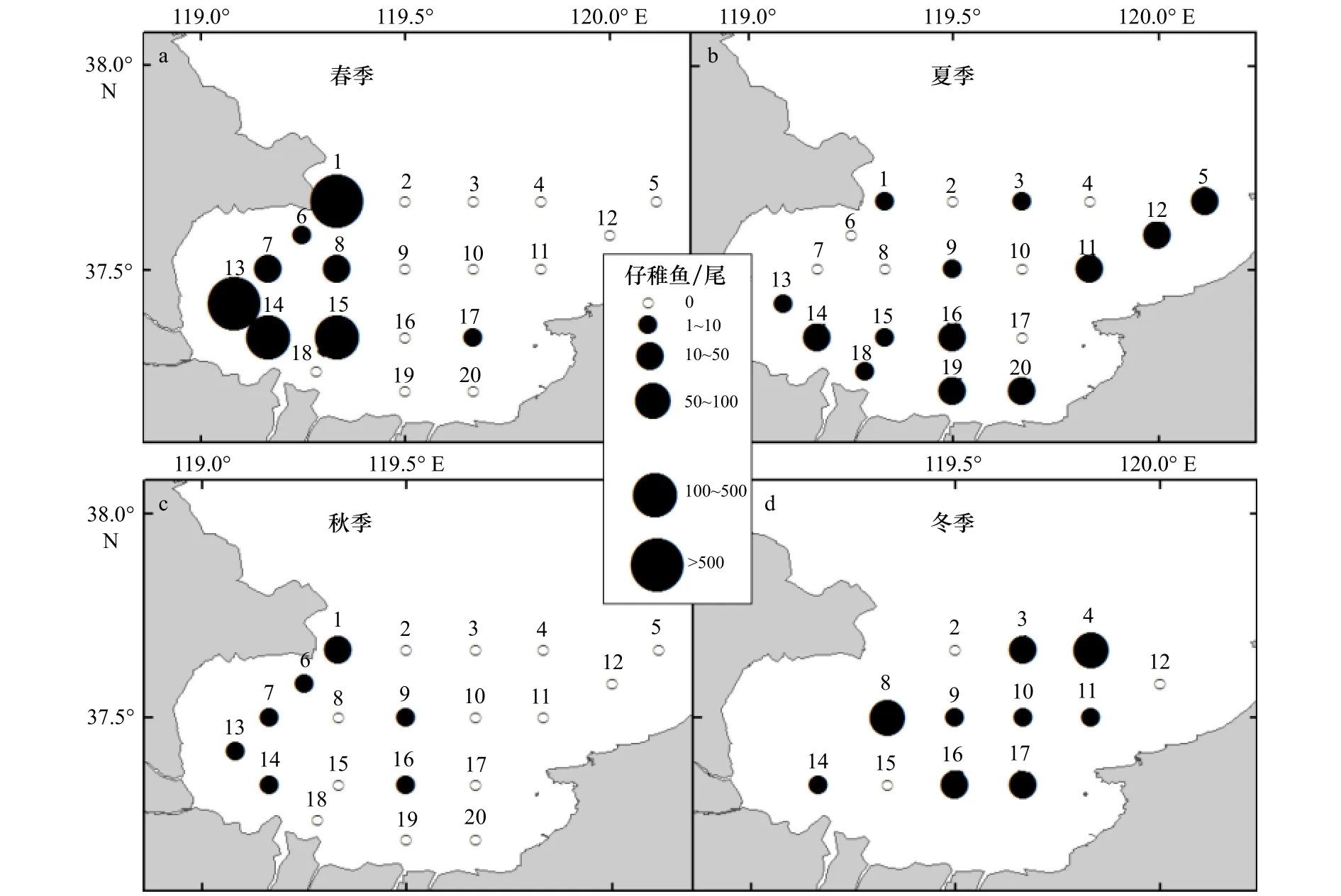
图3 莱州湾海域仔稚鱼空间分布Fig.3 Distribution of larvae in the Laizhou Bay
3.4 群落多样性的季节变化
2015年2 月(冬季),因只采获到长绵鳚1个种类,所以丰富度指数(D)和多样性指数(H′)均为0,均匀度指数(J′)则无法计算。由图4可知,5月(春季)的丰富度指数(D)和多样性指数(H′)均为全年最低,均匀度指数(J′)则为除 2月(冬季)外最低。8月(夏季)的丰富度指数(D)和多样性指数(H′)均为最高,丰富度指数(D)和多样性指数(H′)全年呈先升高后下降的趋势;10月(秋季)的均匀度指数(J′)为除2月外全年最高,均匀度指数(J′)全年除2月外呈现逐渐上升的趋势。
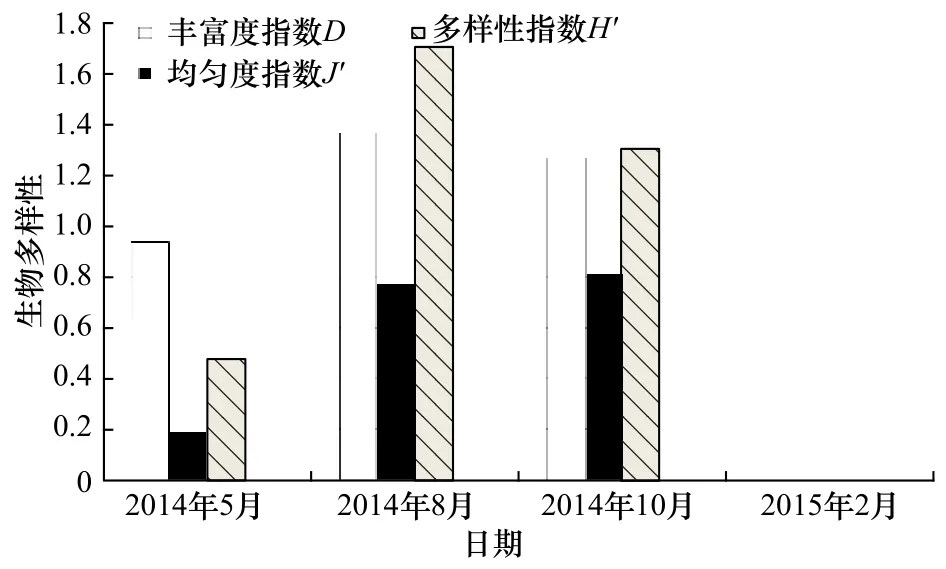
图4 莱州湾海域鱼卵、仔稚鱼物种多样性Fig.4 Species diversity of ichthyoplankton in the Laizhou Bay
3.5 种类更替和群落结构的季节间相似性聚类
由图5可知,2014-2015年调查期间莱州湾海域各季节间鱼卵和仔稚鱼的种类更替率均在80%以上,且随季节变化呈现上升的趋势,从春季到夏季的种类更替率最小,为89.47%,其次为夏季到秋季,为92.31%,秋季到冬季以及冬季到春季的种类更替率均为 100%(图5)。
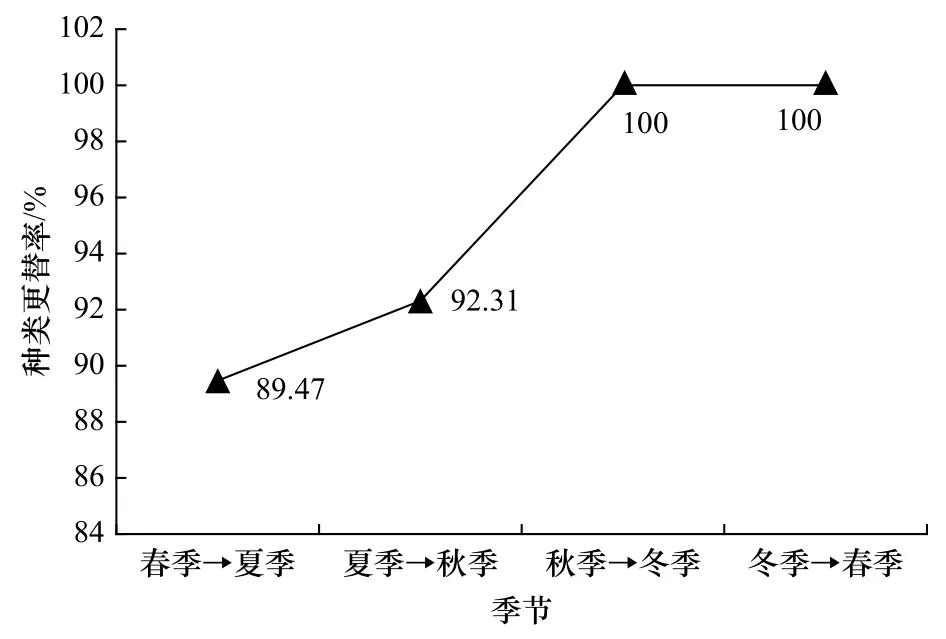
图5 莱州湾海域鱼卵、仔稚鱼种类更替率Fig.5 The species turnover rate of ichthyoplankton in the Laizhou Bay
利用Bray-Curtis相似性系数公式计算不同季节间种类组成的相似性系数,然后运用R语言聚类树对其进行聚类分析,根据融合水平值定义划分水平,选择具有最大跳跃的分组水平,将4个季节的鱼卵、仔稚鱼划分为3个组,进一步聚类分析结果显示,3个分组分别为春组、夏秋组和冬组(图6,图7),其中,春组和夏秋组、春组和冬组主要分歧种均为鳀,鳀对其季节间平均相异性贡献率分别为87.28%和87.22%;夏秋组和冬组间主要分歧种为长绵鳚和赤鼻棱鳀,这两个种类对两个组季节间平均相异性贡献率为76.63%。
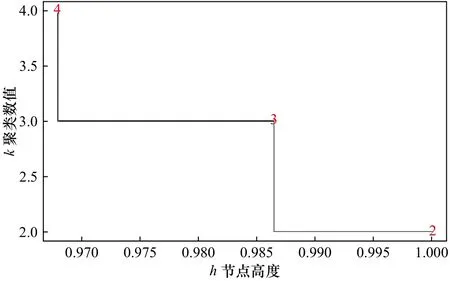
图6 聚类树的融合水平值Fig.6 Fusion level values of cluster tree
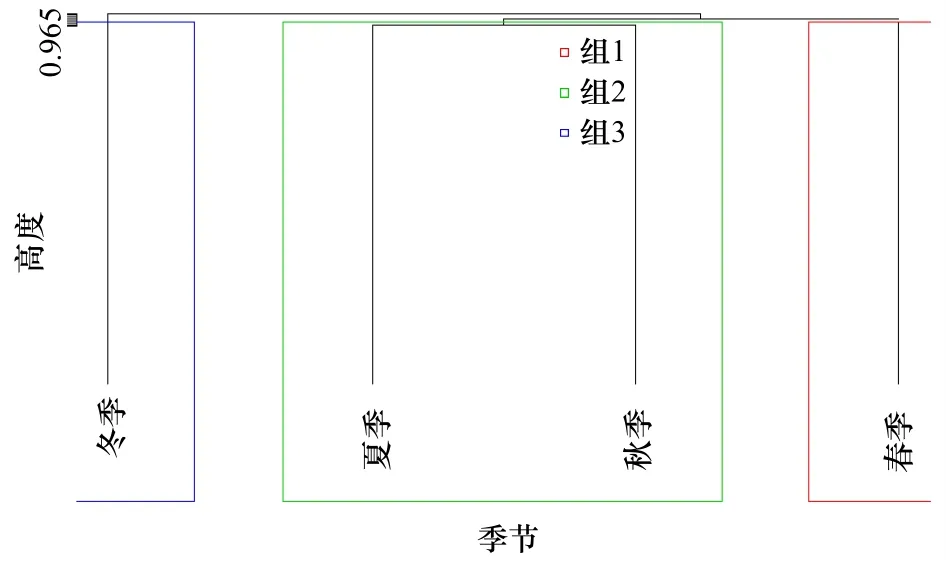
图7 莱州湾海域鱼卵、仔稚鱼群落结构季节变化聚类分析Fig.7 Cluster analysis of seasonal changes in the community structure of ichthyoplankton in the Laizhou Bay
4 讨论
4.1 种类组成和数量分布的季节更替
本研究调查结果显示,莱州湾海域鱼卵、仔稚鱼的种类和数量随季节而更替,从鱼卵、仔稚鱼数量的季节变化来看,春季(5月)为鱼类的产卵盛期,鱼卵和仔稚鱼的数量最多,占全年总量的90%以上,夏季数量大幅减少,秋冬季节进入产卵末期以后数量也随之降至最低点,两季鱼卵和仔稚鱼数量仅占全年数量的0.2%左右[7]。
从种类数的季节变化来看,5月(春季)捕获鱼卵、仔稚鱼 11种,8月(夏季)9种,10月(秋季)5种,2月(冬季)1种,种类数呈现随季节转变而下降的趋势,这种季节变化主要是洄游性种类的季节性分布引起的:春季洄游性的种类(主要为暖水种和暖温种) 进入莱州湾产卵、索饵,这一时期种数较冬季增加;秋末,随着水温的降低,这些种类离开莱州湾至越冬场,仅定居种留下[29]。
从种类组成的季节变化来看,5月(春季)为一年中的产卵盛期,此时鱼卵集中在湾口海域,仔稚鱼则集中在西部海域,鱼卵、仔稚鱼的优势种为陆续进入湾口等近岸水域产卵的鳀和从深水区回到河口产卵的鮻,5月调查中捕获鮻仔稚鱼1 118尾,鱼卵89粒,且鮻多为初孵的仔鱼(初孵仔鱼占鮻仔稚鱼总数的98.2%),而鮻的孵化时间为42 h,所以分析可能是调查时间错过了鮻产卵盛期;春季到夏季的种类更替率为89.47%,鱼卵、仔稚鱼的优势种从春季暖温性中上层洄游鱼类的鳀和暖温性河口定居鱼类鮻变为夏季暖水性中上层洄游鱼类赤鼻棱鳀和布氏银汉鱼,且鱼卵优势种优势度下降,仔稚鱼优势种优势度上升。因为夏季近岸水温比远岸高,所以此时鱼卵、仔稚鱼的分布向湾底近岸集中。夏季和秋季之间的种类更替率为92.31%,秋季和冬季之间的种类更替率为100%,夏季到秋季再到冬季主要种类变化依次为暖水性中上层洄游鱼类的赤鼻棱鳀和布氏银汉鱼变为秋季暖温性中上层溯河洄游性鱼类大银鱼,再到冬季冷温性底层鱼类长绵鳚,形成了种类与环境和季节相对应的动态格局[20],同时也说明在不同的季节,产卵群体的组成和结构也发生了显著的变化,而季节间种类的高更替率则说明鱼类群落结构的时序相对不稳定[20]。分析这种不稳定产生的原因,一方面与莱州湾物种洄游性季节分布有关,另一方面由于近年来莱州湾海域的环境变化和人类活动等因素,使种类更替率明显加快[30-31]。另外,秋冬季节没有捕获到鱼卵,这是因为本研究中调查网具为浮游生物网,调查方法为表层拖网,而莱州湾海域秋冬季节仅有极少数产浮性卵的底栖鱼类,细纹狮子鱼(Liparis tanakae)、大泷六线鱼(Hexagrammos otakii)等秋冬季节繁殖的鱼类均产黏着沉性卵,不适于浮游生物网采集。
4.2 物种多样性
由于春、夏季是莱州湾海域鱼类繁殖的重要季节,有关秋、冬季鱼卵、仔稚鱼多样性指数(H′)可参考的数据很少,所以本研究中仅对莱州湾海域春、夏季不同调查时期的多样性指数(H′)进行对比。
马克平和刘玉明[32]提出,多样性指数(H′)的合理范围为1.5~3.5,本研究中莱州湾海域鱼卵、仔稚鱼群落的多样性指数(H′)只有夏季处于此范围之内,春季、秋季和冬季均低于该范围,将春季多样性指数(H′)与莱州湾以及邻近海域不同时期调查相比较(表3),发现其均处于较低水平。同时,鱼卵、仔稚鱼群落D、J′也都较低,均低于夏季和秋季,说明春季鱼卵、仔稚鱼的结构相对不稳定,种间分布不均匀[20]。鱼类群落在长期严重的外界干扰下,物种多样性会丧失[33],而人类的捕捞是对鱼类群落最主要的外界干扰因素,在人类捕捞活动的长期干扰下,鱼类群落多样性不断下降,从而导致作为其补充群体的鱼卵、仔稚鱼的群落多样性也不断下降[34],另外,根据多样性指数的生态含义分析,在种类数较多,但是种间分布极不均匀的情况下,多样性偏低。某些优势种在个别海区的聚集,使这些站位个体数量在不同种间分配差异加大,也会使多样性偏低[35-36]。本研究符合这种情况:春季航次鱼卵种类虽多,但是除鳀以外的其他种类数量较少,鳀占该航次鱼卵总数的88.3%,成为该航次的绝对优势种;而鳀仔稚鱼虽然没有成为春季航次仔稚鱼的绝对优势种,但是其数量占该航次仔稚鱼总数量的近1/3,并且分布极不均匀,只在1号站位出现,这就导致了春季鱼卵、仔稚鱼的H′较低。
夏季H′在与2007年夏季黄河口以南和以北海域的比较中均处于较高水平(表3),但是因为参考文献的局限性,只有1 a的数据作为参考,并且其调查区域的H′可能会受到黄河调水调沙的影响而导致偏小,所以不能确定2014年夏季H′与莱州湾及邻近海域不同时期调查相比是否处于较高水平,有待于今后的调查和研究中进一步验证。另外,夏季的D和H′均大于秋季,但是J′略小于秋季。因为秋季种类和数量分布比较均匀,鱼卵和仔稚鱼均没有出现优势种;夏季种类和数量虽超过秋季,但是出现了赤鼻棱鳀和布氏银汉鱼两个相对高优势度的物种,所以导致J′略小于秋季。鱼类的繁殖生长和捕捞活动也会造成多样性的季节变化[37],8月的休渔,使莱州湾海域的外界干扰减小,而10月秋季所处时间段为开捕后,人类捕捞干扰增强,加上水温降低,导致洄游性鱼类离开近岸水域,鱼类种类数和数量均减少,所以D和H′均出现下降趋势。
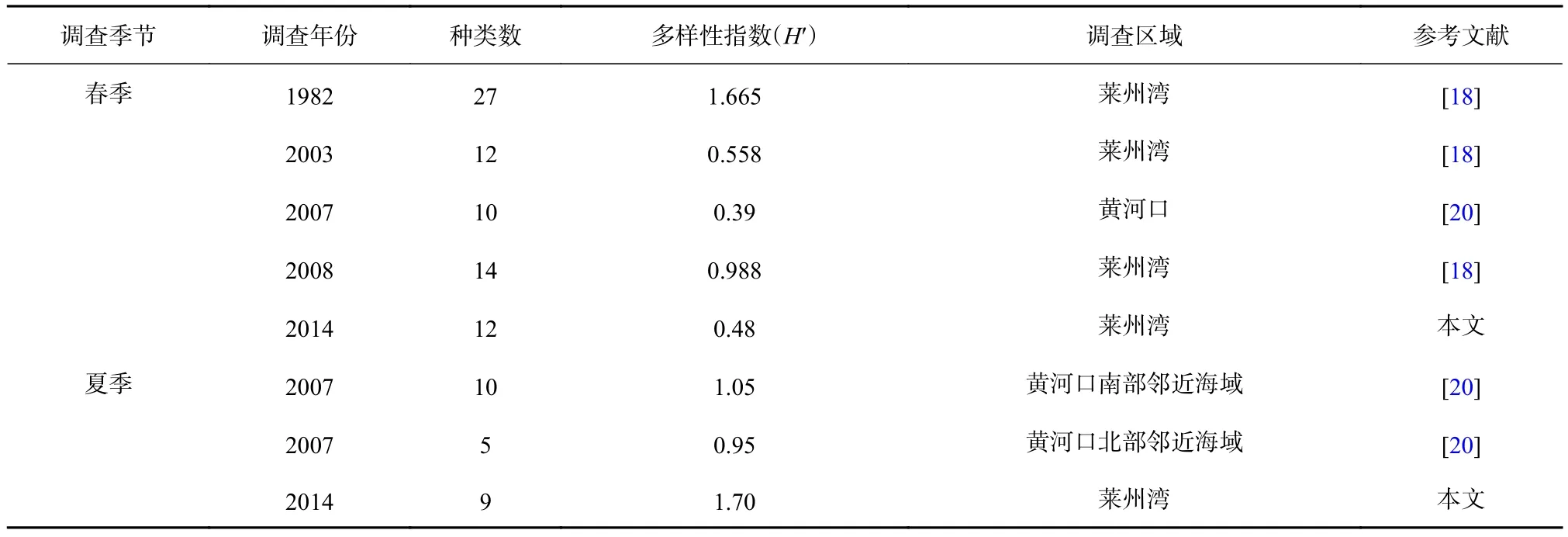
表3 莱州湾及邻近海域不同时期调查春、夏季鱼卵、仔稚鱼的群落多样性( H′ )Table 3 Community diversity of ichthyoplankton in the Laizhou Bay and its adjacent waters in different periods (H′)
4.3 群落结构
莱州湾水域是渤海的传统渔场,也是黄、渤海众多渔业生物的产卵场、育幼场和索饵场[14],对黄、渤海渔业资源的补充具有重要意义[10-11]。近年来,受人类活动以及环境变化等的共同影响,使得莱州湾的渔业资源结构发生了重大变迁。
从鱼卵、仔稚鱼种类组成来看,本研究与黄、渤海海域大面调查的结果存在一定的差异,程济生[38]于1998年5-11月在渤海海域进行调查,采获鱼卵、仔稚鱼共41种。万瑞景和姜言伟[39-40]于1982年4月至1983年4月和1992年8月至1993年6月在渤海海域的调查中,共采获鱼卵61种、仔稚鱼49种;1985-1988年在北黄海和南黄海的调查中采获68种鱼卵、仔稚鱼。从与黄、渤海大面调查的历史资料纵向对比来看,莱州湾海域鱼卵、仔稚鱼的种类数仅为或者不足黄、渤海海域20世纪80、90年代的1/2。说明莱州湾海域鱼卵、仔稚鱼的种类在大幅减少,主要与近年来的过度捕捞和环境退化有关。而于旭光等[41]2008年在辽东湾近岸海域的调查以及郭书新等[4]2014年和2015年在青岛崂山青山湾人工鱼礁区调查的种类数均低于本研究;肖欢欢等[42]2015年在海州湾及邻近海域3个航次的调查采获鱼卵25种,仔稚鱼13种,高于本研究采获的种类数。从与山东海域其他海湾的横向对比来看,种类数目高低不同,种类组成也存在一定的差异,说明鱼卵、仔稚鱼的种类组成因调查海域范围、调查时间以及调查海域的生态环境等因素的不同而存在差异,这种差异也反映了不同海域不同时期鱼类群落结构的差异[4]。另外,将本研究中优势种种类与1998年渤海调查相比,油魣(Sphyraena pinguis)、棘头梅童鱼(Collichthys lucidus)、叫姑鱼(Johnius belangerii)、小黄鱼(Larimichthys polyactis)、蓝点马鲛(Scomberomorus niphonius)、玉筋鱼(Ammodytes personatus)、短鳍红娘鱼(Lepidotrigla micropterus)、许氏平鲉(Sebastods schlegelii)、大泷六线鱼等均缺失[7]。而鳀、鮻、赤鼻棱鳀、斑鰶(Konosirus punctatus)等种类比例有所上升。并且2014-2015年间,高优势度物种的季节演替较为明显,不存在高优势度物种的季节间重叠,与胶州湾和青山湾的研究结果一致,证明种类更替呈现加快的趋势[4,43]。
从空间生态类型来看,莱州湾海域目前以小型中上层低经济价值鱼类的鱼卵、仔稚鱼为主,中上层鱼类和经济价值较低鱼类的鱼卵、仔稚鱼均超过全年总量的95%,而底层鱼类和经济价值较高鱼类的鱼卵、仔稚鱼分别仅占全年数量的3.7%和2.1%。进一步验证了目前莱州湾渔业资源结构正在向中上层、小型低值鱼类类群演化的趋势[11,44-45]。这种演变趋势主要与环境条件的变化和人类的捕捞活动有关。持续的全球变暖使渤海的水温不断升高,造成鱼类早期补充群体的生理周期提前,导致其与饵料的错配,使鱼类早期补充能力下降,进而导致渔业资源的衰退[46]。另外,有研究表明,渤海内捕捞强度自1950年以来增长了近40倍[47],在环境变化和捕捞活动的双重干扰下,渤海鱼类资源急剧下降[16,48-49],小黄鱼、带鱼(Trichiurus japonicas)等种类,曾是山东近海最重要的底层经济鱼类,自1962年秋捕对虾以来,大量损害了它们的幼鱼,不论在育肥场还是越冬场的密集区,均遭到大量的捕捞,极大破坏了底层经济鱼类资源。进入21世纪以来,底层鱼类比例仍处在较低水平上。而小型中上层鱼类的数量开始增加,20世纪90年代以后,鳀、斑鰶等小型中上层鱼类逐渐成为黄、渤海主要的捕捞品种[9,16,50-51],从而导致作为鱼类早期补充群体的鱼卵、仔稚鱼的结构也发生相应的改变[31]。
从摄食习性来看,本研究中亲体为浮游动物食性和底栖动物食性的鱼卵、仔稚鱼大约占采集样品种数的70%,说明莱州湾目前以浮游动物食性和底栖动物食性的鱼类为主[52]。Jin等[51]发现莱州湾的优势种从高营养级和广食性鱼类,向寿命短、浮游生物食性的低营养级中上层鱼类演替。同时,渤海生态系统捕捞渔获物的平均营养级由1982年的2.92下降到2014-2015年的2.68,总的变化趋势为低营养层次种类的生物量百分比增加,高营养层次种类的生物量百分比减少[53-55]。过度捕捞和环境退化导致渤海生态系统稳定性不断降低,莱州湾生态系统压力巨大,而海湾区域多是水生生物重要的栖息地和重要的产卵场、育幼场、索饵场所,对渔业资源的补充非常重要。因此,加强对海湾生态系统的恢复和保护是当前非常紧迫的任务。
